
广西不同林龄杉木、马尾松人工林根系生物量及碳储量特征
2017-06-27 00:39曾馥平彭晚霞
生态学报 2017年7期
韩 畅,宋 敏,杜 虎,曾馥平,彭晚霞,王 华,陈 莉,苏 樑
1 湖南农业大学生物科学技术学院, 长沙 410128 2 湖南农业大学农学院, 长沙 410128 3 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 4 中国科学院环江喀斯特生态系统观测研究站, 环江 547100
广西不同林龄杉木、马尾松人工林根系生物量及碳储量特征
韩 畅1,3,4,宋 敏2,3,4,杜 虎3,4,曾馥平3,4,彭晚霞3,4,王 华1,*,陈 莉1,3,4,苏 樑1,3,4
1 湖南农业大学生物科学技术学院, 长沙 410128 2 湖南农业大学农学院, 长沙 410128 3 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 4 中国科学院环江喀斯特生态系统观测研究站, 环江 547100
为了解不同林龄杉木、马尾松人工林地地下根系生物量及碳储量特征,以广西杉木、马尾松主产区5个不同林龄阶段(幼龄林、中龄林、近熟林、成熟林、过熟林)的人工林为研究对象,采用全根挖掘法和土钻法获取标准木根系生物量、灌草根系生物量和林分细根生物量,并测定其碳含量,分析其不同林龄阶段地下根系生物量和碳储量分配特征。结果表明:杉木、马尾松林地下根系总生物量分别在9.06—31.40 Mg/hm2和7.91—53.40 Mg/hm2之间,各林龄阶段根系总生物量总体上呈现随林龄增加而增加的趋势,杉木林细根生物量随林龄的增加呈现出先减后增的趋势,马尾松呈现出逐渐减小的趋势;林分各层次根系碳含量表现为乔木灌木草本、细根;杉木、马尾松地下根系碳储量变化趋势与生物量变化趋势相同,杉木、马尾松林不同林龄阶段各层次根系和土壤细根总碳储量分别在7.56—21.97 Mg/hm2和8.86—29.95 Mg/hm2之间;地下根系碳储量总体上以乔木根系占优势,且随林龄的增大其比例呈增加的趋势。
碳储量;根系;林龄;杉木;马尾松
根系是植株适应陆地生活并对其生长起关键作用的重要器官,它不但为林木的地上部分提供固定和机械支撑作用,而且为植物吸收、输导养分和水分,通过呼吸和周转消耗光合产物,向土壤输入有机物质,在森林生态系统中物质循环和能量流动中扮演着重要角色[1- 3]。由于根系深埋地下,具有不易观察和测量的特性,其研究力度相对薄弱。20世纪中期以来,森林根系的研究逐渐受到关注,加深了人们对森林生态系统功能和效应的了解[4]。近年来,国内外学者围绕根系生态学、根系分布与结构、根系生物学、根系生理学等研究热点,展开了一系列研究[3,5- 7]。在根系生物量与碳储量方面,国外研究结果表明:根系生物量占总生物量的10%—20%,足见其重要性;而以森林蓄积数据资料为基础的森林生态系统碳储量估算存在较大的不确定性,常未包括林下植被、地表枯落物、根系以及土壤等组分的碳量,因此森林生态系统的碳储量计算和评估的研究需进一步深入[8- 10]。
