
深远海浮游动物生态学研究进展
2017-06-22 13:47王春生
生态学报 2017年10期
孙 栋,王春生,2,*
1 国家海洋局第二海洋研究所, 海洋生态系统与生物地球化学海洋局重点实验室, 杭州 310012 2 国家海洋局第二海洋研究所, 卫星海洋环境动力学国家重点实验室, 杭州 310012
深远海浮游动物生态学研究进展
孙 栋1,王春生1,2,*
1 国家海洋局第二海洋研究所, 海洋生态系统与生物地球化学海洋局重点实验室, 杭州 310012 2 国家海洋局第二海洋研究所, 卫星海洋环境动力学国家重点实验室, 杭州 310012
深远海浮游生态系统依据水深的差别可以被划分为几个子系统,包括上层、中层、深层和深渊层等。目前大量的研究结果表明在海洋上层,由于光照、温度、海冰和营养盐补充等因素的影响,浮游生物群落往往呈现出显著的时空变化。但在海洋中层及更深的区域,传统观点认为随着水深的增加,海洋的理化环境趋向于稳定,在这一区域生态系统的时空变化要弱于海洋上层。同时受调查技术和经费的限制,人们对于这一广阔区域内浮游生态系统时空变化规律的认识要局限的多。随着我国海洋科学的发展和海洋强国战略的实施,海洋科学研究也逐渐由过去的以近海研究为主发展到当前的近海、边缘海和深远海研究协同发展。但与我国近海浮游生态学过去数十年间积累的大量研究成果相比,我国科学家对深远海,特别是中层、深层和深渊层浮游生态学方面的研究极为缺乏。从大洋浮游动物群落的垂直分布及其变化、种间关系与生态位分化、深海浮游动物群落在碳沉降和生物地球化学循环中的作用等多个角度全面总结了当前国内外深海浮游生态学的研究进展,同时介绍过去十余年来深海浮游生态学研究技术手段上的巨大进步,以期为今后国内同行的研究提供参考和借鉴。
浮游生态系统;垂直分布;缺氧;季风;海山;生物地球化学循环
海洋占据了地球表面积的71%,同时也是大量微生物、植物和动物物种的栖息地,它们与海洋环境一起构成了地球上最大的生态系统。依据生物群落的特征与理化环境的显著差异,特别是水深的差别,海洋生态系统可以被划分为浅海生态系统和深远海(大洋)生态系统。前者主要是指位于陆架上的海底及其上方水层中的生态系统,而后者指陆架边缘以外的海洋生态系统,包括位于陆坡、洋盆、洋脊、海沟等海洋地貌上的海底及其上方水层中的生态系统。对于大洋生态系统而言,通常可以依据水深和光照情况的不同划分为数个子系统,包括200 m以浅、光照条件可以维持浮游植物和光合微生物进行净初级生产的海洋上层(epipelagic zone);200—1000 m、光照大大减弱以至于不足以维持浮游植物及光合微生物进行净初级生产的海洋中层(mesopelagic zone);1000 m以下、光照完全消失以至于完全依赖上层海洋生产力输出的广义海洋深层(deep sea)。而对于广义的海洋深层又可以依据环境的差异和生态系统类型的不同而进一步划分为1000—4000 m的狭义海洋深层(bathypelagic zone)、4000—6000 m的深渊层(abyssopelagic zone)和大于6000 m的超深渊层(hadalpelagic zone)(图1)[1-2]。为了表达上的简便,如不特别指明,下文将海洋中层、深层和深渊层等统称为深海。

图1 海洋生态系统的垂直划分(仿[2],修改) Fig.1 Vertical zonation of marine ecosystem (Modified from[2])
海洋浮游生态学是海洋科学的一个重要分支,主要研究内容是浮游生物种群、群落和生态系统等各个层次的时空变换规律及其内在机制。由于采样调查上的便利和与渔业活动的直接关联性,自20世纪初开始人们就对海洋上层的浮游生态系统展开了系统的调查研究[3-4]。目前大量的研究结果表明在海洋上层,由于光照、温度、海冰和营养盐补充等因素的影响,浮游生物群落往往呈现出显著的时空变化[5-6]。但在海洋中层及更深的区域,传统观点认为随着水深的增加,海洋的理化环境趋向于稳定,在这一区域生态系统的时空变化要弱于海洋上层。同时受调查技术和经费的限制,人们对于这一广阔区域内浮游生态系统时空变化规律的认识要局限的多。早期,人们对这一深度范围的浮游动物群落的初步认识和了解主要来源于几次大规模的区域性海洋综合调查(如国际印度洋调查,IIOE),随着过去20年以来全人类对于全球变化问题的重视,人们逐渐认识到海洋中层及更深层区域的浮游生态系统对于生物地球化学循环、海气相互作用和全球变化的重要意义[7-8],并且随着研究技术手段的进步,关于海洋中层及更深层区域浮游生态学的相关研究报道正在快速增加(图 2)。

图2 关于过去25年间深海浮游动物生态学研究的文献计量学分析Fig.2 Bibliometrics analysis of deep-sea zooplankton ecology research over the past 25 years基于Web of Science数据库;海洋中层浮游动物研究,检索逻辑:“zooplankton” and “twilight zone” or “mesopelagic”,检索结果860篇;海洋深层浮游动物研究,检索逻辑:“zooplankton” and “dark zone” or “bathypelagic”,检索结果304篇)
随着我国海洋科学的发展、综合国力的提升和海洋强国战略的实施,海洋科学研究也逐渐由过去的以近海研究为主发展到当前的近海、边缘海和深远海研究协同发展[9]。但是与我国近海浮游生态学过去数十年间积累的大量研究成果相比,我国科学家对深远海,特别是中层、深层和深渊层浮游生态学方面的研究极为缺乏。本文试图从深海浮游动物群落的垂直分布及其变化、种间关系与生态位分化、深海浮游动物群落在生物地球化学循环中的意义等多个角度总结当前国内外对于深海浮游动物生态学的研究进展,并介绍过去十余年来深海浮游生态学研究技术手段上的巨大进步,以期为今后国内同行的研究提供参考和借鉴。
