
不同年份30 d短日照处理对地被菊开花的影响
2017-06-28 16:35:52杨中义王晨李艳锋张春来吕晋慧
山西农业大学学报(自然科学版) 2017年6期
杨中义,王晨,李艳锋,张春来,吕晋慧*
(1.山西农业大学 林学院,山西 太谷 030801; 2.山西农业大学 农学院,山西 太谷 038301)
不同年份30 d短日照处理对地被菊开花的影响
杨中义1,王晨1,李艳锋1,张春来2,吕晋慧1*
(1.山西农业大学 林学院,山西 太谷 030801; 2.山西农业大学 农学院,山西 太谷 038301)
[目的]揭示30 d短日照处理调控菊花花期的机理。[方法]比较不同年份间30 d短日照处理下地被菊花芽分化及开花进程的差异。[结果]2014年出现二次开花现象,第一次开花始花期比对照提前43 d,2015年开花一次,花期与对照一致;株高和冠幅显著小于对照,不同年份间差异不显著,开花量2014年显著高于2015年;2014年df(第一次开花)可溶性蛋白含量在露色期、始花期、盛花期显著高于对照,2015年各开花阶段显著高于对照,不同年份间在现蕾期、始花期、盛花期和末花期差异显著;2014年可溶性糖含量在露色期、始花期显著低于对照,其中2014年df在现蕾期、盛花期显著高于对照,2015年在现蕾期、盛花期显著高于对照,露色期、末花期显著低于对照,不同年份间可溶性糖含量差异显著;花芽分化2014年较2015年提前6 d启动分化,提前43 d完成分化。[结论]短日照抑制植株株高、冠幅等营养生长,同时影响菊花花芽分化及开花进程。30 d短日照是菊花花芽分化和开花的敏感时期,条件适宜时植株提前完成花芽分化并开花,不适宜时不能诱导花芽分化或出现成花逆转。
短日照处理; 花芽分化; 可溶性蛋白; 可溶性糖
地被菊(Chrysanthemummorifolium)开花繁茂、抗逆性强,耐粗放管理,但花期主要集中在秋季。目前,通过短日照处理调控花期是菊花生产中主要采用的技术措施[1]。吴文新等[2]发现,欲使花期为10月下旬至 11 月的菊花在国庆节开放,可于立秋开始遮光处理,每日遮光14 h,45 d左右即可开花。李申军等[3]认为切花菊短日照处理21~28 d可提前开花,多花型菊花则需要更长时间的短日照;刘慧琴等[4]研究表明,切花菊“神马”在短日照下,花蕾形成需要20多天,花朵露白需要50 d左右;邝琦等[5]认为,从短日照至开花,切花小菊需要6~10周,大菊品种需要9~12周;杨娜等[6]的研究结果表明,切花菊‘神马’在短日照下处理23 d就能完成从营养生长到生殖生长的转变,而‘秀芳之力’则需要30 d[7];赵耀等[8]对地被菊进行了光周期调控研究,认为光周期处理天数与不同菊花品种的花期有关,花期越晚的品种,需要光周期处理的时间越长,反之则短。而花芽分化和开花是复杂的形态建成过程,是花卉体内各种因素与外界环境相互协调共同调控的结果[9]。以上研究均着眼于短日照处理对花期的影响,本研究团队发现不同年份间30 d短日照处理后植株花芽分化及开花进程出现较大差异,因此本研究主要从可溶性蛋白、可溶性糖含量等方面入手,分析造成较大差异的原因,进一步揭示短日照对菊花花期的调控机理。
1 材料与方法
1.1 材料
供试材料为短日照敏感型地被菊品种‘紫娇’,其株型紧凑、开花繁茂、花瓣紫色、重瓣性高,自然始花期在9月中旬,是观赏性较高的优良地被菊品种。试验于2014年、2015年5月在山西农业大学林学院中心实验室与苗圃进行。
1.2 方法
1.2.1 短日照处理
选取苗龄相同且长势良好、一致的‘紫娇’扦插苗定植于大田,正常水肥管理。设置2个短日照处理:0 d(对照)、30 d,每个处理30株。6月15日开始,每日18:00至次日8:00,以黑色涂荧遮光布遮光处理14 h。观察植株生长状况并记录相关指标。物候期观测指标包括:(1)花期。其中现蕾期:花蕾直径1~2 mm。露色期:透过花蕾总苞可看到花朵颜色。始花期:10%~30%的花序完全开放。盛花期:50%~70%的花序完全开放。末花期:30%花序出现花色褪色、衰败现象。(2)形态指标。株高:盛花期测量植株生长点与地面间距离。冠幅:盛花期用十字法测量。开花量:盛花期测定植株单株花朵数量。
1.2.2 不同年份30 d短日照处理对‘紫娇’可溶性蛋白和可溶性糖含量的影响