杉木(Cunninghamialanceolate)和马尾松(Pinusmassoniana)均是我国南方广泛种植的树种,在我国森林资源构成中占据较大比例,在维持生态系统平衡方面具有重要地位[11-13]。20世纪后期以来,许多学者开展了杉木和马尾松人工林生物量、碳储量的研究,包括不同林分密度、不同演替阶段、不同经营管理措施、单一树种或混交林生物量结构特征、生产力水平及碳储量分配[14-18]。由于林木根系的难挖掘性,人们对地下根系的研究力度往往低于地上部分。且大量森林植物碳库研究主要以乔木层为主,对于下木层的研究相对薄弱。本研究选择广西杉木、马尾松主产区5个不同林龄阶段的人工林为研究对象,采用全根挖掘法和土钻法对标准木根系生物量、灌草根系生物量和林分细根生物量分别进行测定和研究,分析了杉木和马尾不同生长发育阶段地下根系生物和碳储量分配特征,以期为我国森林碳汇估算及森林生态系统碳循环研究提供科学依据,并为造林、森林生态、森林土壤学、森林立地等提供基础材料。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于广西壮族自治区(104°26′—112°04′E、20°54′—26°24′N),总面积23.67万km2,处于云贵高原东南边缘,两广丘陵的西部,南边朝向北部湾,整个地势为四周多山地与高原,而中部与南部多为平地,地貌为山地丘陵性盆地地貌,属于亚热带季风气候区,全区各地极端最高气温为33.7—42.5 ℃,极端最低气温为-8.4—2.9 ℃,年平均气温在16.5—23.1 ℃之间,各地年降水量均在1070 mm以上,大部分地区为1500—2000 mm,4—9月为雨季,其降水量占全年降水量的70%—85%。研究区植被类型多样,植物物种丰富,以常绿树种为主,植被区域主要为亚热带常绿阔叶林区域。广西全区森林面积达到1252.50万hm2,森林覆盖率52.71%,活立木总蓄积量为5.11亿m3,其面积和蓄积分别占全国的6.41%和3.42%。全区森林资源主要包括杉类、松类、柏类、桉类、栲类、青冈类、软阔类、经济林、竹类、城市森林、石山灌木林这11种类型,其中杉木、马尾松两种林型分别占广西区用材林总面积的64.83%,蓄积的78.34%。因此,选择这两种林型作为人工林研究对象。
1.2 样方选择与建立
参照《IPCC优良做法指南》对系统随机抽样的建议和广西杉木、马尾松人工林的分布特征,基于广西第8次森林资源清查数据和广西用材林龄、林组划分标准(表1),计算2种人工林幼龄林、中龄林、近熟林、成熟林、过熟林5个不同林龄在广西各县(市)的面积、蓄积综合权重,选择权重最大的县(市)为5个不同林龄杉木、马尾松人工林样点,每个样点建立同一林型、同一林龄3块重复样地,各样地的立地条件基本一致,相互距离100 m,样地大小为1000 m2(50 m×20 m),每种人工林15个样地,共计30个样地。将每块样地进一步划分为10个100m2(10 m×10 m)的样方,对样方内胸径(DBH)≥2 cm林木的胸径、树高、冠幅和坐标进行每木调查。样地基本情况见表2。

表1 两种人工林的林龄划分
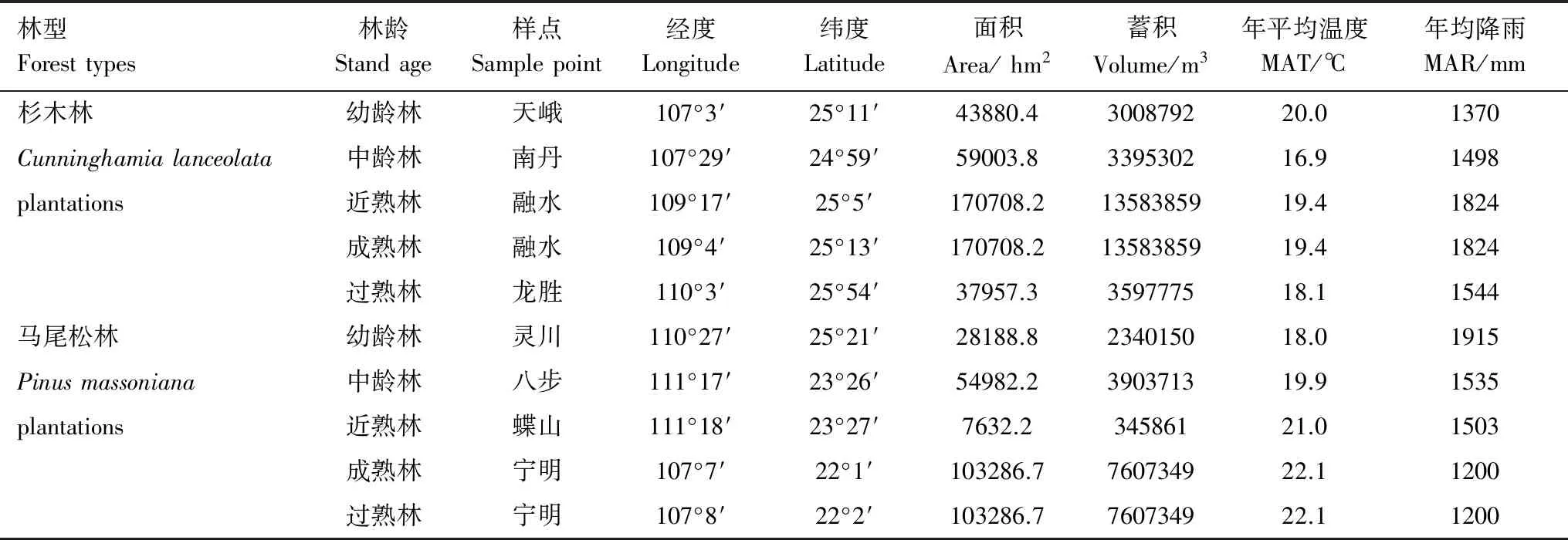
表2 两种人工林不同林龄的样地分布及概况
1.3 根系生物量获取
1.3.1 乔木层