1 深海浮游动物群落的垂直分布及其变化
在海洋上层,随季节变化的光照和温度条件以及由此驱动的海水物理、化学环境的协同变化导致温带和极地海洋初级生产力会呈现出显著的季节特征[10-11],另外在一些热带和亚热带大洋及边缘海,季风引发的上升流强度和营养盐水平的变化也会导致显著的初级生产力季节变化[12],这些变化都通过上行控制效应而进一步导致海洋上层浮游动物的丰度、生物量和群落结构等表现出明显的季节动态[13-14]。而在深海,传统的观点认为这一区域的基本环境特征是温度低,盐度和溶解氧浓度非常稳定,因此在这种环境下深海生物群落的季节变化会很微弱[15]。但是过去大约30年间的研究结果大大改变了这一旧有的观念,人们逐渐认识到深海动物群落几乎完全依赖于海洋上层产生的物质和能量输出[16],因此海洋上层的季节变化必然会通过一系列途径影响到深海动物群落。下面,本文从全球尺度上对热带和亚热带海洋、温带海洋、极地和亚极地海洋等三个差异显著的浮游生态系统分别简要概述,并且对国际学术界特别关注的热带大洋中层缺氧现象、季风及海山等海底地貌结构对大洋浮游生态系统的影响等三方面问题进一步详细论述。
1.1 热带、亚热带海洋
在典型的热带、亚热带大洋,由于强烈的层化作用导致海洋上层缺乏营养盐补充,因此海洋上层的初级生产力很低,同时初级生产力和叶绿素浓度的时空变化不明显[17]。在这种情况下,热带、亚热带海洋的浮游动物群落不仅生物量很低,而且生物量呈现随着水深增加而快速递减的趋势。例如,Steinberg等[18]对中太平洋ALOHA海洋监测站(23°N)0—1000 m浮游动物的垂直分布的研究表明夜间0—50 m浮游动物生物量为7.1—9.5 mg/m3,在300—500 m则快速降低至不到1.0 mg/m3,到500—750 m时更是小于0.5 mg/m3。在极度寡营养的热带西太平洋“采薇”海山附近(15°N)的研究也发现了类似的规律:0—100m水层和100—200m水层的浮游动物丰度分别为72.11 个/m3和36.41 个/m3;而海洋中层的浮游动物丰度很低,各水层的丰度均小于3.00 个/m3。总体来说,在季节变化方面,由于热带、亚热带海洋的表层温度常年较高,水体结构稳定,海洋上层初级生产力状况由于常年受到营养盐的限制而季节变化很小(季风区除外),因此这一区域深海浮游动物群落的季节变化不显著。
1.2 温带海洋
在温带海洋,海洋上层的温度会呈现出较为明显的季节变化[2]。在冬季,不稳定的水体结构有利于深层营养盐的垂直补充,随着春季光照的增加和水温的上升,浮游植物能够利用水体中充沛的营养盐而出现旺发现象[19],这导致温带海洋上层的浮游动物生物量在浮游植物旺发季节往往显著高于其他季节[20],海洋上层较高的生产力会随着浮游生物垂直迁移、粪便和生物残屑的沉降等方式进一步影响深海浮游生物群落[21-22]。因此,与热带、亚热带海洋深海浮游动物群落缺乏季节变化相比,温带海洋深海浮游动物群落往往呈现出较强的季节变化。例如,在东北大西洋(47°N),200—400 m水层的浮游动物生物量春季要显著高于夏季,但在更深的位置(400—1000 m),生物量的季节差别就不再显著,这很好地反映了海洋上层生产力的变化;此外,由于夏季在海洋深层(1000—2000 m)有较大规模的哲水蚤休眠种群存在,因此这一深度的浮游动物生物量呈现出夏季高于春季的特征[23]。另外,Siokou等[20]对春季和秋季爱琴海(36—41°N)0—1000 m浮游动物群落的研究也表明海洋上层和海洋中层的浮游动物生物量和群落结构均存在着明显季节差异,同时在中尺度上,浮游动物群落在上层和中层均表现出较明显空间差异。
1.3 极地、亚极地海洋
对于极地、亚极地海洋生态系统来说,夏季冰层的融化是一个决定性的事件[24],随着冰层融化、温度上升,浮游植物进入旺发状态[11],充足的食物支持了这个季节的海洋上层具有极高的浮游动物生物量,而随着上层营养盐的耗尽,浮游植物生物量下降,积累了充足脂类物质的浮游动物沉降至中层乃至更深的低温水域进入季节性休眠状态[25-26]。在冰岛附近北大西洋(66—71°N)开展的浮游动物群落研究证实了上述理论:冬季海洋中层各个采样深度范围内的生物量都显著高于春季和夏季,而同时海洋上层的生物量则显著低于春季和夏季;进一步的研究发现哲水蚤属和长腹水蚤属的季节性垂直迁移是造成这一现象的主要原因[27]。此外,很多浮游动物类群有昼夜垂直移动习性,这往往造成中低纬度海区浮游动物生物量和群落结构的昼夜垂直分布存在明显差异[18, 28],但在高纬度海区的浮游动物需要充分利用短暂的浮游植物旺发季节积累脂类物质以保证繁殖或休眠活动的需要,因此往往不表现出昼夜垂直迁移现象[26, 29]。另外,由于极地海区水温较低,浮游动物通常需要经历数个生长季的生长才能够达到性成熟,因此在分析高纬度海区浮游动物,特别是桡足类的生态学现象时,优势种群的年龄结构及其变化往往不可忽视[30]。
1.4 热带大洋中层缺氧现象的影响
