
冬小麦光合特性对CO2浓度与土壤含水量的响应机制*
2017-07-18 11:31:02周晓旭张玉翠沈逸群姜寒冰沈彦俊
中国生态农业学报(中英文) 2017年7期
周晓旭,张玉翠,吴 林,沈逸群,姜寒冰,沈彦俊
(1.中国科学院农业水资源重点实验室/河北省节水农业重点实验室/中国科学院遗传与发育生物学研究所农业资源研究中心石家庄 050022;2.中国科学院大学 北京 100049;3.青海师范大学生命与地理科学学院 西宁 810000;4.河北师范大学附属中学 石家庄 050011)
冬小麦光合特性对CO2浓度与土壤含水量的响应机制*
周晓旭1,2,张玉翠1**,吴 林1,3,沈逸群4,姜寒冰1,2,沈彦俊1
(1.中国科学院农业水资源重点实验室/河北省节水农业重点实验室/中国科学院遗传与发育生物学研究所农业资源研究中心石家庄 050022;2.中国科学院大学 北京 100049;3.青海师范大学生命与地理科学学院 西宁 810000;4.河北师范大学附属中学 石家庄 050011)
光合作用是衡量植物对环境响应的重要指标,通过光响应曲线拟合量化光合特征,可从生理机制方面揭示出植物在不同生长环境下自身的调节与适应机制。本文利用Li-6400便携式光合仪测定了冬小麦在4种不同处理条件下灌浆期旗叶的光响应曲线,采用直角双曲线模型(RHM)、非直角双曲线模型(NRHM)、直角双曲线修正模型(RHMM)、指数模型(EM)和指数改进模型(MEM)对光响应数据进行拟合,分析了不同CO2浓度和土壤含水量对冬小麦光合特征的影响。结果表明,直角双曲线修正模型对各处理下冬小麦光响应曲线和光响应曲线参数的拟合值都与实测值较为接近,拟合效果最好;随着CO2浓度升高,各水分处理下冬小麦表观量子效率(α)、光饱和点(LSP)和最大净光合速率(Pnmax)增大,光补偿点(LCP)和暗呼吸(Rd)降低,即CO2浓度升高能有效增加冬小麦的光能转化率和光能利用范围,提高冬小麦的光合能力;随着土壤含水量的降低,冬小麦光补偿点(LCP)和暗呼吸速率(Rd)升高,但表观量子效率(α)、光饱和点(LSP)和最大净光合速率(Pnmax)降低,即冬小麦虽然能通过提高初始光合效率抵消一部分干旱胁迫的影响,但干旱胁迫仍会降低冬小麦光合能力;此外,CO2浓度的增加能抵消部分冬小麦因干旱胁迫引起的光合能力降低。
冬小麦;CO2浓度升高;干旱胁迫;光响应曲线模型;光合能力
由于人类对化石燃料的过度使用与大量森林的乱砍滥伐,大气CO2浓度以每年1.5~2.0 μmol·mol-1的速率快速增长[1],2016年大气CO2浓度已升高到407.7 μmol·mol-1[2],预计21世纪末大气CO2浓度将达到700 μmol·mol-1[3]。随着CO2等温室气体的排放,全球气候变暖,降雨类型发生改变,地区性干旱发生频繁[4]。冬小麦(Triticum aestivumL.)属于C3植物,对CO2浓度变化较为敏感;同时华北地区冬小麦生长雨热不同期,降雨量只能满足生育期需水的1/3[5],因此研究华北平原不同CO2浓度和土壤含水量条件下冬小麦的光响应曲线及其响应特征具有重要意义。
光合作用是植物对环境响应的直接表现,确定植物的光响应曲线是了解植物光合特性的重要方法,通过对其拟合分析,可以估算如表观量子效率(α)、光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Pnmax)、暗呼吸速率(Rd)等众多重要的光合参数。大量研究表明,CO2浓度升高有效提高了植物的Pnmax[6]、LCP[7],而随着土壤含水量的降低,植物的Pnmax、α、LCP均呈降低趋势[8-9]。目前研究光响应曲线的模型很多,应用较为广泛的为直角双曲线模型和非直角双曲线模型[10-14],但这两种模型不能直接计算出饱和光强。叶子飘等[15-17]在直角双曲线模型的基础上提出了修正模型,此模型克服了上述模型的缺点,且能拟合出光抑制的现象。张利阳等[18]通过对毛竹(Phyllostachys pubescensMazel)光响应曲线的拟合得出,指数模型计算出的光饱和点和最大净光合速率更为合理。陈兰英等[19-20]利用指数改进模型对紫茉莉(Mirabilis jalapL.)、高粱(Sorghum vulgarePers.)、大麦等(Hordeum vulgareL.)进行分析拟合,结果表明指数改进模型可准确计算出C3与C4植物的饱和光强和最大净光合速率。
目前,国内外有关CO2浓度升高和干旱胁迫条件对作物光合作用影响的研究虽多,但对利用不同光响应模型拟合分析CO2与土壤水分交互作用对作物光合特性影响的研究却很少。本研究利用以上5种模型对不同CO2浓度和土壤含水量条件下的冬小麦光响应曲线进行拟合,旨在选出冬小麦在不同生长环境下的适宜模型,并进一步分析冬小麦对不同CO2浓度与土壤含水量条件的光合作用响应规律,为未来气候条件下冬小麦高产低耗提供理论依据。
1 材料与方法
1.1 试验地概况与试验方法
试验区选在中国科学院栾城农业生态系统试验站(37°53′N,114°41′E)进行,地处太行山前平原中部,属于半干旱半湿润季风气候,冬小麦生长季降水稀少,平水年仅为100 mm[21]。试验选在适宜水分(约75%田间持水量)和干旱胁迫(约55%田间持水量)两个水分池进行,水分池面积为5 m×10 m,为防止水分的侧向交换,两水分池之间埋设1.5 m隔离墙。
供试材料为冬小麦,品种为‘科农199’,于2015年10月17日播种,翌年6月5日收获,60 cm等行距播种,生育期内施肥磷酸二铵562.5 kg·hm-2。