在每木调查的基础上,充分考虑径级分布,遵循中央径级数量多、两端径级数量少的原则,在样地外选择不同林龄和胸径的杉木15株、马尾松21株作为样木,进行生物量的测定。测定其基径、胸径、树高和冠幅并按2 m分段锯断称量,并分别称取枝条、叶片和花果的鲜质量;根的生物量采用全挖法,称鲜重;然后将根混合取样500 g带回实验室在105 ℃烘箱内杀青2 h,调至75 ℃下烘干至恒重,求出根系干鲜质量之比,由此换算出样木根系的干质量,然后建立根系干质量与样木胸径(D)的幂回归方程(W=aDb)、多项式方程(W=a+bD+cD2和W=a+bD+cD2+dD3)和指数方程(W=aebD),择优选择。根据样地每木调查的结果及相应的回归方程计算乔木层各个体的根系生物量,并由此获得乔木层的根系总生物量。
1.3.2 灌木层、草本层
按“品”字型在样地内设置3个2 m×2 m的小样方,调查所有灌木种类、株丛数、高度、基径、覆盖度,在每个2 m×2 m的小样方内各取1个1 m×1 m的小样框,调查草本种类、株丛数、平均高度、覆盖度。采用全挖法收获样框内所有灌木根、草本根,测定其鲜重,烘干换算成干质量,由此计算单位面积灌木、草本根系生物量。
1.3.3 细根(直径≤2 mm根系)
采用土钻法(内径5 cm)在样地10个小样方中心采集0—20,20—40 cm 的土芯10个,分层混合装袋;将样品用流动水浸泡、漂洗、过筛,拣出细根,风干后称鲜重。然后烘干至恒重,保存样品。
1.4 根系全C含量测定
野外所采集植物根样品经粉碎过筛后用于植物全C测定,植物全C采用重铬酸钾氧化-外加热法。
1.5 数据处理
所有数据及多重比较分析、回归方程模型建立均在Excel 2010和SPSS 18.0中进行处理。
2 结果与分析
2.1 乔木根系生物量回归模型建立
大量森林群落生物量的研究显示,乔木各器官及总生物量与测树因子间普遍存在着相关关系,这种相关关系可用数学模型进行拟合。常见的生物量回归模型有三大类,分别是以胸径(D)、树高(H)或胸径平方与树高的乘积(D2H)为自变量的幂函数方程、多项式方程以及指数函数方程。鉴于树高的估测值误差较大,故选择单变量为树木胸径(D)的回归模型作为本研究根系生物量预测方程。同时,通过分析各模型,择出最优模型。本研究中杉木、马尾松根系生物量回归模型分别为WR= 0.01D2.469和WR=0.023D2.241,经统计学检验二者的决定系数分别为0.904和0.984,表明模型拟合效果非常好,能够使用它对两种人工林的根系生物量进行估测。

表3 根系生物量回归模型
2.2 生物量分配
2.2.1 林分各层次根系生物量分配
由表4可知,杉木人工林乔木根系生物量在6.71—30.30 Mg/hm2之间,总体上表现为随林龄增大而增加的趋势,除中龄林至近熟林阶段生物量有所下降且差异不显著外,其他各林龄间差异均达到显著水平。马尾松人工林乔木根系生物量在0.14—51.16 Mg/hm2之间,随林龄增大而增大,除近熟林和成熟林间差异不显著外,其他各林龄阶段均达到显著性水平。除在成熟林和过熟林阶段杉木根系生物量低于马尾松外,其他林龄阶段生物量均高于马尾林。
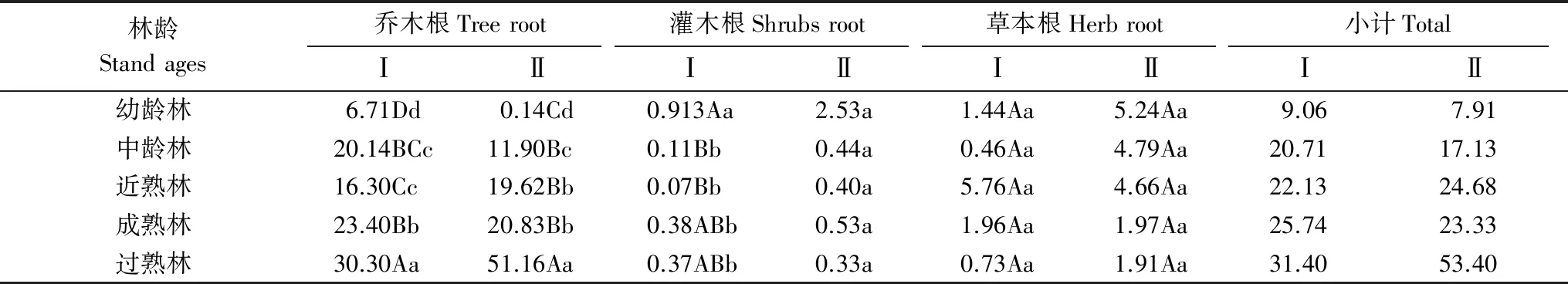
表4 不同林龄阶段各层次根系生物量/(Mg/hm2)
Ⅰ:杉木Cunninghamialanceolataplantations;Ⅱ:马尾松Pinusmassonianaplantations. Similarly here in after