每日上午9:00-11:00随机取植株茎中上部健康叶片。在短日照处理前一天对各处理统一取样一次,从处理开始每隔4 d采样一次,至末花期停止。其中用于测定可溶性糖的材料80 ℃烘干24 h备用。分别采用蒽酮比色法与考马斯亮蓝 G-250染色法进行可溶性糖、可溶性蛋白含量的测定[10],3次重复。
1.2.3 不同年份30 d短日照处理对‘紫娇’花芽分化的影响
从6月8日开始,每隔2 d采样一次,每次随机取10个顶芽,用于制作石蜡切片,荧光显微镜下观察并记录花芽分化时期[4]。
1.2.4 数据统计与分析
使用Excel 2010统计整理数据,SPSS 17.0进行方差分析与多重比较(LSD法)。显著性差异水平为P<0.05,用小写字母标注表示。
2 结果与分析
2.1 不同年份30 d短日照处理对‘紫娇’花期的影响
2014年和2015年30 d短日照处理下‘紫娇’花期差异性明显。2014年30 d短日照处理下出现两次开花现象,而2015年开花一次(表1),说明除短日照外,温度、水分和营养物质等多种因素极有可能影响菊花花期。其中2014年30 d短日照处理下,8月4日第一次开花(30 df),现蕾期、露色期、始花期分别比对照提前44 d、54 d、43 d。30 d短日照处理结束后第8天(7月23号)出现成花逆转,第41天(8月25日)恢复正常生长,这部分花蕾9月16日第二次开花(30 ds);2015年7月22日进入现蕾期,较对照提前10 d,较2014年30 df推迟24 d;9月10日进入露色期,较对照提前5 d,较2014年30 df推迟58 d;始花期与对照一致,较2014年30 df均推迟47 d。8月2日出现成花逆转,并于9月5日恢复正常生长,没有二次开花现象。

表1 2014年和2015年30d短日照处理下‘紫娇’花期差异比较
注:df:第一次开花;ds:第二次开花,下同。
Note: df: the first time flowering of 30d short-day treatment; ds: the second time flowering of 30d short-day treatment; similarly hereinafter.
2.2 不同年份30 d短日照处理对‘紫娇’生长发育状况的影响
‘紫娇’不同年份30 d短日照处理下,植株株高、冠幅显著低于对照,且两年间差异不显著(图1-A,B),说明短日照是影响株高、冠幅的关键因素;2014年短日照处理后植株开花量与对照无显著差异,但显著高于2015年(图1-C),说明温度、水分和营养物质等多种因子极有可能影响植株开花量。

图1 2014年和2015年30 d短日照处理下地被菊'紫娇'生长发育的差异Fig.1 Difference on plant growth and development of 30 d short-day treatment
2.3 不同年份30 d短日照处理对‘紫娇’可溶性蛋白含量的影响
随着花期各阶段的推进,2014年30 df植株的可溶性蛋白含量呈先升后降趋势,始花期含量最高;30 ds植株的可溶性蛋白含量呈先降后升趋势,盛花期含量最低,而对照可溶性蛋白含量总体呈下降趋势。2014年30 df在露色期、始花期、盛花期和末花期植株可溶性蛋白含量显著高于对照,其中露色期、始花期、盛花期对照可溶性蛋白含量显著高于30 ds(图2-A)。
2015年30 d短日照处理和对照植株的可溶性蛋白含量均呈先降后升趋势,在盛花期含量最低。30 d短日照处理下植株各开花阶段可溶性蛋白含量显著高于对照(图2-B)。
现蕾期、盛花期和末花期,2014年30 df、ds可溶性蛋白含量显著低于2015年;露色期2014年30 df可溶性蛋白含量与2015年无明显差异,显著高于30 ds;始花期2014年30 df显著高于2015年和30 ds(图2-C)。
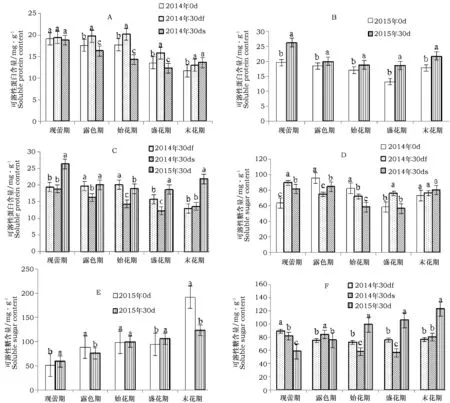
图2 2014年和2015年30 d短日照处理下地被菊'紫娇'可溶性蛋白、可溶性糖含量的差异Fig.2 Difference of soluble protein content and soluble sugar content of 30 d short-day treatment between 2014 and 2015