现代海洋学研究发现在热带东太平洋、北印度洋和热带东大西洋等海区的中层均存在着大面积的缺氧层,这种真光层以下的缺氧现象主要是由于海洋上层沉降下来的有机质在热带海区相对温暖的中层被大量微生物分解利用,好氧微生物的活动消耗了水体中的溶解氧,但稳定的水体结构导致中层水体缺乏垂直方向上的交换,从而无法获得上层和深层富氧水的补充,最终导致在海洋中层出现一个溶解氧水平显著低于海洋上层一个数量级甚至更低的缺氧层(oxygen minimum zone, OMZ)[31- 33]。海水中充沛的溶解氧通常是大多数海洋动物的生存都必须依赖的重要环境条件[34]。对于浮游动物来说,极低的溶解氧环境通常会对其垂直分布特征产生显著的限制作用。例如,Wishner等[35]发现阿拉伯海中部水域(16°N)的中层(250—500 m)可以出现溶解氧低于0.2 mL/L的OMZ区域,同时浮游动物生物量的垂直分布呈现出0—200 m水层最高,500—1000 m水层其次,250—500 m水层最低的双峰模式。与类似纬度范围、但不存在中层缺氧现象的西太平洋和中太平洋的研究结果相比,这一结果很好地体现了中层缺氧现象对深海浮游动物垂直分布的强烈限制作用。另外,一些浮游动物种类在长期的进化过程中具备了对低氧环境的特殊适应能力,例如研究发现一种阿拉伯海中层分布的光水蚤Lucicutiagrandis集中分布于600—1000 m的深度,而同步测定的这一水层溶解氧仅为0.07—0.15 mL/L 的水平;一些乳点水蚤属Pleuromamma、长腹水蚤属Metridia和真哲水蚤属Eucalanus的种类也被证明对海洋中层的缺氧环境有着极佳的适应能力[36]。因此,由于不同种类的浮游动物对于OMZ表现出的耐受差异,海洋中层的缺氧现象不仅仅可以影响浮游动物生物量和丰度的垂直分布,而且还可以显著影响群落结构的垂直分布模式和昼夜垂直移动(diel vertical migration, DVM)模式如下[37-38]:
(1) 上层-中层双峰分布型:种群在OMZ以浅的真光层存在一个丰度上的峰值,在OMZ水层丰度很低,在OMZ以深的水层存在一个次级峰值,表明自然状态下这是一个上层-中层广深度分布的种类,因躲避OMZ而出现双峰分布。
(2) 跨OMZ-DVM型:夜间种群垂直迁移至OMZ以浅的真光层中摄食,白天垂直沉降至OMZ以深的富氧低温水层(种群在DVM过程中经过OMZ的时间很短暂)。
(3) 上层单峰分布型:种群在OMZ以浅的真光层存在一个丰度上的峰值,在OMZ水层和OMZ以深丰度均很低,即OMZ以下不存在丰度的次级峰值(这表明自然状态下这是一个海洋上层分布的种类,因此不受OMZ限制)。
(4) 次表层-中层双峰分布型:种群在混合层中的分布并不均匀,表层丰度很低,在次表层存在峰值,在OMZ水层丰度很低,在OMZ以深又存在一个次级峰值(这表明自然状态下种群应为次表层-中层分布,因躲避OMZ而出现双峰分布)。
1.5 季风的影响
在远离大陆的热带、亚热带海区,通常情况下由于稳定的海流和温度条件导致海洋上层生态系统的季节变化很微弱(如热带中太平洋)。但是在一些受到季风影响的热带、亚热带海区,季风和环流的季节变化及上升流、温盐状况的变化会对海洋上层生态系统产生深刻影响[39-40]。例如在阿拉伯海,夏季强烈的西南季风会导致索马里近海出现很强的离岸上升流和深入阿拉伯海离岸区的芬勒特射流[41],充沛的营养盐导致阿拉伯海上层浮游动物生物量急剧提高[42]。在海洋中层,时间序列沉积物捕获器的数据清晰地表明海洋上层初级生产力的大幅提高导致中层POC通量在季风季节的显著升高[43],同时这一水层浮游动物生物量和群落结构也出现了相应的变化[44-45]。在海洋深层,尽管季风导致的POC通量季节变化依然清新存在,但浮游动物生物量的季节变化几乎完全消失(15—16°N)[44]。这些研究表明,深海浮游动物群落受海洋上层季风影响的变化幅度会随着深度的增加而逐渐减小。
1.6 海山的影响
按照一般的定义,海山是指高出海底达到1000 m及以上,且不露出海面的海底凸起物[46]。基于不同海山顶部距离海平面的高度差异和它们在水文及生态特征上的差别,可以将海山分为浅海海山(海山顶部进入真光层)、中深度海山(海山顶部比真光层深,但小于400m)和深海海山(海山顶部比400m深)[47]。经过约半个世纪以来对太平洋和大西洋各类海山区的生态研究和渔业调查,人们发现海山区的水体和海底均拥有明显不同于大洋其他区域的独特生态系统,其典型特征就是海山区的浮游生物、游泳生物和底栖生物在生物量、丰度、多样性等方面都显著不同于周边的大洋生态系统和海底生态系统,这一系列现象被归纳为“海山效应”(Seamount effects)[48-49]。针对浅海海山的浮游生态学调查较多,很多这类调查得到海山顶部及附近真光层中的初级生产力、浮游植物和浮游动物生物量均显著高于周边大洋水域的结论[50-51],而针对深海海山及其对中层、深层浮游动物群落影响的研究却非常缺乏。一个涵盖不同深度海山浮游生态系统的例子是Martin和Christiansen[52]对于东北大西洋三座海山的调查,他们发现两座中深度海山顶部0—200 m层浮游动物生物量要低于周边海域,同时海域中层浮游动物生物量差别不大;但对同区域一座深海海山的调查发现海山山坡附近的中层水体浮游动物生物量要显著低于周边海域。又如,在东太平洋的一个深海海山区,Saltzman和Wishner[37]也发现桡足类的生物量和丰度在水深约600—800 m处具有一个明显的次级峰值,这一深度与海山顶部的水深非常接近(960 m),而通常情况下在这一深度桡足类的丰度会大大低于上层水体。这些研究表明不同深度的海山对浮游动物群落的影响可以显著不同,同时过去的研究很多时候也忽视了海山对于中层和深层浮游动物群落的影响,即使在关于浅海海山的研究中亦是如此。
与此同时,人们也从物理海洋学、生态动力学、地质学、动物行为学等不同角度提出了一系列假说来试图解释这些现象,比较有代表性的包括“上升流-初级生产提高”假说[53]、“垂直迁移受阻-空窗”假说[54]、“能量效率-外来补充”假说[55]和“逆上升流/下降流”假说[56]等如下。
(1) “上升流-初级生产提高”假说:此假说认为海山区复杂动力过程产生的上升流和涡旋将导致营养盐丰富的深层海水进入真光层,这些营养盐促进了各类浮游植物的生长从而极大提高了海山区真光层的初级生产力,这种促进作用在寡营养的大洋区域会尤其明显,这进一步支持了浮游动物和食物链上更高营养级生物量的增加。