不同林型不同林龄灌木层生物量的大小不同,由于个体小、空间大、阳光充足有利于灌木生长,处于幼龄林时期的杉木和马尾松人工林的灌木根生物量高于其它林龄。其中,杉木人工林的灌木根生物量大小依次为幼龄林成熟林过熟林中龄林近熟林,马尾松为幼龄林成熟林中龄林近熟林过熟林。除杉木幼龄林灌木根系生物量极显著高于杉木林其他林龄阶段外,杉木、马尾松其他林龄阶段差异均未达到显著水平。同一林龄不同林型灌木层生物量大小不同,除过熟林马尾松灌木根系生物量略小于杉木林外,其他林龄阶段灌木根系生物量均为马尾松大于杉木。
杉木林草本层根系生物量随林龄增长的变化规律不明显,近熟林阶段生物量最高,达到5.76 Mg/hm2,马尾松草本层根系生物量则随林龄增大呈逐渐减小的趋势,但两人工林各林龄阶段差异均未达到显著水平。杉木、马尾松两者间除近熟林马尾松草本层根系生物量低于杉木林外,其余阶段均高于杉木林。
杉木、马尾松林地下根系总生物量分别在9.06—31.40,7.91—53.40 Mg/hm2之间,各林龄阶段根系总生物总体上呈现随林龄增加而增加的趋势,但马尾松成熟林阶段(23.33 Mg/hm2)略低于近熟林阶段生物量(24.68 Mg/hm2)。杉木根系总生物量在幼龄林、中林龄和成熟林阶段高于马尾松,在近熟林和过熟林阶段则低于马尾松林。
2.2.2 细根生物量分配
通过对杉木林不同土层深度细根生物量的研究发现,不同林龄杉木林0—20、20—40、0—40 cm土层深度植物细根总生物量的大小顺序总体上为过熟林成熟林近熟林幼龄林中龄林,呈现出先减后增的趋势(表5)。0—40 cm细根生物量在1.04—9.64 Mg/hm2之间,除中龄林0—20 cm土层细根生物量略低于20—40 cm土层外,其余林龄段0—20 cm土层的生物量均高于20—40 cm土层。马尾松人工林不同林龄各土层细根生物量均随着林龄的增加呈现出逐渐减小的趋势,0—40 cm细根生物量在1.44—12.32 Mg/hm2之间,上层土壤(0—20 cm)细根生物量均大于下层(20—40 cm)细根生物量。
2.3 根系碳含量
如表5所示,不同林型不同林龄各层次根系碳含量不同。杉木人工林乔木根、灌木根、草本根、0—20 cm土层细根、20—40 cm土层细根的碳含量分别在583.730—514.770、481.31—544.73、586.81—386.34、575.50— 352.80、569.18—377.51 g/kg之间,其平均含量分别是548.09、503.73、440.35、456.20、434.84 g/kg,表现为乔木根灌木根0—20 cm土层细根草本根20—40 cm土层细根。马尾松人工林乔木根、灌木根、草本根、0—20 cm土层细根、20—40 cm土层细根的碳含量分别在552.82—526.99、528.14—489.61、476.92—441.12、479.86—384.01、468.28—425.94 g/kg,平均含量分别为539.33、501.25、459.66、433.34、443.29 g/kg,表现为乔木根灌木根草本根20—40 cm土层细根0—20 cm土层细根。杉木人工林幼龄林各层次碳含量均高于其他林龄阶段,其中草本根碳含量最高(586.81 g/kg)。

表5 不同林龄阶段细根生物量/(Mg/hm2)
2.4 碳储量分配
2.4.1 林分各层次碳储量分配
由表6可知,杉木、马尾松林各层次根系碳储量及根系总碳储量变化趋势与生物量变化趋势相同。杉木、马尾松乔木层根系碳储量分别在3.92—17.27 Mg/hm2和0.07—28.28 Mg/hm2之间,灌木根系碳储量在0.03—0.50 Mg/hm2和0.17—1.34 Mg/hm2之间,草本根系碳储量在0.28—2.34 Mg/hm2和0.87—2.32 Mg/hm2之间,根系总碳储量在5.26—17.74 Mg/hm2和3.37—29.34 Mg/hm2之间。
