
60Co-γ对海州常山扦插过程内源激素含量的变化影响
2017-07-03 12:32张莹婷王良桂华雅洁杨秀莲
山东林业科技 2017年6期
何 岭,张莹婷,王良桂,华雅洁,胡 蝶,王 欣,陈 敏,杨秀莲
(南京林业大学 风景园林学院,江苏 南京 210037)
海州常山(Clerodendrum trichotomum),别名臭梧桐、泡火桐、追骨风等。产辽宁、甘肃、陕西以及华北、中南、西南各地,常出现在海拔2400m以下的灌丛中。朝鲜、日本及菲律宾北部也有分布[1]。海州常山不仅是有待开发的观赏木本花卉,也是有生物油用和药用的经济作物[2]。近年来,海州常山关于海州常山的研究逐渐丰富,但目前关于海州常山育种研究罕见报道,这会制约海州常山大面积地开发及推广。
60Co-γ辐射是最广泛使用的辐射方法,射线的高能量诱导生物的遗传物质变异,并重新组合得到有利的突变体[3]。赵云龙[4]用60Coγ对大白杜鹃进行辐射处理,吲哚乙酸(IAA)、赤霉素(GA3)、玉米素核苷(ZR)含量降低,脱落酸(ABA)含量增加,较高含量的脱落酸使大白杜鹃产生矮化现象。刘春贵等[5]使用60Coγ射线辐射路易斯安那鸢尾种子,发现100 Gy辐射处理会促进种子萌发率和成苗率,可用于种苗生产。60Coγ辐射诱变材料主要有种子、胚、组培苗、愈伤组织、枝条 、幼苗等,枝条辐射后常嫁接嫁接于母株上,辐射后扦插较为少见。近年来,史玉敏等[6]研究60Co-γ辐射对桂花插穗的影响,结果表明桂花根形态特征、新生叶的可溶性蛋白质和可溶性糖含量与辐照剂量呈负相关。
插穗内源激素的动态变化与生根有密切关系[7]。增加60Co-γ辐射剂量会提高植株的变异率,同时也会提高植株死亡率。笔者在试验研究中发现,非过高剂量的60Co-γ辐射并不会明显降低死亡率,而是延缓插穗物候期,导致生根前插穗内部营养物质耗尽死亡。本研究用不同剂量60Co-γ射线辐射海州常山枝条,测定插穗萌发过程中ABA、GA3、ZR、IAA 4种内源激素含量的动态变化和比值大小,旨在研究60Co-γ辐射后插穗生根与内源激素含量之间的关系,为今后辐射育种、施用外源激素方法以提高成活率提供一定的理论依据。
1 材料与方法
1.1 试验材料