(2) “垂直迁移受阻-空窗”假说:此假说认为浅海海山和中深度海山将对浮游动物的昼夜垂直迁移产生强烈的阻碍作用,因此白天在海山上方水体中的植食性浮游动物将因为无法下沉进入安全深度而遭到捕食,从而可以在海山上方的水体中观察到一个低丰度和低生物量的植食性浮游动物空窗。
(3) “能量效率-外来补充”假说:此假说认为很多海山区单纯依赖“上升流-初级生产提高”机制增加的生产力并不足以支持各类浮游动物和游泳动物的代谢活动,因此必然存在某些周边大洋区域物质和能量补充海山区生态系统的途径。
(4) “逆上升流/下降流”假说:某些浮游动物对海山区存在的上升流或下降流做出行为学上的响应,通过主动的逆流运动而保持在特定深度。
但当前的主要问题是大量的野外证据无法完全证实其中任何一种假说,不但对于同一海山生态系统不同年份的调查中会得到完全不同的结论,甚至针对同一海山在同一航次调查的不同时间段获得的数据都有显著差别[50- 52, 57-58]。针对这些不一致的野外证据,Genin[47]建议只有在较浅的海山,当存在较长时间和较为强烈的上升流,使得富营养的深海海水能够上升进入真光层并且在真光层中停留足够长的时间以满足浮游植物种群增殖时,经典的“上升流-初级生产提高”假说才可以用于解释海山区存在较高浮游动物生物量的现象。此外,上述的大部分假说(除“能量效率-外来补充”假说外)都无法从理论上解释在深海海山区发现的中层和深层浮游动物聚集现象。因此,关于海山,特别是深海海山对于中层和深层浮游动物群落的影响还需要深入研究。
2 深海浮游动物群落的种间关系和生态位分化
2.1 在群落尺度上
长期以来,生物多样性的形成和维持机制一直是生态学研究的一个核心问题。经典观点认为物种在群落中的共存是以生态位分化为前提的,换言之,生态位完全重叠的物种可能因为对相同资源产生激烈竞争而发生竞争排斥,因此无法在同一群落中稳定共存[59-60]。虽然近年来生态理论在解释一些限制性资源不多、物种生态位分化不明显、但依然存在稳定共存现象的生物群落时遇到了巨大挑战[61]。但在开阔大洋,水体中垂直方向上的资源条件会随着水深的增加而急剧减少[62],即对于各类浮游动物来说,深海环境通常是一个资源极端受限制的生境。因此,生态位理论对于解释深海浮游动物群落的种间关系仍然是非常恰当的。由于目前对于深海浮游动物群落内部的捕食和寄生等种间关系的研究仍然极度缺乏,故本文主要从种间竞争和生态位分化角度简要阐述深海浮游动物群落的种间关系问题。
对于深海的浮游动物来说,食物生态位分化是至关重要的,而这一过程往往通过空间(水深)生态位的分化而实现。概括而言,研究浮游动物食物生态位的技术手段主要有胃含物分析[63]、脂质分析[64]、分子生物学分析[65]和稳定同位素分析[66]等。其中,随着近年来分析技术的进步,利用脂类标志物和稳定同位素分析研究浮游动物(也包括其他海洋动物)生态位的方法逐渐成为主流趋势[67-68]。例如,Laakmann等[69]对北极深海几种桡足类(包括真刺水蚤科和鹰嘴水蚤科)生态位分化机制的研究表明同一属的近缘种之间通过各自偏好不同的水层深度而实现空间生态位的分化;又如,在南大西洋深海桡足类的研究中发现不同个体的营养级与其栖息的水深成正相关,而且同一物种不同发育阶段个体间和雌雄间都呈现出空间生态位上的显著分化[66]。综上所述,对于深海浮游动物群落而言,较为简单的食物网结构和较为单一的资源限制模式导致近缘种的同域共存必然出现针对食物资源的竞争排斥,个体和种群对低种间竞争环境的偏好导致群落中的不同物种表现出空间(水深)生态位的分化。但不可忽视的是,由于采样调查上的困难,迄今尚未见到有关不同类群和不同水深的深海浮游动物群落面临的捕食压力方面的研究,因此无法评估来源于中层和深层的小型游泳动物及滤食性鱼类对它们的捕食压力在这种空间(水深)生态位分化中的贡献。
2.2 在生物地理学尺度上
传统上,对于生态位方面的研究主要在种群和群落尺度上开展[59, 66],但随着生物地理学、群落生态学和分子系统学的发展和融合,近年来人们逐渐开始在更大的宏观尺度或者生物地理学尺度上探讨物种的生态位变化规律。其中最为重要的发现是“生态位保守性”假说(niche conservatism, NC)[70]的提出。该假说认为某一物种或者某一谱系的生态位及与生态位相关的某些生态特征随着物种或谱系的进化过程而变化不大,即生态位在生物进化过程中具有一定的保守性。大量的研究表明这种现象在陆地动植物中普遍存在[71-72],但是少量试图验证海洋浮游动物生态位保守性假说的研究却发现同一物种不同地理种群间的气候生态位存在显著分化,从而不支持此假说[73]。由于这一假说在海洋浮游动物中的实证研究极度缺乏,因此我们目前无法得到更全面的认识。但是近期一个关于大空间尺度上深海浮游动物分子系统发育关系的研究却有助于我们对上述假说的理解:该研究发现在北极海域和南大洋中层分布的一种桡足类(Gaetanustenuispinus)的不同地理种群间存在显著的遗传分化,但是同样分布于两个海区深层的另一种桡足类(G.brevispinus)的不同地理种群间则不存在显著的遗传分化[74]。这一研究尽管没有获得两个物种不同地理种群间的准确生态位数据,但是根据两个物种栖息深度的不同和不同深度下环境特征稳定性的差异,可以推测海洋深层极端稳定的环境导致物种进化的速度变得非常缓慢,同时海洋深层热盐环流的存在导致即使南北极之间的地理种群间可能并不存在绝对意义上的空间隔离;与之相比,海洋中层的环境特征仍有明显的季节变化和地理差异,这有利于大尺度上不同地理种群间的隔离分化。在此基础上,可以进一步预测海洋浮游动物生态位的保守性程度会随着物种栖息深度的增加而加强,海洋上层分布的浮游动物在同一气候区域内或者同一海区内就可能不具备生态位的保守性;海洋中层分布的浮游动物在更大的地理尺度上也可能不具备生态位的保守性;但海洋深层分布的浮游动物很可能在全球尺度上仍然具有较强的生态位保守性。不过,由于这类大尺度上的实证研究仍然很缺乏,目前这一假说尚无法很好的验证。