2.4 不同年份30 d短日照处理对‘紫娇’可溶性糖含量的影响
2014年30 df植株的可溶性糖含量呈先降后升趋势,30 ds和对照植株呈升降升的趋势,且在盛花期含量最低。30 d短日照处理下末花期植株可溶性糖含量与对照无显著差异,其余各阶段差异性显著,其中现蕾期、盛花期2014年30 df可溶性糖含量显著高于30 ds和对照,露色期、始花期2014年30 df、ds显著低于对照(图2-D)。
2015年30 d短日照处理植株可溶性糖含量呈上升趋势,对照植株则呈升降升趋势,均在末花期含量最高。30 d短日照处理下,始花期阶段植株可溶性糖含量与对照一致,其它开花阶段植株可溶性糖含量与对照差异显著,其中现蕾期、盛花期显著高于对照,露色期、末花期显著低于对照(图2-E)。
除露色期外,2014年30 df,ds和2015年30 d短日照处理可溶性糖含量差异显著。2015年30 d短日照处理可溶性糖含量,露色期显著低于2014年30 ds,与30 df无显著差异;现蕾期显著低于2014年30 df,ds;其它时期则显著高于2014年30 df,ds(图2-F)。
2.5 不同年份30 d短日照处理对‘紫娇’花芽分化进程的影响
‘紫娇’花芽分化进程为:(Ⅰ)未分化期。(Ⅱ)花芽分化启动期。(Ⅲ)总苞鳞片分化期:包括总苞鳞片分化初期和末期。(Ⅳ)小花原基分化期:包括小花原基分化初期和末期。(Ⅴ)花冠形成期:分为花冠形成初期、中期和末期。
2014年30 d短日照处理和对照均于6月17日启动分化,对照于8月28日花芽分化完成,30 d短日照处理于8月10日第一轮花芽分化完成,之后部分顶芽再次进入筒状小花分化盛期,并于8月31日完成第二轮分化。
2015年30 d短日照处理下,于6月23日启动分化,总苞鳞片分化末期、小花原基分化初期和小花原基分化末期分别较对照提前10 d、12 d、12 d,其它花芽分化阶段出现时期与对照一致,30 d短日照处理和对照于8月23日和26日分别完成分化。
2014年30 d短日照处理下各花芽分化阶段出现时期均早于2015年,其中2014年较2015年提前6 d启动分化,提前43 d完成分化(图3),说明除短日照外,温度、水分和营养物质等多种因素极有可能影响菊花花芽分化。

图3 2014年和2015年不同短日照处理紫娇花芽分化的差异Fig.3 Difference of floral bud differentiation of 30 d short-day treatment between 2014 and 2015
3 讨论
可溶性蛋白含量是植物重要的生理指标。花芽分化诱导前期,叶片蛋白含量有一定的积累,伴随花期的推进,蛋白质被不断运输至生长点。盛花期开花量大,所需蛋白质也较多,故叶片中可溶性蛋白含量急剧下降至最低。末花期开花数量减少,所需蛋白质减少,含量有所回升,这与李亚莉[11],刘志鹏等[12]等研究结果类似。盛花期和末花期,30 d短日照处理下2014年可溶性蛋白含量显著低于2015年,原因可能是2014年植株花期提前,营养生长时间短,积累可溶性蛋白少,同时,植株提前开花需要较多可溶性蛋白,8月25日成花逆转的花蕾恢复正常生长、第二次开花,需要更多的蛋白供给满足植株正常生长,综上,导致可溶性蛋白含量显著低于2015年。
可溶性糖是植物体内一种重要的化合物,其含量的高低反映了植物体内可利用态物质和能量的供应基础。花芽分化时期可溶性糖一部分作为呼吸基质被消耗,另一部分被转化为花芽分化中所需的营养物质[11]。本研究中,2014年各处理‘紫娇’可溶性糖含量整体表现出先下降,末花期再度回升现象,可能是由于花发育过程中可溶性糖被不断消耗,到末花期花器官衰老对营养消耗减少,叶片中可溶性糖积累增多,同时也为后期子房发育储备营养物质。而2015年30 d短日照处理下可溶性糖含量呈上升趋势,与刘志鹏等[12],钟晓红等[13],李兴军等[14]的研究结果一致。2015年‘紫娇’植株营养生长期长,光合作用时间长,积累碳水化合物多,同时,山西8月中旬后气温迅速下降,植株呼吸作用减弱,消耗碳水化合物少,因此,可溶性糖含量呈上升趋势。