根据CO2浓度与土壤含水量不同,试验设置4个处理:1)CO2浓度400 μmol·mol-1,适宜水分(W1);2)CO2浓度750 μmol·mol-1,适宜水分(W2);3)CO2浓度400 μmol·mol-1,干旱胁迫(D1);4)CO2浓度750 μmol·mol-1,干旱胁迫(D2)。于2016年5月10日至5月24日期间(小麦灌浆期)的晴朗天气上午9:00—11:30,采用Li-6400便携式光合仪(Li-Cor Inc.,USA)进行数据测量。采用LI-6400-02B人工光源测量叶室,测量时大气相对湿度约为60%,温度设定为25℃,光合有效辐射(PAR)梯度(μmol·mol-1·s-2)为1 500、1 400、1 200、1 000、800、600、400、200、150、100、50、20、0,CO2浓度分别设定为400 μmol·mol-1和750 μmol·mol-1。为保证各处理组试验条件一致,测量时每组处理选取3株长势一致的小麦对旗叶进行测定。
1.2 光响应曲线模型
所采用的光响应曲线模型如下:直角双曲线模型(RHM)[22]:
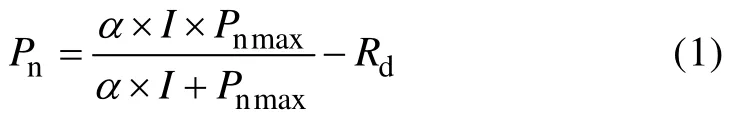
式中:Pn为净光合速率(μmol·m-2·s-1),α为表观量子效率(μmol·mol-1),I为光量子通量密度(μmol·m-2·s-1),Pnmax为最大净光合速率(μmol·m-2·s-1),Rd为暗呼吸速率(μmol·m-2·s-1)。该模型为一个无极值的函数,因此不能直接估算植物的饱和光强。
非直角双曲线模型(NRHM)[22]:

式中:θ为非直角双曲线凸度,其他参数意义同式(1)。与直角双曲线模型一样,该模型为无极值函数,饱和光强不能直接进行估算。
直角双曲线修正模型(RHMM)[23]:

式中:β、γ为模型修正系数,其他参数意义同式(1)。此模型为有极值的函数,可由式(3)直接求取植物的光饱和点、光补偿点、最大净光合速率等。
指数模型(EM)[24]:

式中:各参数意义同式(1)。此模型函数同样无极值,估算饱和光强时,假设Pn等于0.90Pnmax时所对应的光照强度为饱和光强[25]。
指数改进模型(MEM)[20]:
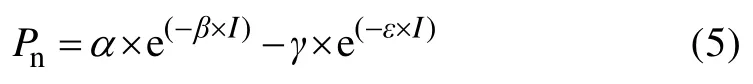
式中:α、β、γ、ε均为模型系数,其他参数意义同式(1)。
为了更好地检验光响应曲线拟合值与实测值的接近程度,定义相对误差(RE):

式中:y和yi分别为光响应曲线参数的实测值和拟合值,RE越小,则拟合效果越好。
2 结果与分析
2.1 各模型对光响应曲线的拟合比较
由图1a可以看出直角双曲线模型对于W2处理下冬小麦的光响应曲线拟合结果较好;但对于W1、D1、D2处理下光响应曲线的拟合值与实测值差异极大,在PAR处于600~1 000 μmol·m-2·s-1范围内时,对D1和D2处理下的拟合值偏小于实测值,且当W1、D1、D2处理下冬小麦由于受到的光能“过剩”,净光合速率达到饱和后逐渐降低出现光抑制现象时[26],拟合值却一直处于上升趋势,不能很好地拟合出冬小麦的光抑制现象。由图1b和图1d可以看出,非直角双曲线模型和指数模型对各处理下冬小麦光响应曲线拟合结果与直角双曲线模型拟合结果相似,都对W2处理有较好的拟合结果,但不能拟合出W1、D1和D2处理下的光抑制现象,这是由于这3种模型方程都为不存在极点的渐近线,由此求出的净光合速率只能随着PAR的增加而增大。由图1c可以看出,直角双曲线修正模型对各处理下光响应曲线都有较好的拟合结果,拟合值与实测值一致,且能很好地拟合出W1、D1、D2处理下的光抑制现象。由图1e可以看出,指数改进模型不仅对W2拟合结果较好,且能很好地拟合出各处理下的光抑制现象,但对D2处理下光响应曲线的拟合值与实测值存在较大偏差。
2.2 各模型对冬小麦光响应曲线拟合参数的比较
光响应曲线参数包括表观量子效率(α)、光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Pnmax)和暗呼吸速率(Rd),各模型拟合值与实测值对比结果如表1。通过对各参数拟合值与实测值的综合对比可知,各处理下都以直角双曲线修正模型对光响应曲线参数的拟合结果最好,而其他模型除个别参数的拟合值与实测值较为接近外,多数参数拟合值与实测值差别较大。α是显示植物光合作用光能转化效率的重要指标之一,大田条件下长势良好的植物α值在0.04~0.07[27],随着土壤含水量的降低,其值逐渐降低[28]。由于CO2的肥效作用在一定程度上增强了作物的光合能力,因此随着CO2浓度的升高,α值逐渐增大。通过对比各模型对α的拟合结果可知,直角双曲线模型拟合值明显偏高,其他模型拟合值更符合实际情况。由于直角双曲线模型与非直角双曲线模型通过求解线性方程的方法计算LSP,其拟合值远小于实测值,这与很多研究结果一致[15-17,29];而指数模型则利用0.9Pnmax[25]或0.99Pnmax[29]所对应的光合强度为饱和光强的方法来计算LSP,这种方法具有很强的人为性,并不具有生物学意义,且不能求解出D1处理下的LSP。直角双曲线模型与非直角双曲线模型拟合出的Pnmax偏大于实测值,这与许多研究结果相一致[15-16,30-31]。指数改进模型虽能很好地拟合出光抑制现象,但其拟合参数多与实测值有较大偏差,与直角双曲线修正模型相比拟合效果较差。
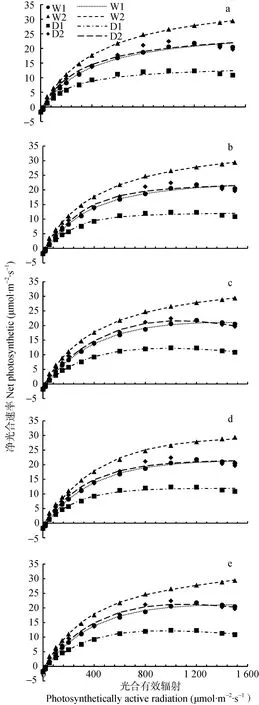
图1 直角双曲线模型(a)、非直角双曲线模型(b)、直角双曲线修正模型(c)、指数模型(d)和指数改进模型(e)对冬小麦光响应曲线的拟合Fig.1 Light response curves of winter wheat fitted by rectangular hyperbolic model (a),non-rectangula hyperbolic model(b),rectangular hyperbolic modified model (c),exponental model (d) and modifier exponental model (e)
为进一步判断各模型对冬小麦光响应曲线拟合值与实测值的偏差程度,将各模型对冬小麦光响应曲线参数拟合值与实测值进行相对误差分析,相对误差越小,说明拟合值与实测值越接近,拟合精度越高。由图 2可知,直角双曲线修正模型对各处理下冬小麦光响应曲线各参数拟合值与实测值的相对误差都较小,且整体拟合相对误差最小,拟合值更为接近实测值,其中以对Pnmax和LSP的拟合结果最好。其余4个模型只对个别参数拟合相对误差较小,其余参数拟合较差,无法兼顾各参数的整体拟合效果。