3 深海浮游动物群落在碳沉降和生物地球化学循环中的作用
海洋是一个巨大的碳库,有估算认为海洋上层的光合作用固定的碳大约占全球初级生产固定无机碳总量的一半左右[75],而由于生物碳泵(biological carbon pump)的存在,上层海洋固定的碳被大量输送到了深海[76]。有估计甚至认为如果生物碳泵失效,未来大气中的CO2浓度将比现在上升大约50%[77]。因此,在当今大气CO2浓度上升和全球变化的大背景下,海洋生物碳泵及其影响因素得到了各国政府和科学家的高度重视。由于国内已经有关于海洋生物碳泵方面的专门论述[78-79],对于边缘海的浮游生物在海洋生物碳泵方面的研究也有介绍[80],本文着重介绍过去的研究者们相对忽视的两类浮游动物垂直迁移现象对海洋生物碳循环的意义。
3.1 昼夜垂直迁移的意义
很多海洋浮游动物都有昼夜垂直迁移(diel vertical migration, DVM)的习性,通常这种现象表现为夜间上浮至上层或者表层摄食,而白天下沉至数十米至数百米不等的海洋深处[81]。经典观点认为白天的下沉有利于躲避一些依赖于视觉的捕食者,但是也有证据表明在一些类群中躲避捕食并非一个主要原因[26]。这种昼夜垂直迁移已经被证明存在重要生态意义和生物地球化学循环上的意义:一方面,抛开这些昼夜垂直迁移发生机理方面的争论,过去20余年间学术界对于“浮游动物夜晚在表层摄食浮游植物,白天下沉到深层后,其自身呼吸代谢、排泄物和被更高营养级动物捕食都导致它们是上层有机碳往深层输送的一个重要途径”这一基本结论是没有争议的[82-83]。例如,Takahashi等[84]在东北太平洋亚极地海区的研究表明仅仅两种优势桡足类的昼夜垂直迁移对于海洋上层到海洋中层垂直碳通量的贡献率就达到15%之多;而在温带北大西洋的研究也表明上述机制在300 m深度的碳通量贡献可以达到POC通量的71.4%[22]。此外,由于海洋上层初级生产力和不同浮游动物类群昼夜垂直迁移强度的季节差异,浮游动物昼夜垂直迁移对于海洋垂直碳通量的贡献率也会出现明显的季节变化,如在北太平洋亚极地生态系统中这一机制在春季的贡献率就要显著低于其他季节[84]。另一方面,浮游动物的昼夜垂直迁移对于维持海洋中层的浮游生态系统也至关重要:在太平洋热带海区和亚极地海区的研究表明海洋中层损失的POC通量根本无法满足这一深度浮游动物和浮游细菌的能量需求,仅仅细菌的碳需求就达到同水层POC损失量的3—10倍之多,两者之间如此大的差距只能依赖浮游动物垂直迁移带来的能量和物质补充[85]。
3.2 季节性垂直迁移的意义
很多大型的桡足类存在季节性垂直迁移(seasonal vertical migration, SVM)现象,这种现象往往伴随着个体的发育过程,例如北太平洋亚极地海区的一些大型哲水蚤会在上层海洋摄食和生长,待个体成熟后下沉到数百米的深海交配和产卵,并且不会再次上浮到海洋上层[86];而极地海区的一些种类则要经历数年的发育期才能够成熟,期间会多次经历季节性垂直迁移过程[87],因此这种垂直迁移又被称为“个体发育垂直迁移”(ontogenetically vertical migration)。例如,在北太平洋亚极地海区,几种优势的新哲水蚤和真哲水蚤在浮游植物旺发季节(4—7月)会一直停留在海洋上层摄食,随着秋季温度下降和浮游植物生物量的急剧降低,它们逐渐下沉到海洋中层(8—9月),然后冬季会停留在那个深度休眠[88],而据估计仅其中一种(Neocalanusflemingeri)的季节性垂直迁移活动为深海(1000 m深度)带来的垂直碳通量贡献就可达到总POC通量的9%之多[89]。与之类似,在北大西洋高纬度海区,春夏季积累了丰富脂类物质的飞马哲水蚤Calanusfinmarchicus种群会在夏末逐渐下沉到600—1400 m的深海中进入休眠[86, 90],尽管每只哲水蚤平均仅含有200 μg的脂类,但由于其拥有极大的种群规模,因此这一季节性垂直迁移意味着巨量的脂类物质下沉到了深海之中。最近的一项研究计算了休眠种群的死亡率、代谢消耗和排泄之后认为大约有44%—93%的脂类物质最终在种群休眠的深海中被消耗掉,这意味着1—4 g C m-2a-1的深海碳通量[91]。
关于浮游动物垂直迁移在生物地球化学循环中的意义,当前的研究主要还存在几方面的不足:一是人们对中层浮游动物面临的捕食压力很难准确估算,而遭受捕食通常是海洋浮游桡足类最主要的死亡原因[92],因此这导致很难准确区分浮游动物自身呼吸代谢、排泄物和个体死亡三者对于这一过程的相对贡献(例如有研究认为呼吸代谢的贡献要占绝对优势(64%—98%)[84],但也有结果表明排泄物的贡献要大于呼吸代谢[89]);二是目前的研究主要涉及海洋上层和中层,而一些极地海区浮游动物的季节性垂直迁移可以达到1000 m以下的海洋深层[23],但迄今关于深层浮游动物的垂直迁移与生物碳泵之间关系的研究仍然极度缺乏;三是目前在高生产力海区(如南大洋、白令海、北大西洋的亚极地海区等)和中等生产力海区(如热带东太平洋海区)的研究较多,而在寡营养海区(如热带西太平洋暖池区和北太平洋亚热带环流区等)的研究很少。
4 当前深海浮游生态学研究的技术手段简介
4.1 浮游生物拖网