在2017年3月6日,于南京林业大学白马教学科研基地采取生长健壮、无病虫害一年生休眠枝条(芽鳞开始松动)为试验材料。将枝条剪成长度为10cm左右的插穗,切口上平下斜,上切口离最上芽为1cm,下切口离下芽为0.5cm,脱脂棉包裹保湿。
1.2 试验方法
1.2.1 辐射处理与扦插试验
于2017年3月7日,经江苏省农业原子能农业利用研究所进行60Co-γ射线辐射处理,剂量15Gy,剂量率为1.1 Gy.min-1。同时设未经辐射处理的为对照(CK),每处理60根插穗,3个重复。扦插前,用400mg/LNAA溶液浸泡插穗基部1h[8],后插于基质中,基质配方为V泥炭:V珍珠岩:V蛭石:V河沙=1:1:1:2,基质经 800 倍多菌灵溶液消毒。 后日常养护管理。
1.2.2 内源激素的测定
考虑到物候及辐射效应,于扦插之日起,分别隔 40d、47d、54d、64d、74d、84d、104d 于每个处理组中随机选择10根插穗,洗净擦干,削取插穗基部1~2cm皮部,称量分装后存放于-80℃冰箱中备用,最后委托中国农业大学化控中心统一测定。各时期样品测定的内源激素含量测定采用酶联免疫吸附法(ELISA),激素具体包括吲哚乙酸 (IAA)、赤霉素(GA3)、玉米素核苷(ZR)和脱落酸(ABA)。
1.3 数据分析
用Excel 2003软件数据分析与制图。
2 结果与分析
2.1 60Co-γ对扦插生根过程中激素含量变化的影响
2.1.1 生长素(IAA)
由图1表明,扦插后60Co-γ射线15Gy辐射的插穗IAA含量呈现先下降再上升最后再下降的趋势。对照处理的插穗扦插40-47d内呈下降趋势,后于47-54d时,上升至125.99ng.g-1,扦插54d后不定根逐渐伸长,对照处理的IAA含量逐渐下降。而60Co-γ射线处理的插穗IAA含量在扦插40-54d缓慢上升;54-64d为辐射处理插穗愈伤组织分化成根原基,IAA含量下降;64-104d随着根尖的生长素的合成,IAA含量呈上升的变化趋势。辐射处理插穗IAA含量谷值比对照延迟17d,两者IAA含量分别为 107.93ng.g-1、91.31ng.g-1。
2.1.2 细胞分裂素(玉米素ZR)