2.3 不同CO2浓度和土壤含水量对冬小麦光合作用的影响
由于直角双曲线修正模型在光响应曲线及其参数模拟方面表现较好,故选取该模型对冬小麦灌浆期旗叶光响应曲线进行拟合,分析不同CO2浓度和土壤含水量对冬小麦灌浆期旗叶光合作用的影响。由图3可以看出,在PAR低于400 μmol·m-2·s-1时,各处理下冬小麦Pn对PAR响应敏感,随PAR增加而迅速增加,当PAR高于400 μmol·m-2·s-1后,各处理下冬小麦的Pn增加速率逐渐减小。而当PAR大于1 000 μmol·m-2·s-1时,由于CO2浓度和土壤含水量的差异,各处理下冬小麦的Pn对PAR的响应出现差异:D1和D2处理下冬小麦Pn在PAR约为1 100 μmol·m-2·s-1时达到光饱和,W1处理下冬小麦Pn在PAR约为1 300 μmol·m-2·s-1时达到光饱和,之后3个处理下冬小麦Pn随PAR的增加而降低,出现光抑制现象;而W2处理下冬小麦Pn则随着PAR的增加一直处于增加趋势,出现非光饱和现象。这说明CO2浓度升高能显著提高冬小麦光能利用率,促进冬小麦的光合作用。在相同光照条件下,W2处理下冬小麦Pn最高,D1处理下冬小麦Pn最低,即CO2浓度和土壤含水量升高均能使冬小麦净光合速率提高,增强光合作用。而在PAR处于200~1 300 μmol·m-2·s-1时,D2处理下冬小麦净光合速率明显大于W1处理,即在一定范围的光照强度下,CO2对冬小麦的影响作用大于土壤含水量的影响,且CO2浓度的增加能够抵消一部分由于水分亏缺造成冬小麦净光合速率降低,提高其光合能力。

表1 不同处理下不同模型的冬小麦光响应曲线参数拟合值与实测值对比Table 1 Model-fitted and measured values of parameters of light response curves of winter wheat under different treatments

图2 不同模型模拟的不同处理下冬小麦光响应曲线参数拟合值与实测值的相对误差Fig.2 Relative errors of model-fitted and measured values of parameters of light response curves of winter wheat under different treatments
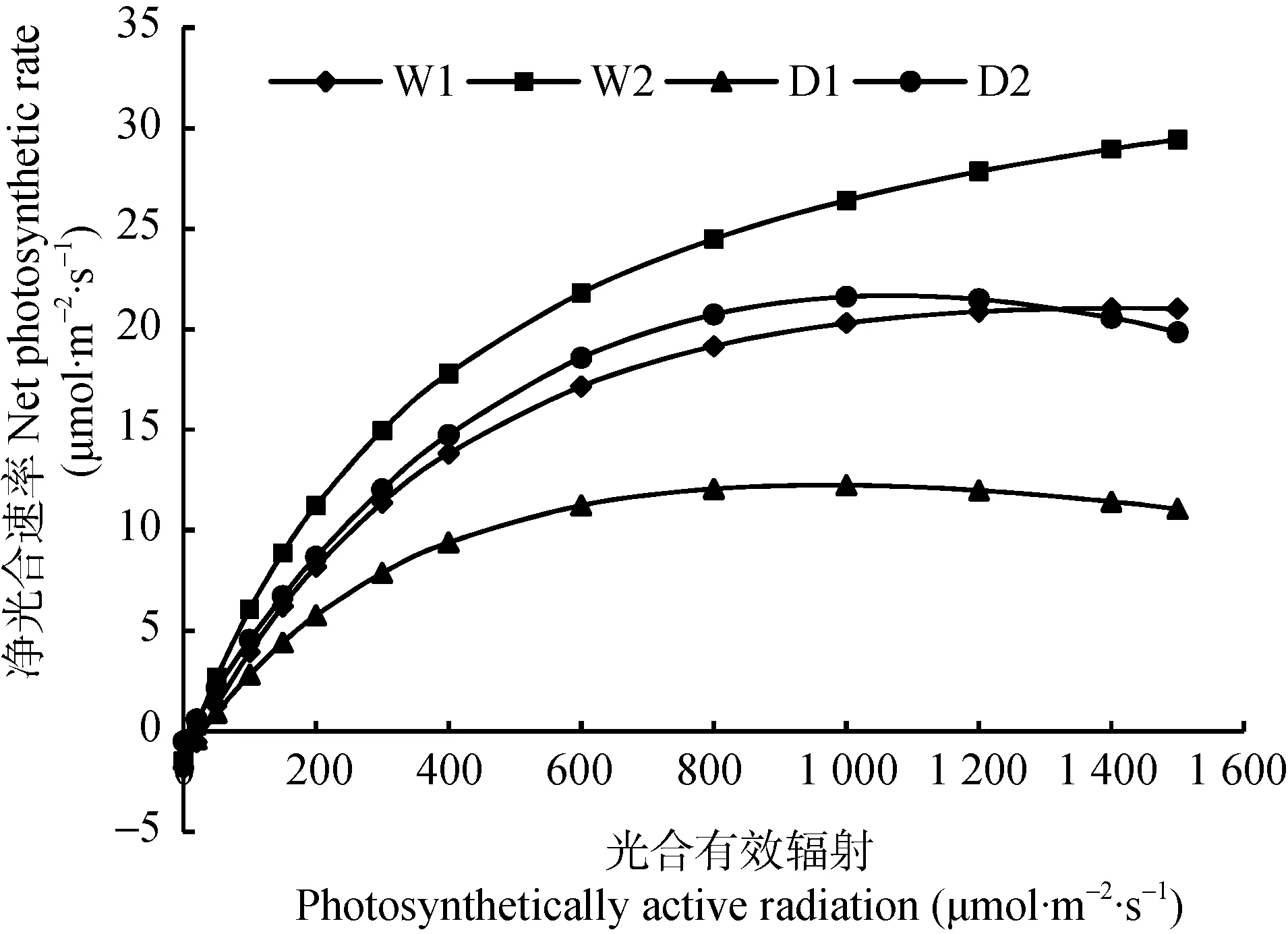
图3 不同处理下冬小麦灌浆期旗叶光响应曲线Fig.3 Light response curves of flag leaves of winter wheat under different treatments of CO2concentration and soil water conditions
为进一步分析CO2浓度和土壤含水量对冬小麦光合特性的影响,对各处理下冬小麦光响应参数进行分析对比。α是衡量植物光合作用光能转化效率的重要指标之一[32],α值越高,则代表植物转化利用光能的能力越强。结合表1分析可知,W2处理下冬小麦α值明显高于其他处理,W1处理和D1处理α差异不大,D1处理α值最小,说明CO2浓度和土壤含水量升高能明显提高冬小麦光能转化效率,使其保持较高的光合能力。在一条件下,Pnmax代表植物叶片的最大光合能力[33]。冬小麦Pnmax在W2处理下明显高于W1处理,W1处理明显高于D1处理,这说明CO2浓度和土壤含水量升高都能显著提高冬小麦叶片最大光合能力,且W2处理比W1处理增加32.77%,而D2处理下比D1处理下增加了48.28%,说明CO2浓度升高冬小麦叶片光合能力的促进作用在干旱胁迫条件下大于适宜水分条件下。此外,D1处理下冬小麦Pnmax较W1处理下降低43%,D2处理下冬小麦Pnmax较W2处理下降低24%,进一步说明CO2浓度升高对冬小麦因干旱胁迫引起的光合能力下降有一定的补偿作用。
LCP和LSP分别代表植物利用弱光和强光的能力,LCP越低说明植物利用弱光能力越强,LSP越高,则说明植物利用强光能力越强[27]。Rd则反映植物自身代谢速率的高低[34]。由于CO2浓度的增加,W2处理下冬小麦LCP低于W1处理,而LSP远高于W1处理,且Rd比W1处理降低31%,这说明CO2浓度升高明显增加了冬小麦的光强利用范围,提高了冬小麦对光的生态适应能力,且降低了冬小麦的代谢速率,提高冬小麦的光合作用,具有一定的肥效作用,这与Ainsworth等[35]和Long等[36]的研究结果相一致。由于受到干旱胁迫影响,D1处理下冬小麦LCP与Rd均低于W1处理,说明干旱胁迫条件下冬小麦通过提高利用弱光的能力和降低自身的生理代谢来提高自身的初始光合效率[37],但与W1处理下相比,D1处理下冬小麦LSP与Pnmax仍然较低,这说明在一定干旱程度下冬小麦虽然能通过提高初始光合效率来抵消部分水分胁迫的影响,但干旱胁迫依然降低了冬小麦的光合能力。
3 结论
通过对比5个模型(直角双曲线模型、非直角双曲线模型、直角双曲线修正模型、指数模型与指数改进模型)在不同CO2浓度与土壤含水量条件下对冬小麦灌浆期旗叶光响应曲线及参数的拟合结果发现,直角双曲线修正模型适用于各处理下冬小麦光响应曲线及其参数的拟合;同时,该模型可以很好地拟合出低CO2浓度与低土壤含水量条件下冬小麦因不能利用较强光照而出现的光抑制现象。
对直角双曲线修正模型拟合出的冬小麦光响应曲线及参数进行分析对比可知,CO2浓度升高能有效地提高冬小麦表观量子效率(α)、光饱和点(LSP)和最大净光合速率(Pnmax),降低冬小麦光补偿点(LCP)和暗呼吸速率(Rd),从而提高冬小麦光能转化率和最大净光合速率,增加光能利用范围和初始光合效率,即CO2浓度升高能有效增强冬小麦的光合能力;当土壤含水量较低时,冬小麦通过提高自身初始光合效率抵消了一部分干旱胁迫的影响,但α、LSP和Pnmax与适宜水分相比仍明显降低,即干旱胁迫使冬小麦光合能力明显降低,光合作用减弱;此外,CO2浓度的升高对干旱胁迫引起的光合作用降低有一定的补偿作用,且CO2浓度升高对冬小麦光合能力的增强作用在干旱胁迫条件下大于适宜水分条件下。