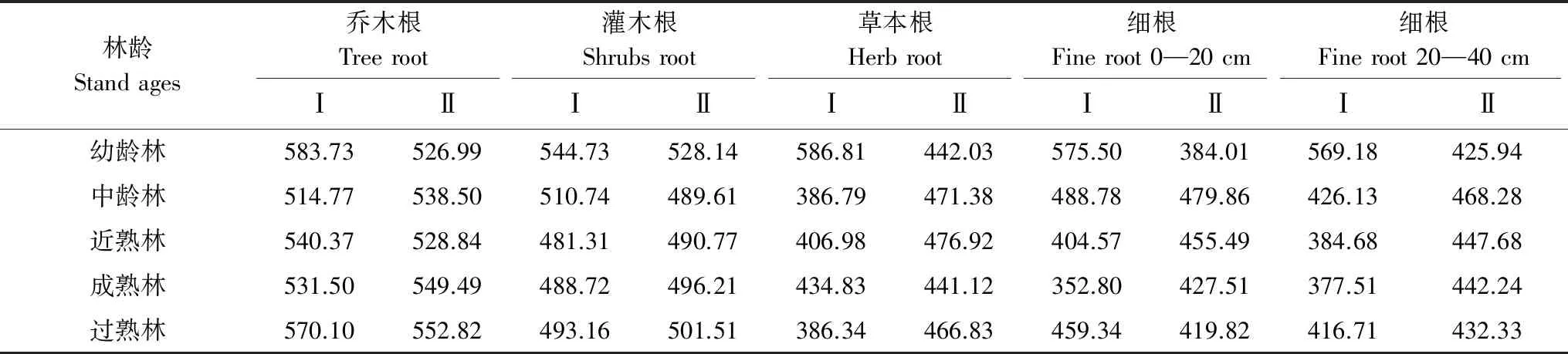
表6 不同林龄阶段各层次根系碳含量/(g/kg)
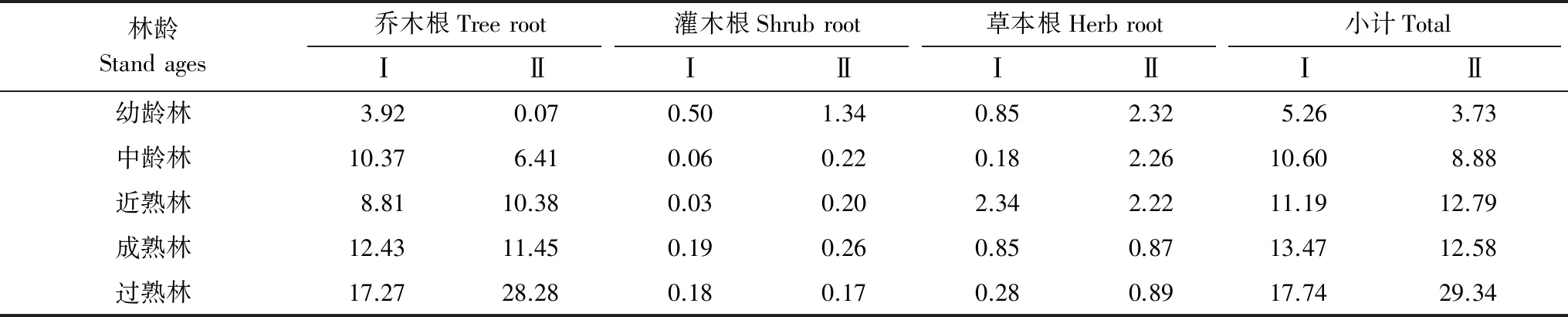
表7 不同林龄阶段各层次根系碳储量/(Mg/hm2)
2.4.2 细根碳储量分配
由表8可知,杉木、马尾松人工林不同土层深度细根碳储量随林龄的变化与生物量变化趋势相似。杉木人工林0—40 cm土层细根总碳储量在0.47—4.23 Mg/hm2之间,0—20 cm和20—40 cm土层深度细根碳储量分别为0.23—2.31,0.24—1.93 Mg/hm2,除中龄林0—20 cm土层细根碳储量略低于20—40 cm土层外,其余林龄阶段碳储量均为0—20 cm土层大于20—40 cm土层。不同林龄马尾松人工林0—40 cm土层细根总碳储量在0.61—4.95 Mg/hm2之间,其0—20 cm土层与20—40 cm土层细根碳储量分别为0.51—2.69,0.09—2.26 Mg/hm2,均为表现为0—20 cm土层大于20—40 cm土层。
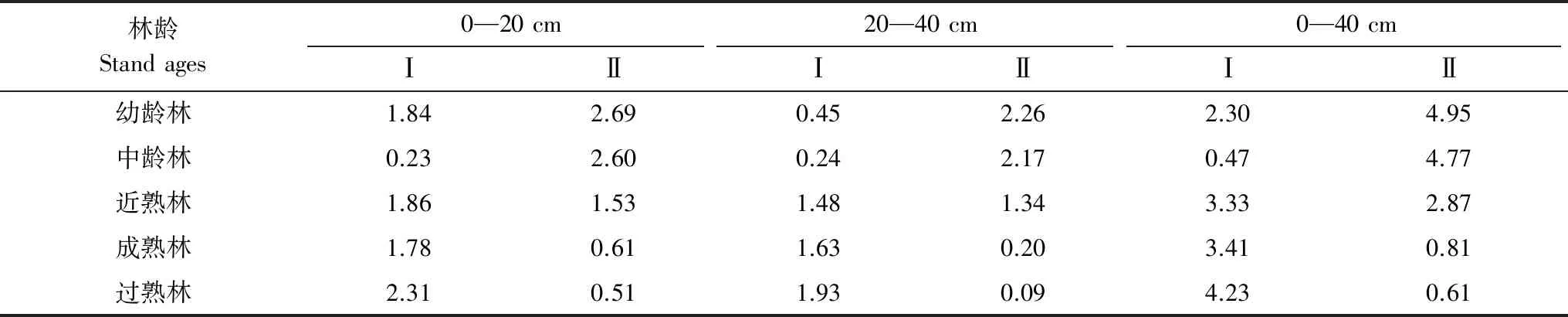
表8 不同林龄阶段细根碳储量/(Mg/hm2)
2.4.3 根系碳储量分配比例
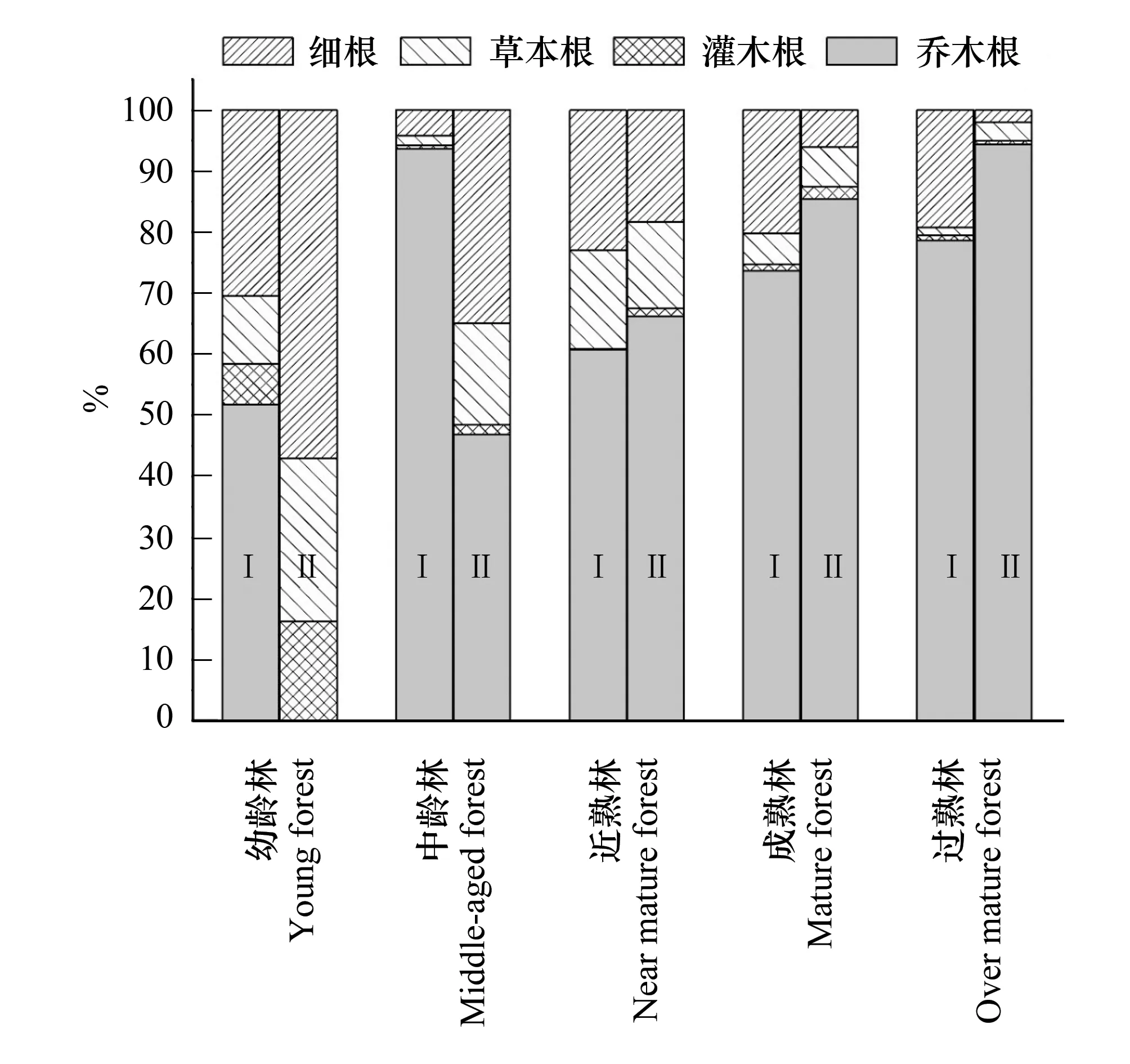
图1 不同林龄阶段各层次碳储量分配比例 Fig.1 Distribution ratio of carbon storage at various levels in different stages of forest age
由表6和表8可知,杉木林不同林龄阶段各层次根系和土壤细根总碳储量分别为7.56、11.07、14.52、16.88、21.97 Mg/hm2,随林龄增大呈逐渐增加的趋势。马尾松各林龄阶段地下根系总碳储量分别为8.68、13.65、15.66、13.39、29.95 Mg/hm2,除成熟林阶段比近熟林阶段有所下降外,总体上呈现逐步增加的趋势。马尾松成熟林阶段地下根系总碳储量小于杉木林,其余各阶段均大于杉木林地下总碳储量。
由图1可知,杉木人工林乔木根系碳储量占地下总碳储量的绝对优势,其比例为51.82%—78.63%,均超过50%,除中林龄所占比例达93.64%外,其余各阶段基本随林龄增加而增加;灌木根所占比例为0.23%—6.58%,草本根所占比例为1.28%—16.15%,细根所占比例为4.25%—30.43%;各林龄阶段各层次所占比例均表现为乔木根细根草本根灌木根。除马尾松人工林除幼龄林乔木根所占比例为0.85%外,其他阶段在46.94%—94.43%之间,随林龄的增大而增大,是地下根系碳储量的主要储量位置,灌木根、草本根和细根所占比例分别为0.55%—15.40%、2.98%—26.70%和2.04%—57.05%,总体上各层次所占比例表现为随林龄增大而逐渐减小的趋势。马尾松幼龄林阶段各层次碳储量所占比例表现为细根草本根灌木根乔木根,过熟林阶段表现为乔木根草本根细根灌木根,其他林龄阶段为乔木根细根草本根灌木根。