由图1可见,ZR在海州常山插穗生根过程中总体呈下降上升的趋势。对于对照处理的插穗在不定根诱导期40-64d时,ZR含量下降,扦插64d后,随着不定根的伸长,ZR含量上升后又趋于稳定;经辐射处理的插穗,40-74d为不定根诱导期,ZR含量在74d达到最低值,后不定根伸长生长,ZR含量逐渐上升,其中辐射处理ZR含量的最低值出现期比对照处理组晚10d。
2.1.3 赤霉素(GA3)
海州常山插穗扦插过程中,GA3含量变化趋势为先上升后降低。在扦插前期40-47d,GA3含量无明显变化,且对照组与处理组无明显差距,扦插后47-54d,GA3含量均急剧上升,对照与处理水平GA3含量分别达到22.22ng.g-1和17.62ng.g-1。64d后GA3含量又降至扦插初期水平,且对照组比处理组变化幅度大。试验中,辐射处理组与对照组不定根形成时间不同,但GA3峰值出现在相同的时间段。
2.1.4 脱落酸(ABA)
根据图1可知处理组和对照组插穗的ABA含量总体呈下降趋势。40-64d不定根诱导期间对照组ABA含量持续降低,最低值为117.44ng.g-1;64-104d插穗内ABA含量有一定的上升。60Co-γ射线辐射处理的插穗ABA含量在40-64d差异不大,可能是辐射引起的胁迫作用使得ABA含量处于一个较高的稳定水平;64-74d时,愈伤组织形成不定根ABA含量下降,随后下降幅度有所减缓,84d后ABA含量再次降低,根系持续伸长生长。
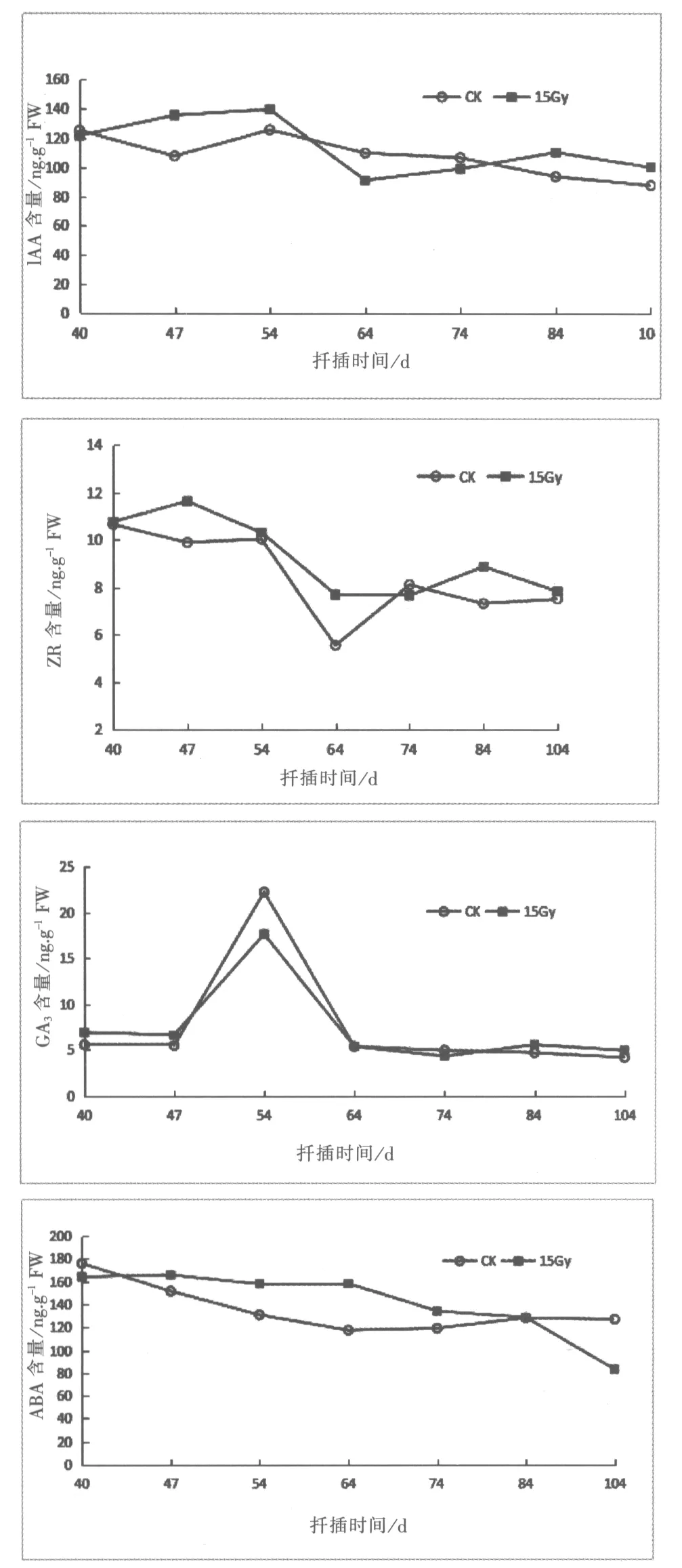
图1 60Co-γ射线辐射后不同内源激素含量的变化Fig.1 Variations of different endogenous hormones content treated by 60Co-γradiation
2.2 60Co-γ对扦插生根过程激素的比值变化
2.2.1 生长素与脱落酸(IAA/ABA)
如图2所示,60Co-γ射线辐射处理的插穗IAA/ABA比值呈先上升再下降最后再上升的趋势,而对照组表现出先上升再下降的变化趋势。辐射处理的IAA/ABA比值在40-54d呈上升趋势,在这可能是不定根诱导期IAA含量升高和ABA含量降低引起的,随着不定根的出现比值随IAA含量降低而降低;64-104d,IAA/ABA比值呈上升的趋势。对照处理组插穗IAA/ABA含量在40-54d呈上升变化趋势,最高达0.96ng.g-1;54d后比值逐渐降至扦插前期水平。取样期间,辐射处理组插穗IAA/ABA比值多低于对照组处理水平。