致谢 感谢赵彦茜(河北地质大学)、安塞(河北科技大学)和任晓东(青海师范大学)对本试验的帮助和支持。
References
[1]IPCC.Climate Change 2007:The Physical Science Basis[R].Cambridge:Cambridge University Press,2007
[2]CO2Now.Earth’s CO2home page[EB/OL].[2016-05].http://www.co2.earth/monthly-co2
[3]Waton R T,Rodhe H,Oescheger H,et al.Greenhouse gases and aerosols[M]//Houghton J T,Jenkins G J,Ephraums J J.Climate Change:The IPCC Scientific Assessment.Cambridge:Cambridge University Press,1990:1–40
[4]Sgherri C L M,Quartacci M F,Menconi M,et al.Interactions between drought and elevated CO2on alfalfa plants[J].Journal of Plant Physiology,1998,152(1):118–124
[5]毛飞,霍志国,李世奎,等.中国北方冬小麦播种期底墒干旱模型[J].自然灾害学报,2003,12(2):85–91 Mao F,Huo Z G,Li S K,et al.A drought model of soil moisture in seedtime of winter wheat in north of China[J].Journal of Natural Disasters,2003,12(2):85–91
[6]于显枫,张绪成.高CO2浓度和遮荫对小麦叶片光能利用特性及产量构成因子的影响[J].中国生态农业学报,2012,20(7):895–900 Yu X F,Zhang X C.Effects of elevated atmospheric CO2concentration and shading on leaf light utilization and yield of wheat[J].Chinese Journal of Eco-Agriculture,2012,20(7):895–900
[7]李清明,刘彬彬,邹志荣.CO2浓度倍增对干旱胁迫下黄瓜幼苗光合特性的影响[J].中国农业科学,2011,44(5):963–971 Li Q M,Liu B B,Zou Z R.Effects of doubled CO2concentration on photosynthetic characteristics of cucumber seedlings under drought stresses[J].Scientia Agricultura Sinica,2011,44(5):963–971
[8]陆燕元,马焕成,李昊民,等.土壤干旱对转基因甘薯光合曲线的响应[J].生态学报,2015,35(7):2155–2160 Lu Y Y,Ma H C,Li H M,et al.Light response characteristics of photosynthetic of transgenic sweet potato under drought stress[J].Acta Ecologica Sinica,2015,35(7):2155–2160
[9]于艳梅,徐俊增,彭世彰,等.不同水分条件下水稻光合作用的光响应模型的比较[J].节水灌溉,2012,(10):30–33 Yu Y M,Xu J Z,Peng S Z,et al.Evaluation of photosynthesis light response model for rice leaf under different water conditions[J].Water Saving Irrigation,2012,(10):30–33
[10]Farquhar G D,von Caemmerer S,Berry J A.A biochemical model of photosynthetic CO2assimilation in leaves of C3species[J].Planta,1980,149(1):78–90
[11]Thornley J H M.Instantaneous canopy photosynthesis:Analytical expressions for sun and shade leaves based on exponential light decay down the canopy and an acclimated non-rectangular hyperbola for leaf photosynthesis[J].Annals of Botany,2002,89(4):451–458
[12]陆佩玲,于强,罗毅,等.冬小麦光合作用的光响应曲线的拟合[J].中国农业气象,2001,22(2):12–14 Lu P L,Yu Q,Luo Y,et al.Fitting light response curves of photosynthesis of winter wheat[J].Chinese Journal of Agrometeorology,2001,22(2):12–14