菊花从营养生长到生殖生长需要一定时间,开花不但与自身的开花机理有关,还与外界环境密切相关[15],是由光照、温度、水分、营养状况和开花基因等各种因子互作、协同调控花期[9]。马月萍等[16]也认为植物分生组织由营养生长向生殖生长转变过程中,多种环境因子参与改变植物的成花反应。适于秋菊生长的温度是5 ℃~25 ℃,32 ℃以上则生长缓慢[17~21]。水分会影响植物的花芽分化和开花,适当干旱有利于植物花芽分化,连续阴雨天、天气湿度较大等都会延迟开花[22]。本研究中30 d短日照处理不同年份间花期差异较大,而50 d短日照处理下‘紫娇’不同年份间花期稳定[23,24],说明30 d短日照处理对‘紫娇’而言是较敏感的时期。除短日照外,温度、水分和营养物质等多种因素极有可能影响植株花芽分化,条件适宜时花芽分化顺利进行,花期提前,内外因素不适宜时,出现成花逆转或花芽分化推迟现象。本实验是在气象因素如温度、水分等非可控条件下进行的,不同年份间温度、水分及基因表达等的差异性导致了不同年份间花芽分化及开花进程差异较大,因此,本课题组今后将在可控条件下研究温度、水分和营养条件等对菊花花芽分化及开花的影响,同时,研究开花基因对菊花花期的影响。
4 结论
短日照抑制植株株高、冠幅等营养生长,同时影响菊花花芽分化及开花进程。30 d短日照是菊花花芽分化和开花的敏感时期,条件适宜时植株提前完成花芽分化并开花,不适宜时不能诱导花芽分化或出现成花逆转。
[1]倪月荷,汪觉先.菊花栽培与鉴赏[M].上海:上海科学技术出版社,2000:71-73.
[2]吴文新,王洪铭.菊花花期调控技术的研究概况及展望[J].福建农业科技,2001(3):21-23.
[3]李申军,王红艳,孙雅娟.菊花花期调控技术[J].吉林林业科技,2011,40(2):51.
[4]刘慧琴,章心惠,王宏航,等.出口切菊—日本“神马”花期调控和花梗长度控制[J].浙江农业科学,2006(6):642-643.
[5]邝琦,王文通,乔志钦,等.21个切花菊品种的光周期与花期调控研究[J].安徽农业科学,2011,39(7):3861-3866.
[6]杨娜,郭维明,陈发棣,等.光周期对秋菊品种“神马”花芽分化和开花的影响[J].园艺学报,2007,34(4):965-972.
[7]Zhang W, Fukai S, Goi M. Morphological observation on capitulum initiation and floret development ofDendranthemaspecies native to Japan[J]. Journal of the Japanese Society for Horticultural Sciences, 1996, 65(2): 506-507.
[8]赵耀,吕晋慧,陈阳,等.3种短日照依赖性地被菊光周期调控研究[J].山西农业大学学报(自然科学版),2013,33(1):22-26.
[9]宁云芬,汪勐炜,贾蕊,等.遮光处理下一品红花芽分化的形态学观察[J].北方园艺,2016(5):73-76.
[10]李合生,孙群,赵世杰.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:184-185,194-197.
[11]李娅莉.不同光周期对山茶花影响的研究[D].雅安:四川农业大学,2005.
[12]刘志鹏,刘明,张孟韬.短日照诱导对小豆三个生理生化指标的影响[J].南方农业,2016,10(15):10-11.
[13]钟晓红,罗先实,陈爱华.李花芽分化与体内主要代谢产物含量的关系[J].湖南农业大学学报(自然科学版),1999,25(1)31-35.
[14]李兴军,李三玉,汪国云.杨梅花芽孕育期间叶片酸性蔗糖酶活性及糖类含量的变化[J].四川农业大学学报,2000,18(2):164-166.
[15]宛敏渭,刘秀珍.中国物候观测方法[M].北京:科学出版社,1979:42-44,49-52.
[16]马月萍,戴思兰.菊花花芽分化时间的初步研究[C].∥中国菊花研究论文集,2002:124-126.
[17]石万里,姚毓璆.菊花花芽分化初步研究[J].园艺学报,1990,17(4):309-312.