2.2.2 赤霉素与脱落酸(GA3/ABA)
由图2可见,海州常山插穗GA3/ABA比值变化趋势基本相同,GA3/ABA在插穗生根过程中呈先上升后降低的趋势。GA3/ABA比值在扦插40-47d差距不大,但在54d辐射组和对照组插穗GA3/ABA比值同时急剧上升,分别达到0.17ng.g-1和0.11ng.g-1。扦插64d,比值下降至扦插前期水平;74-104d,处理组比值呈缓慢上升趋势,而对照组变化趋势与其相反。
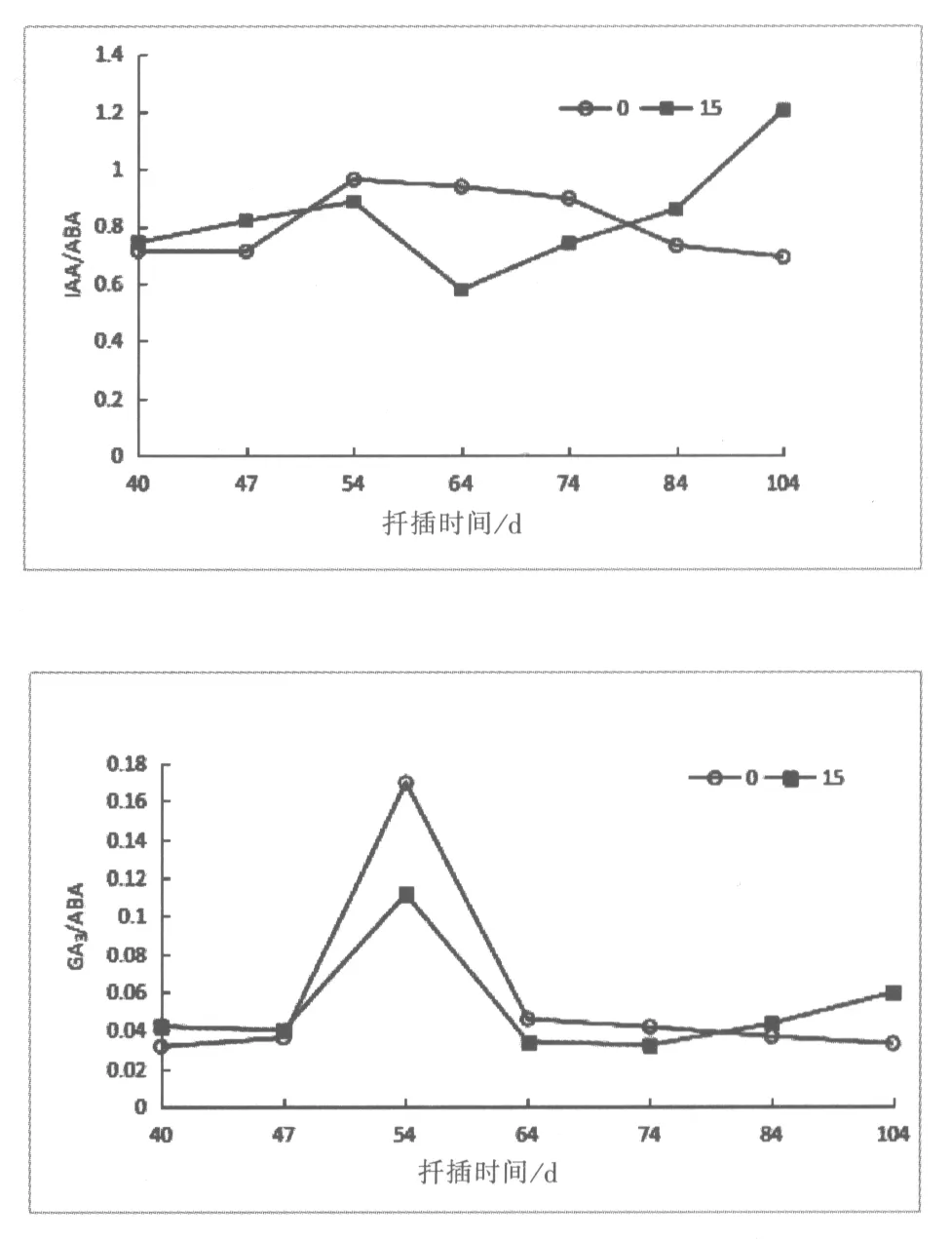
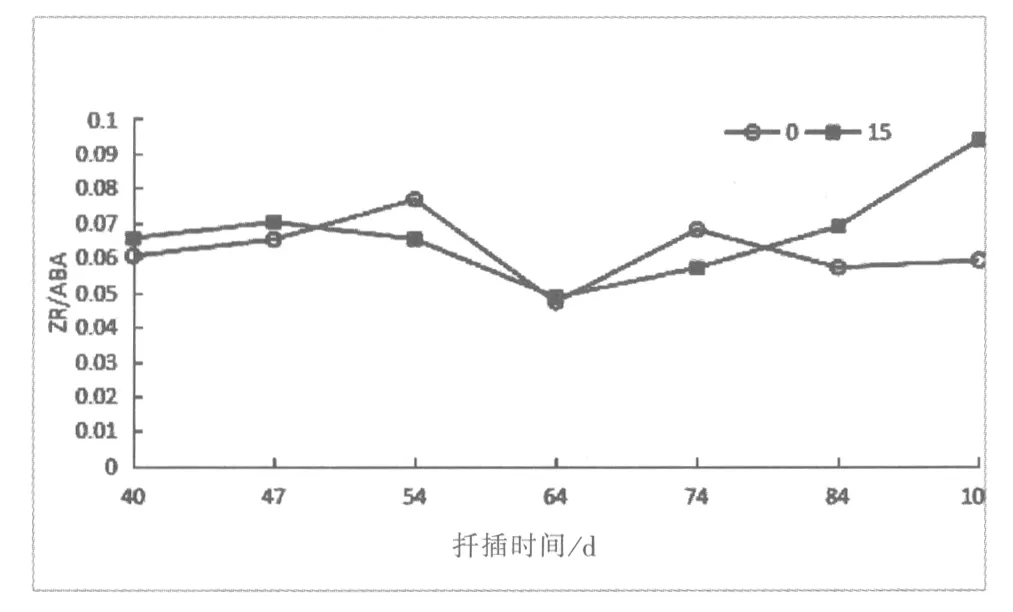
图2 60Co-γ射线辐射后IAA/ABA,GA3/ABA和ZR/ABA的变化Fig.1 Variations of IAA/ABA,GA3/ABA and ZR/ABA treated by60Co-γradiation
2.2.3 细胞分裂素与脱落酸(ZR/ABA)
由图2分析可知,辐射组比值ZR/ABA呈现出先下降后上升的趋势,对照组比值变化趋势由上升到下降再至上升最后下降的变化趋势。在不定根伸长前,40~64d内ZR/ABA比值下降,在64d降至最低0.049ng.g-1,之后一直呈上升趋势。对照组ZR/ABA比值于扦插后40~54d内逐渐升高,达到0.076ng.g-1,此时正值不定根形成时期。扦插54d-64d比值急剧下降,随着不定根的发育,比值上升继而下降至扦插前期水平。
3 结论与讨论
60Co-γ射线辐射会对植物产生有益的突变效应,同时也会对植物DNA、生物膜系统、酶系统产生伤害。杨振等[9]以库尔勒香梨为试验材料,采用60Coγ射线进行处理,发现辐射会抑制枝条的萌发和生长,但随着辐射效应的降低,库尔勒香梨枝条会恢复正常生长。同时,在扶芳藤[10]荔枝[11]的研究中也有类似的发现,辐射影响激素的产生,这与植物的生长发育有着密切的关系[12]。本研究中,海州常山插穗经15Gy的60Co-γ射线辐射后,不定根的生长较对照组延迟了10d。在辐射育种过程中,插穗生根前应加强日常养护管理,提供充足的水肥,以保证辐射材料的成活率。