[13]刘宇峰,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,21(8):76–79 Liu Y F,Xiao L T,Tong J H,et al.Primary application on the non-rectangular hyperbola model for photosynthetic light-response curve[J].Chinese Agricultural Science Bulletin,2005,21(8):76–79
[14]谭念童,林琪,李玲燕,等.限量补灌对旱地冬小麦灌浆期旗叶光响应及产量的影响[J].华北农学报,2010,25(4):145–151 Tan N T,Lin Q,Li L Y,et al.Effects of limited irrigation on light-response of flag leaves and grain yield in dry-land winter wheat at filling stage[J].Acta Agriculturae Boreali-Sinica,2010,25(4):145–151
[15]叶子飘,于强.一个光合作用光响应新模型与传统模型的比较[J].沈阳农业大学学报,2007,38(6):771–775 Ye Z P,Yu Q.Comparison of a new model of light response of photosynthesis with traditional model[J].Journal of Shenyang Agricultural University,2007,38(6):771–775
[16]叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356–1361 Ye Z P,Yu Q.Comparison of new and several classical models of photosynthesis in response to irradiance[J].Journal of Plant Ecology,2008,32(6):1356–1361
[17]叶子飘,李进省.光合作用对光响应的直角双曲线修正模型和非直角双曲线模型的对比研究[J].井冈山大学学报:自然科学版,2010,31(3):38–44 Ye Z P,Li J S.Comparative investigation light response of photosynthesis on non-rectangular hyperbola model and modified model of rectangular hyperbola[J].Journal of Jinggangshan University:Natural Science,2010,31(3):38–44
[18]张利阳,温国胜,王圣杰,等.毛竹光响应模型适用性分析[J].浙江农林大学学报,2011,28(2):187–193 Zhang L Y,Wen G S,Wang S J,et al.Four light-response models to estimate photosynthesis ofPhyllostachys pubescens[J].Journal of Zhejiang A &F University,2011,28(2):187–193
[19]陈兰英,黎云祥,钱一凡,等.改进指数模型对紫茉莉光合—光响应及CO2响应适用性研究[J].广西植物,2013,33(6):839–845 Chen L Y,Li Y X,Qian Y F,et al.Applications studies of the modified exponential model on photosynthesis-light response and CO2response curves ofMirabilis jalapa[J].Guihaia,2003,33(6):839–845
[20]陈卫英,陈真勇,罗辅燕,等.光响应曲线的指数改进模型与常用模型比较[J].植物生态学报,2012,36(12):1277–1285 Chen W Y,Chen Z Y,Luo F Y,et al.Comparison between modified exponential model and common models of light-response curve[J].Chinese Journal of Plant Ecology,2012,36(12):1277–1285
[21]Sun H Y,Shen Y J,Yu Q,et al.Effect of precipitation change on water balance and WUE of the winter wheat-summer maize rotation in the North China Plain[J].Agricultural Water Management,2010,97(8):1139–1145
[22]Thornley J H M.Mathematical Models in Plant Physiology[M].London:Academic Press,1976:86–110
[23]Ye Z P.A new model for relationship between irradiance and the rate of photosynthesis inOryza sativa[J].Photosynthetica,2007,45(4):637–640