[18]王希,朱秀珍,王彩云.短日处理对菊花开花及形态指标的影响[J].安徽师范大学学报(自然科学版),2013,36(1):60-63.
[19]ABDUL MK, SIMON P. Spectral filters and temperature effects on the growth and development of chrysanthemums under low light integral[J].Plant Growth Regulation, 2006,49(1):61-68.
[20]COCK shull K E, KOFRANEK A M. High night temperatures delay flowering produce abnormal flowers and retard stem growth of cut-flower chrysanthemums[J]. Scientia Hort, 1994, 56(3):217-234.
[21]HAHN E J, CHO Y R, LEE Y B. Air temperature and relative humidity affect the growth of chrysanthemum plantlets in the microponic system[J]. Journal of the Korean Society for Horticultural Science, 1998, 39(5):625-628.
[22]陈超,夏春华.海南地区年菊花期的调控技术[J],热带农业科学,1998(6):59-65.
[23]赵耀.早花期地被菊杂交育种研究[D].太谷:山西农业大学,2013.
[24]王晨.短日照处理对地被菊‘紫娇’花芽分化及开花的影响[D].太谷:山西农业大学,2016.
(编辑:韩志强)
Studied the influence of 30d short-day treatment on flowering of Ground-cover Chrysanthemum between different years
Yang Zhongyi1, Wang Chen1, Li Yanfeng1, Zhang Chunlai2, Lv Jinhui1*
(1.CollegeofForestry,ShanxiAgriculturalUniversity,Taigu030801,China; 2.CollegeofAgriculture,ShanxiAgriculturalUniversity,Taigu030801,China)
[Objective] Reveal the mechanism of 30d short-day treatment to control the florescence of Chrysanthemum. [Method] The differences of 30d short-day treatment on flower bud differentiation and flowering of Chrysanthemum were compared between different years. [Results] There was a secondary flowering phenomenon in 2014, and the early flowering stage was earlier 43 d than control in first time flowering. Only flowering one in 2015, the flowering was same with control. Plant height and crown were significantly lower than control, but the differences were non-significant in different years. And the flowers amount of 2014 was significantly higher than 2015. The soluble protein contents of 30 df in 2014 were significantly higher than control in visible flower color stage, early flowering stage and full bloom flowing stage; the contents of all flowering stages were significantly higher than control in 2015; the contents were significantly difference in visible flower bud stage, early flowering stage, full bloom flowing stage and later flowering stage between different years. The soluble sugar contents of 2014 were significantly lower than control in visible flower color stage and early flowering stage, and them of 30 df were significantly higher than control in visible flower bud stage and full bloom flowing stage; the contents of 2015 were significantly higher than control in visible flower bud stage and full bloom flowing stage, and lower than control in visible flower color stage and later flowering stage; the soluble sugar contents were significantly difference in different years. Flower bud differentiation started in 2014 earlier 6d than 2015, and earlier 43 d completed flower bud differentiation. [Conclusion] Short-day treatment inhibits the development in plant height and crown, and influences the flower bud differentiation and flowering of Chrysanthemum. 30 d short-day is the sensitive period to flower bud differentiation and flowering of Chrysanthemum. When plants grow in a suitable condition, they will complete advanced flower bud differentiation and flowering, if not, they will not induce differentiation or appear flowering reversion.
Short-day treatment, Flower bud differentiation, Soluble protein, Soluble sugar
2016-12-28
2017-02-08
杨中义(1992-),男(汉),山西怀仁人,硕士研究生,研究方向:园林植物遗传育种
*通信作者:吕晋慧,教授,硕士生导师,Tel:0354-6288329;Email: 335493446@qq.com
国家“十二五”科技支撑计划课题(2013BAD01B07);国家自然基金项目(30972021);山西农业大学青年学科带头人及学术骨干计划项目(XG201213)
S688.4
A
1671-8151(2017)06-0419-06
猜你喜欢
气象水文海洋仪器(2022年2期)2022-07-08 09:48:48
农产品市场周刊(2022年4期)2022-05-21 15:15:28
农产品市场周刊(2022年4期)2022-03-23 22:34:20
金桥(2021年9期)2021-11-02 05:54:52
金桥(2021年9期)2021-11-02 05:54:30
落叶果树(2020年3期)2020-06-17 05:11:28
河北果树(2020年1期)2020-02-09 12:31:08
星星·散文诗(2017年2期)2017-07-05 11:34:16
小布老虎(2016年18期)2016-12-01 05:47:41
广东农业科学(2016年3期)2016-07-27 02:07:13