3 讨论
森林生物量的获取可通过直接与间接两种途径:前者因破坏性大,很少使用;后者是确定生物量的主要方法,也叫生物量模型法。目前常用的生物量拟合模型变量有胸径(D)、树高(H)、密度(S),常见的预测模型包括以胸径为单变量的模型、以胸径和树高为变量的模型以及以胸径、高度、密度为变量的模型[17,19]。由于树高在大多数调查中存在着较大误差,因此在森林生物量的预测中以胸径为单变量的模型被广泛运用。本研究得出以胸径为单变量的幂函数模型具有较高精度,能够对杉木、马尾松两种人工林根系生物量进行有效预测。
森林生物量与许多生物学因素和非生物学因素密切相关,如区域的水热条件、土壤条件以及森林的类型、年龄、优势种的组成,活立木密度等[20-21]。人工林乔木根生物量往往随林龄的增长而迅速增加,徐冰等[20]对中国36类主要森林类型研究表明,大多数森林类型生物量密度与林龄呈现Logistic曲线关系,即随林龄的增加生物量、碳密度呈上升趋势。然而,由于受林分抚育措施等的影响,其增长趋势也会发生变化。本研究中杉木、马尾松乔木根系生物量与林分根系总生物量(乔木根、灌木根、草本根)呈现出随林龄增大而增加的趋势,而在林分抚育等管理措施下,杉木人工林乔木根系中林龄至近熟林阶段生物量有所下降,在自然和人为因素的影响各林龄阶段增长幅度有所不同,同邓坤枚等[22]得出云南松林根系总生物量随林分年龄呈递增趋势的结论一致。
一般而言,随土壤层次的加深,林木根系会大幅减少[23-24]。本研究中,随土层深度的增加,土壤质地愈发紧实,温度愈低,养分含量减少,两种人工林的细根生物量都呈现出逐渐降低的趋势,这与云南松林根系生物量主要集中分布在0—30 cm的土层中[22]的研究结果及长白山地区3种森林类型[27]和密歇根北部阔叶林[28]的根系生物量随土层深度分布的规律一致。细根生物量随土层的加深而减少,且多集中分布在土壤表层,主要受土壤理化性质和温度、水分含量的影响[29]。土壤各层次细根分布的差异可反映出细根功能转换及土壤资源有效性的变化情况。由于表层土壤养分含量高,水分充足,温度适宜,因而细根主要集中在表层内以获取水分和养分;而分布在下层的细根主要功能为吸收水分[28]。本研究中,除中龄林0—20 cm土层细根生物量略低于20—40 cm土层外,其余林龄段0—20 cm土层的生物量均高于20—40 cm土层,符合随土层加深而减少的规律性变化,可见生物量主要集中分布在表层土壤中。
本研究中,杉木、马尾松林各层次根系碳储量、根系总碳储量及各土层深度细根碳储量随林龄的变化趋势与生物量变化趋势相似。杉木和马尾松人工林不同土层深度细根碳储量大体上表现为随土层的加深而减少,与陶玉华等[16]及黄林等[29]的研究结果一致。在根系碳储量分配比例上,除马尾松幼龄林乔木根系碳储量所占比例很少外,杉木和马尾松其他林龄乔木层根系碳储量比例均占绝对优势,呈现出随林龄增大比例越大的趋势,而灌木和草本根系碳储量很少,土壤细根也是森林生态系统地下碳储量的重要组分。总之,地下根系碳储量是森林生态系统碳储量估算中不可忽视的一部分,对地下过程的研究有助于更准确地估算森林生态系统总碳库存。
最后,本研究采用全根挖掘法和土钻法获取根系生物量时会产生某些误差,尤其在进行细根取样时,难以避免地会发生损失。但相较于其他耗时久、工作量大却依旧无法保证没有损耗的方法而言,只要在取根时足够仔细,使用全根挖掘法和土钻法一样能得到预期结果。