前人的研究多证明生长素是促进生根的因子[13-16]。本研究显示,在海州常山插穗生根过程中,激素与扦插生根关系密切,不定根形成前内源IAA含量下降。不定根形成后IAA含量上升。不定根形成前根原基的诱导消耗大量内源IAA,不定根形成后随着根尖合成新的IAA,IAA含量上升以保持根系的生长。对照组和辐射处理组在IAA含量谷值上差距17d,辐射处理伤害了插穗定芽合成IAA的功能,从而延缓插穗的物候期。
ZR属于细胞分裂素,对细胞分裂有促进效应,也可以延缓细胞衰老、解除顶端优势等[17]。一些研究表明,高浓度细胞分裂素会抑制插穗不定根的生成,而低水平有利于生根[18]。本试验中,海州常山插穗生根在不定根诱导及生长初期ZR含量低,表明内源ZR的降低有利于不定根的形成。随着根系的生长,ZR含量的上升促进不定根分化。60Co-γ射线辐射延长了内源ZR含量变化趋势,表明辐射延缓了插穗不定根的形成。
赤霉素可以促进植物节间伸长与种子萌发,并且与植物开花结果密切相关。目前内源GA3含量与插穗生根关系尚未有明确定论。前人在树牵牛[19]的研究中发现,GA3可显著提高植株生根率,促进根系生长。但闫文涛等[20、21]认为内源GA3活性降低有利于插穗不定根原基的诱导,低水平GA3能促进植物根尖发育。本试验中,插穗不定根形成前,GA3含量均有个骤升过程,这代表不定根的形成需要内源GA3的促进。
经过60Co-γ处理后,海州常山扦插生根过程中辐射处理插穗ABA含量高于对照值。ABA含量的降低表明低含量的ABA有助于插穗生根,这与前人[22、23]的研究结论相似。辐射易引起ABA含量上升,内源ABA会通过影响生长素在插穗内的分布和含量,进而影响根尖分生区细胞分裂,从而抑制不定根伸长[24]。
植物扦插过程中,不同内源激素的含量和变化的动态平衡是插穗生根机理,内源激素含量比值与不定根生成的难易程度密切相关[25-27]。一般认为I-AA/ABA高,有助于根原基诱导分化[25,28]。本研究发现,60Co-γ射线辐射处理插穗的 IAA/ABA,GA3/A-BA,ZR/ABA比值在不定根诱导分化期均低于对照值,多在扦插54d比值达到峰值,于64d达到谷值以利于不定根生成,这与杨雪萌等[29]对菊花品种天坠玉露及杨霄霞[27]对香椿的研究结论相似。3者比值变化趋势与扦插生根趋势大体吻合,内源激素变化节奏与生根机理之间是协同的,但辐射会降低插穗内促进生根激素的含量,刺激抑制激素的生成,从而延缓生根。