浮游生物拖网是最传统和最为广泛使用的浮游动物采样调查设备。早在1828年,人们就开始利用浮游生物拖网采集浮游生物,此后达尔文在乘坐“贝格尔”号进行环球考察时也使用过此类设备[4]。随着对海洋科学认识的深入,人们逐渐开始试图发展一些能够深入到海洋深处,并且分不同水层采集浮游动物样品的设备。当前,深海调查时应用较多的浮游生物分层拖网主要包括BIONESS(Bedford Institute of Oceanography Net and Environmental Sampling System)、MOCNESS(Multiple Opening/Closing Net and Environmental Sensing System)和HYDRO-BIOS Multinet三类,其主要性能指标见表1。
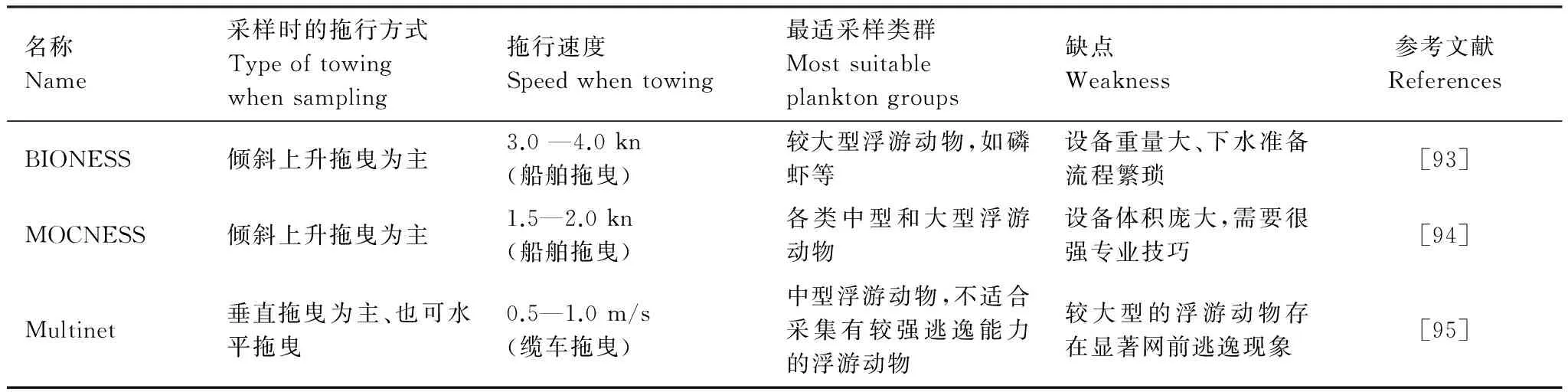
表1 当前深海浮游生物调查中应用广泛的三类主要分层拖网简介
4.2 光学调查设备
由于经典的网采-镜检模式无法快速高效获得浮游动物群落的生态数据,因此人们开发了一系列基于各类光学技术的浮游动物调查设备,当前应用较多的包括VPR(Video Plankton Recorder)、OPC(Optical Plankton Counter)和LOPC(Laser Optical Plankton Counter)、UVP(Underwater Video Profiler)微光成像系统以及搭载在AUV/ROV/HOV等各类潜器上的摄录设备等。关于海洋浮游动物的图像观测技术及相关进展,国内近期也有专门报道[96]。
4.3 声学调查设备
在各类海洋调查仪器中,基于声学原理的调查设备是很重要的一个类别。其中,声学多普勒海流剖面仪(Acoustic Doppler Current Profiler, ADCP)是一类基础的物理海洋调查设备,由于其测定海流的基本原理是捕捉水中各类悬浮粒子的声波反射,因此这类设备可以用于研究不同粒径浮游动物在水体中的丰度和分布情况[97]。声学调查方法的主要优势有二:首先,由于当前各类ADCP已经广泛装备于各式海洋调查船、锚定长期观测系统及CTD采水器等设备平台,因此可以通过分析这些ADCP数据获取经典网采调查方法很难获得的长期性、大范围的浮游动物调查数据[98];其次,可以很方便的进行时间上的连续站或者空间上的连续剖面调查,并实时得到调查数据[28]。
4.4 沉积物捕获器
现代的沉积物捕获器(Sediment Trap)通常是为采集特定水深、长时间序列的海洋沉降颗粒物样品而设计的装置,目前已经成为全球变化研究和生物地球化学循环研究领域非常重要的技术手段[99]。由于很多中层和深层的浮游动物具有较强的垂直迁移能力,而沉积物捕获器的设计又导致其一旦落入样品管则很难再次逃逸,因此沉积物捕获器的时间序列沉积物样品中往往混杂有数量不一的浮游动物样品[100]。尽管沉积物捕获器获取的浮游动物样品毫无疑问具有很大的随机性和选择性,但是在高生产力的海区,仍可与浮游生物拖网获取的数据结合来分析浮游动物群落的季节变化规律[101]。
5 我国的研究现状
长期以来,我国浮游生态学研究的重点区域是近海和边缘海的上层水体,而由于经费和技术手段的限制,对边缘海和大洋的中层及深层浮游动物生态学研究很少。早期较有代表性的工作是在20世纪70年代至80年代对南海的综合调查中,国家海洋局第三海洋研究所和中国科学院南海海洋研究所分别对这一海区中层和深层的浮游动物生物量和物种多样性等基本群落特征进行了初步研究[102- 104]。此后,随着国家南北极综合科考工作的展开,我国科学家分别对北冰洋的楚科奇海、北太平洋的白令海及南大洋的南设得兰岛附近海域上层和中层的浮游动物群落结构进行了研究[105- 107]。近年来,随着我国“一带一路”国家战略及“走向深远海”海洋发展战略的实施,我国科学家依托国家海洋局“全球变化与海气相互作用”专项、中国科学院“热带西太平洋海洋系统物质能量交换及其影响”先导专项、中国大洋矿产资源研究开发协会“中国富钴结壳勘探合同区的资源与环境调查”专项及“中国多金属结核勘探合同区的资源与环境调查”专项、科技部“超深渊生物群落及其与关键环境要素的相互作用机制研究”973项目等重大科技计划的资助,逐渐开展了一系列边缘海、大洋和极地海域的深海浮游动物生态学研究,特别是对西北太平洋的马里亚纳海沟、雅浦海沟、采薇海山和雅浦海山等区域展开了较为系统的浮游生态学研究。
6 展望
(1)在深海浮游动物群落垂直分布及其变化方面,已有的研究报道主要集中在北半球的极地亚极地、南大洋、东太平洋和阿拉伯海等区域,对于与我国关系最密切的西太平洋海区,这类研究还非常缺乏。此外,在我国南海海盆和东海的深水区,大量的研究表明上层浮游生态系统会受到季风和环流的显著影响[12-13, 108],基于上文所述阿拉伯海区域的研究结果,中层和深层生态系统也会受到相应的影响。
(2)在深海浮游动物群落的种间关系和生态位分化方面,早期受到稳定同位素分析技术上的限制,必须通过先确定基准营养级的稳定同位素基线,然后测定目标动物的δ15N并与之比对,从而获得目标动物或类群的营养级[69, 109]。这一方法的最大缺陷在于人为选取的基准营养级有时会与真实情况存在显著偏差,而近期氨基酸特异性稳定同位素分析方法的进步已经可以有效避免这个问题[110]。因此,这一方法的应用必然极大促进今后深海浮游动物种间关系的研究。
(3)在深海浮游动物与生物地球化学循环的研究方面,已有的研究主要集中在生产力较高的热带、亚热带边缘海和高纬度海区,对于广大寡营养海区的研究极为缺乏。在当前全球气候变化和人类活动的大背景下,寡营养海区(如热带亚热带的西太平洋、中太平洋和中印度洋等)生态系统的重要性日益受到重视[111-112],今后在这些海域的研究应更多的考虑深海浮游动物生态学与生物地球化学循环和全球变化研究的紧密结合,有针对性的开展工作。
[1] 冯士筰, 李凤岐, 李少菁. 海洋科学导论. 北京: 高等教育出版社, 1999.