[1] Canadell J, Jackson R B, Ehleringer J B, Mooney H A, Sala O E, Schulze E D. Maximum rooting depth of vegetation types at the global scale. Oecologia, 1996, 108(4): 583- 595.
[2] Dannowski M, Block A. Fractal geometry and root system structures of heterogeneous plant communities. Plant and soil, 2005, 272(1/2): 61- 76.
[3] 王政权, 郭大立. 根系生态学. 植物生态学报, 2008, 32(6): 1213- 1216.
[4] 贺金生, 王政权, 方精云. 全球变化下的地下生态学: 问题与展望. 科学通报, 2004, 49(13): 1226- 1233.
[5] 李子敬, 陈晓, 舒健骅, 孙红彦, 丛日晨. 树木根系分布与结构研究方法综述. 世界林业研究, 2015, 28(3): 13- 18.
[6] 陈红, 冯云, 周建梅, 徐振国, 廉超, 郭起荣. 植物根系生物学研究进展. 世界林业研究, 2013, 26(5): 25- 29.
[7] 孙兆国. 镧和酸雨对大豆幼苗根系生长和氮素营养的复合影响研究[D]. 无锡: 江南大学, 2014.
[8] 黄建辉, 韩兴国, 陈灵芝. 森林生态系统根系生物量研究进展. 生态学报, 1999, 19(2): 270- 277.
[9] 刘其霞, 常杰, 江波, 袁位高, 戚连忠, 朱锦茹, 葛滢, 沈琪. 浙江省常绿阔叶生态公益林生物量. 生态学报, 2005, 25(9): 2139- 2144.
[10] 杨丽韫, 罗天祥, 吴松涛. 长白山原始阔叶红松林不同演替阶段地下生物量与碳、氮贮量的比较. 应用生态学报, 2005, 16(7): 1195- 1199.
[11] 冯宗炜, 陈楚莹, 张家武, 王开平, 赵吉录, 高虹. 湖南会同地区马尾松林生物量的测定. 林业科学, 1982, 18(2): 127- 134.
[12] 田大伦. 杉木林生态系统学. 北京: 科学出版社, 2003: 15- 16.
[13] 周政贤. 中国马尾松. 北京: 中国林业出版社, 2001: 53- 60.
[14] 林生明, 徐土根, 周国模. 杉木人工林生物量的研究. 浙江林学院学报, 1991, 8(3): 21- 27.
[15] 张治军, 王彦辉, 袁玉欣, 李志勇, 曹磊, 张国增, 于澎涛, 王颖. 马尾松天然次生林生物量的结构与分布. 河北农业大学学报, 2006, 29(5): 37- 43.
[16] 陶玉华, 冯金朝, 马麟英, 隆卫革, 曹书阁. 广西罗城马尾松、杉木、桉树人工林碳储量及其动态变化. 生态环境学报, 2011, 20(11): 1608- 1613.
[17] 王维枫, 雷渊才, 王雪峰, 赵浩彦. 森林生物量模型综述. 西北林学院学报, 2008, 23(2): 58- 63.
[18] 黄从德, 张健, 杨万勤, 唐宵, 赵安玖. 四川省及重庆地区森林植被碳储量动态. 生态学报, 2008, 28(3): 966- 975.