[24]Bassman J H,Zwier J C.Gas exchange characteristics ofPopulus trichocarpa,Populus deltoidesandPopulus trichocarpa×P.deltoidesclones[J].Tree Physiology,1991,8(2):145–159
[25]王满莲,冯玉龙,李新.紫茎泽兰和飞机草的形态和光合特性对磷营养的响应[J].应用生态学报,2006,17(4):602–606 Wang M L,Feng Y L,Li X.Effects of soil phosphorus level on morphological and photosynthetic characteristics ofAgeratina adenophoraandChromolaena odorata[J].Chinese Journal of Applied Ecology,2006,17(4):602–606
[26]Powles S B.Photoinhibition of photosynthesis induced by visible light[J].Annual Review of Plant Physiology,1984,35:15–44
[27]Long S P,Humphries S,Falkowski P G.Photoinhibition of photosynthesis in nature[J].Annual Review of Plant Physiology and Plant Molecular Biology,1994,45:633–662
[28]郭春芳,孙云,张木清.土壤水分胁迫对茶树光合作用—光响应特性的影响[J].中国生态农业学报,2008,16(6):1413–1418 Guo C F,Sun Y,Zhang M Q.Effect of soil water stress on photosynthetic light response curve of tea plant (Camellia sinensis)[J].Chinese Journal of Eco-Agriculture,2008,16(6):1413–1418
[29]段爱国,张建国.光合作用光响应曲线模型选择及低光强属性界定[J].林业科学研究,2009,22(6):765–771 Duan A G,Zhang J G.Selection of models of photosynthesis in response to irradiance and definition of attribute of weak light[J].Forest Research,2009,22(6):765–771
[30]黄红英,窦新永,孙蓓育,等.两种不同生态型麻疯树夏季光合特性的比较[J].生态学报,2009,29(6):2861–2867 Huang H Y,Dou X Y,Sun B Y,et al.Comparison of photosynthetic characteristics in two ecotypes ofJatropha curcasin summer[J].Acta Ecologica Sinica,2009,29(6):2861–2867
[31]Kyei-Boahen S,Lada R,Astatkie T,et al.Photosynthetic response of carrots to varying irradiances[J].Photosynthetica,2003,41(2):301–305
[32]Yu Q,Zhang Y Q,Liu Y F,et al.Simulation of the stomatal conductance of winter wheat in response to light,temperature and CO2changes[J].Annals of Botany,2004,93(4):435–441
[33]伍维模,李志军,罗青红,等.土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响[J].林业科学,2007,43(5):30–35 Wu W M,Li Z J,Luo Q H,et al.Effects of soil water stress on light response curves of photosynthesis ofPopulus euphraticaandPopulus pruinosa[J].Scientia Silvae Sinicae,2007,43(5):30–35
[34]张劲松,孟平,高峻.板蓝根光合及水分生理生态特性[J].东北林业大学学报,2004,32(3):26–28 Zhang J S,Meng P,Gao J.Characteristics of photosynthesis and water physio-ecology ofIsatis indigotica[J].Journal of Northeast Forestry University,2004,32(3):26–28
[35]Ainsworth E A,Long S P.What have we learned from 15 years of free-air CO2enrichment (FACE)? A meta-analytic review of the responses of photosynthesis,canopy properties and plant production to rising CO2[J].New Phytologist,2005,165(2):351–372
[36]Long S P,Ainsworth E A,Rogers A,et al.Rising atmospheric carbon dioxide:Plants FACE the future[J].Annual Review of Plant Biology,2004,55:591–628