[2] Duxbury A C, Duxbury A B, Sverdrup K A. An Introduction to the World′s Oceans. 6th ed. New York, USA: McGraw-Hill, 2000.
[3] Hardy A C. A new method of plankton research. Nature, 1926: 118: 630- 632.
[4] Fraser J H. The history of plankton sampling // Tranter D J, ed. Zooplankton Sampling. Paris: UNESCO, 1968: 11- 18.
[5] Ayón P, Criales-Hernandez M I, Schwamborn R, Hirche H J. Zooplankton research off Peru: a review. Progress in Oceanography, 2008, 79(2/4): 238- 255.
[6] 孙松. 中国区域海洋学: 生物海洋学. 北京: 海洋出版社, 2012.
[7] Buesseler K O, Lamborg C H, Boyd P W, Lam P J, Trull T W, Bidigare R R, Bishop J K, Casciotti K L, Dehairs F, Elskens M, Honda M, Karl D M, Siegel D A, Silver M W, Steinberg D K, Valdes J, Van Mooy B, Wilson S. Revisiting carbon flux through the ocean′s twilight zone. Science, 2007, 316(5824): 567- 570.
[8] Ikeda T, Sano F, Yamaguchi A. Respiration in marine pelagic copepods: a global-bathymetric model. Marine Ecology Progress Series, 2007, 339: 215- 219.
[9] 秦蕴珊, 尹宏. 西太平洋——我国深海科学研究的优先战略选区. 地球科学进展, 2011, 26(3): 245- 248.
[10] Hunt G L Jr, Stabeno P, Walters G, Sinclair E, Brodeur R D, Napp J M, Bond N A. Climate change and control of the southeastern Bering Sea pelagic ecosystem. Deep Sea Research Part II: Topical Studies in Oceanography, 2002, 49(26): 5821- 5853.
[11] Sambrotto R N, Niebauer H J, Goering J J, Iverson R L. Relationships among vertical mixing, nitrate uptake, and phytoplankton growth during the spring bloom in the southeast Bering Sea middle shelf. Continental Shelf Research, 1986, 5(1/2): 161- 198.
[12] Tang D L, Kawamura H, Van Dien T, Lee M. Offshore phytoplankton biomass increase and its oceanographic causes in the South China Sea. Marine Ecology Progress Series, 2004, 268: 31- 41.
[13] Li K Z, Yin J Q, Huang L M, Zhang J L, Lian S M, Liu C M. Distribution and abundance of thaliaceans in the northwest continental shelf of South China Sea, with response to environmental factors driven by monsoon. Continental Shelf Research, 2011, 31(9): 979- 989.
[14] Madhupratap M, Gopalakrishnan T C, Haridas P, Nair K K. Mesozooplankton biomass, composition and distribution in the Arabian Sea during the fall intermonsoon: implications of oxygen gradients. Deep Sea Research Part II: Topical Studies in Oceanography, 2001, 48(5/7): 1345- 1368.
[15] Menzies R J. Conditions for the existence of life on the abyssal sea floor. Oceanography and Marine Biology: An Annual Review, 1965, 3: 195- 210.
[16] Smith K L Jr, Kaufmann R S, Baldwin R J, Carlucci A F. Pelagic-benthic coupling in the abyssal eastern North Pacific: an 8-year time-series study of food supply and demand. Limnology and Oceanography, 2001, 46(3): 543- 556.
[17] 陈兴群, 林荣澄. 东北太平洋中国合同区的叶绿素a和初级生产力. 海洋学报, 2007, 29(5): 146- 153.
[18] Steinberg D K, Cope J S, Wilson S E, Kobari T. A comparison of mesopelagic mesozooplankton community structure in the subtropical and subarctic North Pacific Ocean. Deep Sea Research Part II: Topical Studies in Oceanography, 2008, 55(14- 15): 1615- 1635.
[19] Siegel D A, Doney S C, Yoder J A. The North Atlantic spring phytoplankton bloom and Sverdrup′s critical depth hypothesis. Science, 2002, 296(5568): 730- 733.
[20] Siokou I, Zervoudaki S, Christou E D. Mesozooplankton community distribution down to 1000 m along a gradient of oligotrophy in the Eastern Mediterranean Sea (Aegean Sea). Journal of Plankton Research, 2013, 35(6): 1313- 1330.
[21] Azam F, Long R A. Oceanography: sea snow microcosms. Nature, 2001, 414(6863): 495- 498.
[22] Steinberg D K, Carlson C A, Bates N R, Goldthwait S A, Madin L P, Michaels A F. Zooplankton vertical migration and the active transport of dissolved organic and inorganic carbon in the Sargasso Sea. Deep Sea Research Part I: Oceanographic Research Papers, 2000, 47(1): 137- 158.
[23] Koppelmann R, Weikert H. Temporal changes of deep-sea mesozooplankton abundance in the temperate NE Atlantic and estimates of the carbon budget. Marine Ecology Progress Series, 1999, 179: 27- 40.
[24] Schandelmeier L, Alexander V. An analysis of the influence of ice on spring phytoplankton population structure in the southeast Bering Sea. Limnology and Oceanography, 1981, 26(5): 935- 943.