综上所述,60CO-γ射线辐射会通过调控几种激素含量和比值的变化来抑制海州常山插穗生根。在今后的研究工作中,使用合适的促进生根激素对海州常山插穗进行处理,可提高辐射育种材料的成活率。
[1]中国科学院中国植物志委员会,裴鉴,陈守良,等.中国植物志:马鞭草科[Z].北京:科学出版社,1982.
[2]杨秀莲,林燕青,王良桂.海州常山花粉低温贮藏及其生理生化特征研究 [J].西北植物学报,2015,35(9):1808-1814.
[3]李志能,刘国锋,包满珠.悬铃木种子~(60)Coγ辐照及其苗期生物学性状调查 [J].核农学报,2006,20(4):299-302.
[4]赵云龙,李朝婵,陈训.~(60)Co-γ射线辐射对大白杜鹃的矮化效应[J].北方园艺,2012(19):48-50.
[5]李风童,包建忠,孙叶,等.~(60)Co-γ射线辐照德国鸢尾杂交种子的生物效应 [J].核农学报,2017,31(8):1469-1474.
[6]史玉敏,罗先真,严恒,等.60Co-γ射线辐照对桂花枝条生长和生理指标的影响及耐辐照性评价 [J].核农学报,2017,31(2):350-356.
[7]赵云龙,陈训,李朝婵.糙叶杜鹃扦插生根过程中生理生化分析[J].林业科学,2013,49(6):45-51.
[8]刘军,曾德静,王铖,等.植物生长调节剂对海州常山硬枝扦插生根的影响 [J].东北林业大学学报,2012,40(8):10-12.
[9]杨振,李疆,梅闯,等.~(60)Co-γ辐照对库尔勒香梨枝条当代诱变效应初报 [J].新疆农业科学,2012,49(5):848-855.
[10]敖妍,张国盛,鲁韧强,等.扶芳藤种子与枝条的60Coγ 辐射效应[J].核农学报,2006,20(3):202-204.
[11]秦献泉,徐宁,朱建华,等.荔枝枝条辐射生物学效应研究初报[J].种子,2016,35(7):5-9.