[37]邹琦.作物抗旱生理生态研究[M].济南:山东科学技术出版社,1994:155–283 Zou Q.Research on Eco-physiological Responses of Plant to Drought Conditiond[M].Jinan:Shandong Science and Technology Press,1994:155–283
Response mechanism of photosynthetic characteristics of winter wheat to CO2concentration and soil water content*
ZHOU Xiaoxu1,2,ZHANG Yucui1**,WU Lin1,3,SHEN Yiqun4,JIANG Hanbing1,2,SHEN Yanjun1
(1.Key Laboratory of Agricultural Water Resources,Chinese Academy of Sciences / Key Laboratory of Water Saving Agriculture,Hebei Province / Center for Agricultural Resources Research,Institute of Genetics and Developmental Biology,Chinese Academy of Sciences,Shijiazhuang 050022,China;2.University of Chinese Academy of Sciences,Beijing 100049,China;3.College of Biologic and Geographic Sciences,Qinghai Normal University,Xining 810000,China;4.Hebei Normal University Attached School,Shijiazhuang 050011,China)
Photosynthesis can reveal the response of plant to the environment.By fitting the light response curve,the physiological mechanism of plant under different growth conditions could be clarified.Under four treatments of two CO2concentrations (400 and 750 μmol·mol-1) and two soil water contents [45% (drought stress) and 75% (suitable water condition) of fieldcapacity],Li-6400 portable photosynthetic system was used to measure the light response curves of flag leaves of winter wheat.Five photosynthetic light response models,including rectangular hyperbolic model (RHM),non-rectangular hyperbolic model(NRHM),rectangular hyperbolic modified model (RHMM),exponential model (EM) and modified exponential model (MEM),were used to fit the light response curve of winter wheat.The effects on photosynthesis of winter wheat of different treatments were analyzed.The results showed that the light saturation point (LSP) and the max net photosynthetic (Pnmax) fitted by the rectangular hyperbolic model and the non-rectangular hyperbolic model were larger than the measured values.The rectangular hyperbolic model and the non-rectangular hyperbolic model could not fit the photoinhibition.Meanwhile,the exponential model could simulate well of LSP andPnmaxwhile could not show the photoinhibition,either.Although the modified exponential model could reveal the photoinhibition,there were serious bias in the parameters of light response curve,and the parameters were lower than the other models.In general,the rectangular hyperbolic modified model was the best model for the simulation of light response curve and its parameters.The increase of CO2concentration