[19] 李燕, 张建国, 段爱国, 相聪伟. 杉木人工林生物量估算模型的选择. 应用生态学报, 2010, 21(12): 3036- 3046.
[20] 徐冰, 郭兆迪, 朴世龙, 方精云. 2000—2050年中国森林生物量碳库: 基于生物量密度与林龄关系的预测. 中国科学: 生命科学, 2010, 40(7): 587- 594.
[21] 侯振宏, 张小全, 徐德应, 于澎涛. 杉木人工林生物量和生产力研究. 中国农学通报, 2009, 25(5): 97- 103.
[22] 邓坤枚, 罗天祥, 张林, 王学云, 李长会. 云南松林的根系生物量及其分布规律的研究. 应用生态学报, 2005, 16(1): 21- 24.
[23] Leuschner C, Hertel D, Schmid I, Koch O, Muhs A, Hölscher D. Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility. Plant and Soil, 2004, 258(1): 43- 56.
[24] Schenk H J, Jackson R B. The global biogeography of roots. Ecological Monographs, 2002, 72(3): 311- 328.
[25] 张艳杰, 温佐吾. 不同造林密度马尾松人工林的根系生物量. 林业科学, 2011, 47(3): 75- 81.
[26] 高祥, 丁贵杰, 翟帅帅, 陈模芳, 杜华东. 不同林分密度马尾松人工林根系生物量及空间分布研究. 中南林业科技大学学报, 2014, 34(6): 71- 75.
[27] 杜晓军, 刘常富, 金罡, 石小宁. 长白山主要森林生态系统根系生物量研究. 沈阳农业大学学报, 1998, 29(3): 229- 232.
[28] Burton A J, Pregitzer K S, Hendrick R L. Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests. Oecologia, 2000, 125(3): 389- 399.
[29] 黄林, 周立江, 王峰, 黄茹, 田锋, 齐代华, 唐元会. 红壤丘陵区典型植被群落根系生物量及碳储量研究. 水土保持学报, 2009, 23(6): 134- 138.
Biomass and carbon storage in roots ofCunninghamialanceolataandPinusmassonianaplantations at different stand ages in Guangxi
HAN Chang1,3,4, SONG Min2,3,4, DU Hu3,4, ZENG Fuping3,4, PENG Wanxia3,4, WANG Hua1,*, CHEN Li1,3,4, SU Liang1,3,4
CollegeofBioscienceandBiotechnology,HunanAgriculturalUniversity,Changsha410128,China2CollegeofAgriculture,HunanAgriculturalUniversity,Changsha410128,China3KeyLaboratoryofAgro-ecologicalProcessesinSubtropicalRegions,InstituteofSubtropicalAgriculture,ChineseAcademyofSciences,Changsha410125,China4HuanjiangObservationandResearchStationofKarstEcosystem,ChineseAcademyofSciences,Huanjiang547100,China
CunninghamialanceolataandPinusmassonianaplantations cover a large area in Guangxi. In the present study, dynamics of biomass and carbon storage in roots of the two types of plantations were investigated. The plantations were selected according to five stand ages, i.e., young forest, middle-aged forest, near mature forest, mature forest and over mature forest. Two methods, i.e., whole soil excavation and soil auger, were adopted for root sampling. Roots were divided according to trees, shrubs, or grass. The results showed that total root biomass inC.lanceolataandP.massonianaplantations ranged from 9.06 to 31.40 and 7.91 to 53.40 Mg/hm2, respectively. Total root biomass increased with stand age. The fine root biomass inC.lanceolataplantations decreased initially but then increased with stand age, but that inP.massonianaplantations decreased with stand age. Root carbon content in different soil horizons changed in the order of tree shrub herb roots. Carbon storage in roots of both types of plantations changed similarly to root biomass. The carbon storage of roots in different soil horizons ranged from 7.56 to 21.97 and 8.86 to 29.95 Mg/hm2, respectively, forC.lanceolataandP.massonianaplantations. Overall, the carbon storage of roots was dominated by tree roots that increased with forest age.
carbon storage; roots; stand age;Cunninghamialanceolata;Pinusmassoniana
10.5846/stxb201511292394
中国科学院科技服务网络计划STS项目(KFJ-EW-STS-092); 国家自然科学基金项目(31370485, 31370623, 31400412,31460135); 广西科技惠民计划项目(桂科转1599001- 6);广西特聘专家资助项目
2015- 11- 29; 网络出版日期:2016- 08- 30
韩畅,宋敏,杜虎,曾馥平,彭晚霞,王华,陈莉,苏樑.广西不同林龄杉木、马尾松人工林根系生物量及碳储量特征.生态学报,2017,37(7):2282- 2289.
Han C, Song M, Du H, Zeng F P, Peng W X, Wang H, Chen L, Su L.Biomass and carbon storage in roots ofCunninghamialanceolataandPinusmassonianaplantations at different stand ages in Guangxi.Acta Ecologica Sinica,2017,37(7):2282- 2289.
*通讯作者Corresponding author.E-mail: wangchina926@163.com
猜你喜欢
防护林科技(2022年5期)2022-08-22
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
水土保持研究(2020年2期)2020-03-19
绿色科技(2019年5期)2019-11-29
乡村科技(2019年21期)2019-02-21
生态学报(2018年17期)2018-10-18
四川农业大学学报(2018年3期)2018-09-04
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