[12]郭少玲,高健,徐有明,等.60Coγ辐射对毛竹种子萌发过程中内源激素含量的影响 [J].中国农学通报,2013,29(25):26-31.
[13]Dey K,Ghosh A,Dey A N,et al.Efficacy of IAA con-centration and cutting length on rooting of stem cuttings in Spondias pinnata Linn[J].Indian Journal of Horticul-ture,2017,74(1):127.
[14]Dong N,Pei D,Yin W.Tissue-specific localization and dynam ic changes of endogenous IAA during poplar leaf rhizogenesis revealed by in situ immunohistochem-istry[J].Plant Biotechnology Reports,2012,6(2):165-174.
[15]吴文浩,曹凡,刘壮壮,等.NAA对薄壳山核桃扦插生根过程中内源激素含量变化的影响 [J].南京林业大学学报(自然科学版),2016,40(5):191-196.
[16]孙晓梅,韩华,王笑山.不同株龄日本落叶松插穗的内源激素含量与生根的关系[J].植物生理学报,2009,45(3):217-222.
[17]李合生.现代植物生理学 [M].高等教育出版社,2012.
[18]郭素娟,凌宏勤,李凤兰.白皮松插穗生根的生理生化基础研究[J].北京林业大学学报,2004,26(2):43-47.
[19]Nanda K K,Anand V K,Chibbar R N.The promotive effect of gibberellic acid on the production of adventi-tious roots on stem cuttings of Ipomoea fistulosa.[J].Planta,1972,105(4):360.
[20]闫文涛,佟兆庆,魏俊,等.不同生长调节剂对蒙古栎嫩枝扦插生根的影响 [J].西北林学院学报,2017,(3):116-121.
[21]董胜君,刘明国,戴菲,等.山杏嫩枝扦插生根过程中插穗内源激素含量的变化 [J].经济林研究,2013,31(4):108-114.
[22]麻文俊,张守攻,王军辉,等.日本落叶松扦插生根期内源激素和营养物质及酚酸含量变化特征 [J].西北植物学报,2013,33(1):109-115.
[23]马振华,王吉斌,李才文,等.四倍体刺槐硬枝扦插生根过程中内源激素的变化 [J].中南林业科技大学学报,2013,28(6):18-23.
[24]袁冰剑,张森磊,曹萌萌,等.脱落酸通过影响生长素合成及分布抑制拟南芥主根伸长 [J].中国生态农业学报,2014,22(11):1341-1347.
[25]敖红,王崑,冯玉龙.长白落叶松插穗的内源激素水平及其与扦插生根的关系 [J].植物研究,2002,22(2):190-195.
[26]董胜君,刘明国,戴菲,等.山杏嫩枝扦插生根过程中插穗内源激素含量的变化 [J].经济林研究,2013,31(4):108-114.
[27]杨霄霞,潘晓芳.香椿插穗内源激素变化与生根关系[J].广西林业科学,2015,44(2):132-136.
[28]]Kelen M, Ozkan G. Relationships Between RootingAbility and Changes of Endogenous IAA and ABA Dur-ing the Rooting of Hardwood Cuttings of someGrapevine Rootstocks[J]. European Journal of Horticul-tural Science, 2003, 68(1):8-13.
[29]杨雪萌,房伟民,陈发棣,等.两个菊花品种扦插生根过程及其插穗碳氮营养和内源激素的变化 [J].南京农业大学学报,2010,33(4):19-24.
猜你喜欢
作物研究(2022年1期)2022-11-27
作物学报(2022年10期)2022-07-21
小学生优秀作文(低年级)(2022年3期)2022-03-29
中国现代中药(2022年12期)2022-02-18
现代农业研究(2021年10期)2021-11-02
连云港师范高等专科学校学报(2021年4期)2021-03-04
连云港师范高等专科学校学报(2021年4期)2021-03-04
传媒评论(2019年4期)2019-07-13
江苏教育(2019年8期)2019-06-12
天然产物研究与开发(2018年4期)2018-05-07