effectively improved the apparent quantum efficiency,light saturation point and maximum net photosynthetic rate of winter wheat,and reduced the light compensation point and dark respiration rate of winter wheat.Meanwhile,winter wheat energy conversion rate,maximum net photosynthetic rate,light use scope and initial photosynthetic efficiency were increased,which suggested that the increase of CO2concentration could effectively enhance the photosynthetic capacity of winter wheat.When the soil moisture content was low,the effects of drought stress was partly offset by increased initial photosynthetic efficiency of winter wheat,but the apparent quantum efficiency,light saturation point and maximum net photosynthetic rate were reduced obviously than the appropriate soil moisture content,namely the photosynthetic ability of winter wheat decreased significantly under drought stress.In addition,the increase of CO2concentration had a certain compensation effect on the decrease of photosynthesis caused by drought stress,and the enhancement effects of CO2concentration increase on photosynthetic capacity of winter wheat under drought stress was greater than that under appropriate soil moisture content.
Winter wheat;CO2concentrations enhancement;Drought stress;Photosynthetic light-response model;Photosynthetic capacity
Dec.23,2016;accepted Feb.26,2017
S311
:A
:1671-3990(2017)07-1034-08
10.13930/j.cnki.cjea.161167
周晓旭,张玉翠,吴林,沈逸群,姜寒冰,沈彦俊.冬小麦光合特性对CO2浓度与土壤含水量的响应机制[J].中国生态农业学报,2017,25(7):1034-1041
Zhou X X,Zhang Y C,Wu L,Shen Y Q,Jiang H B,Shen Y J.Response mechanism of photosynthetic characteristics of winter wheat to CO2concentration and soil water content[J].Chinese Journal of Eco-Agriculture,2017,25(7):1034-1041
*国家重点研发计划课题(2016YFC0401403)和国家自然科学基金项目(91425302,41471027,31400375)资助
** 通讯作者:张玉翠,主要从事生态水文方面研究。E-mail:yczhang@sjziam.ac.cn
周晓旭,研究方向为生态水文学。E-mail:zhouxiaoxu14@mails.ucas.ac.cn
2016-12-23 接受日期:2017-02-26
*This study was supported by the National Key Research and Development Plan of China (2016YFC0401403) and the National Natural Science Foundation of China (91425302,41471027,31400375).
** Corresponding author,E-mail:yczhang@sjziam.ac.cn
猜你喜欢
中学生数理化·七年级数学人教版(2023年9期)2023-11-30 03:09:24
环境保护与循环经济(2021年7期)2021-11-02 08:10:54
哈尔滨轴承(2020年1期)2020-11-03 09:16:22
中国奶牛(2019年10期)2019-10-28 06:23:36
小学生学习指导(低年级)(2019年9期)2019-09-25 07:43:28
中学生数理化·七年级数学人教版(2019年4期)2019-05-20 10:03:44
电子制作(2018年23期)2018-12-26 01:01:22
中学生数理化·七年级数学人教版(2017年9期)2017-08-15 00:53:19
新课程·下旬(2017年7期)2017-08-14 11:12:50
西北林学院学报(2015年5期)2015-01-03 05:12:20