[25] Hagen W, Auel H. Seasonal adaptations and the role of lipids in oceanic zooplankton. Zoology, 2001, 104(3/4): 313- 326.
[26] Pearre S Jr. Eat and run? The hunger/satiation hypothesis in vertical migration: history, evidence and consequences. Biological Reviews, 2003, 78(1): 1- 79.
[27] Gislason A, Silva T. Abundance, composition, and development of zooplankton in the Subarctic Iceland Sea in 2006, 2007, and 2008. ICES Journal of Marine Science, 2012, 69(7): 1263- 1276.
[28] Valle-Levinson A, Castro L, Cáceres M, Pizarro O. Twilight vertical migrations of zooplankton in a Chilean fjord. Progress in Oceanography, 2014, 129: 114- 124.
[29] Blachowiak-Samolyk K, Kwasniewski S, Richardson K, Dmoch K, Hansen E, Hop H, Falk-Petersen S, Mouritsen L T. Arctic zooplankton do not perform diel vertical migration (DVM) during periods of midnight sun. Marine Ecology Progress Series, 2006, 308: 101- 116.
[30] Ji R B, Ashjian C J, Campbell R G, Chen C S, Gao G P, Davis C S, Cowles G W, Beardsley R C. Life history and biogeography ofCalanuscopepods in the Arctic Ocean: an individual-based modeling study. Progress in Oceanography, 2012, 96(1): 40- 56.
[31] Fiedler P C, Talley L D. Hydrography of the eastern tropical Pacific: a review. Progress in Oceanography, 2006, 69(2/4): 143- 180.
[32] Pennington J T, Mahoney K L, Kuwahara V S, Kolber D D, Calienes R, Chavez F P. Primary production in the eastern tropical Pacific: a review. Progress in Oceanography, 2006, 69(2/4): 285- 317.
[33] Stramma L, Schmidtko S, Levin L A, Johnson G C. Ocean oxygen minima expansions and their biological impacts. Deep Sea Research Part I: Oceanographic Research Papers, 2010, 57(4): 587- 595.
[34] Childress J J, Seibel B A. Life at stable low oxygen levels: adaptations of animals to oceanic oxygen minimum layers. The Journal of Experimental Biology, 1998, 201(8): 1223- 1232.
[35] Wishner K F, Gowing M M, Gelfman C. Mesozooplankton biomass in the upper 1000 m in the Arabian Sea: overall seasonal and geographic patterns, and relationship to oxygen gradients. Deep Sea Research Part II: Topical Studies in Oceanography, 1998, 45(10/11): 2405- 2432.
[36] Fabian H, Koppelmann R, Weikert H. Full-depths zooplankton composition at two deep sites in the western and central Arabian Sea. Indian Journal of Marine Sciences, 2005, 34(2): 174- 187.
[37] Saltzman J, Wishner K F. Zooplankton ecology in the eastern tropical Pacific oxygen minimum zone above a seamount: 2. Vertical distribution of copepods. Deep Sea Research Part I: Oceanographic Research Papers, 1997, 44(6): 931- 954.
[38] Saltzman J, Wishner K F. Zooplankton ecology in the eastern tropical Pacific oxygen minimum zone above a seamount: 1. General trends. Deep Sea Research Part I: Oceanographic Research Papers, 1997, 44(6): 907- 930.
[39] Liu K K, Chao S Y, Shaw P T, Gong G C, Chen C C, Tang T Y. Monsoon-forced chlorophyll distribution and primary production in the South China Sea: observations and a numerical study. Deep Sea Research Part I: Oceanographic Research Papers, 2002, 49(8): 1387- 1412.
[40] Qu T D. Upper-layer circulation in the South China Sea. Journal of Physical Oceanography, 2000, 30(6): 1450- 1460.
[41] Schott F. Monsoon response of the Somali Current and associated upwelling. Progress in Oceanography, 1983, 12(3): 357- 381.
[42] Luo J G, Ortner P B, Forcucci D, Cummings S R. Diel vertical migration of zooplankton and mesopelagic fish in the Arabian Sea. Deep Sea Research Part II: Topical Studies in Oceanography, 2000, 47(7/8): 1451- 1473.
[43] Honjo S, Dymond J, Prell W, Ittekkot V. Monsoon-controlled export fluxes to the interior of the Arabian Sea. Deep Sea Research Part II: Topical Studies in Oceanography, 1999, 46(8/9): 1859- 1902.
[44] Koppelmann R, Fabian H, Weikert H. Temporal variability of deep-sea zooplankton in the Arabian Sea. Marine Biology, 2003, 142(5): 959- 970.
[45] Koppelmann R, Frost J. The ecological role of zooplankton in the twilight and dark zones of the ocean // Mertens L P, ed. Biological Oceanography Research Trends. New York: Nova Science Publishers, Inc., 2008: 67- 130.
[46] Menard H W. Marine Geology of the Pacific. New York: McGraw-Hill, 1964: 271- 271.
[47] Genin A. Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies. Journal of Marine Systems, 2004, 50(1/2): 3- 20.
[48] Rogers A D. The biology of seamounts. Advances in Marine Biology, 1993, 30: 305- 305.
[49] Boehlert G W, Genin A. A review of the effects of seamounts on biological processes // Keating B H, Fryer P, Batiza R, Boehlert G W, eds. Seamounts, Islands, and Atolls. Washington, DC: American Geophysical Union, 1987: 319- 334.
[50] Dower J F, Mackas D L. “Seamount effects” in the zooplankton community near Cobb Seamount. Deep Sea Research Part I: Oceanographic Research Papers, 1996, 43(6): 837- 858.
[51] Comeau L A, Vézina A F, Bourgeois M, Juniper S K. Relationship between phytoplankton production and the physical structure of the water column near Cobb Seamount, northeast Pacific. Deep Sea Research Part I: Oceanographic Research Papers, 1995, 42(6): 993- 1005.
[52] Martin B, Christiansen B. Distribution of zooplankton biomass at three seamounts in the NE Atlantic. Deep Sea Research Part II: Topical Studies in Oceanography, 2009, 56(25): 2671- 2682.
[53] Uda M, Ishino M. Enrichment pattern resulting from eddy systems in relation to fishing grounds. Journal of the Tokyo University of Fisheries, 1958, 44(1/2): 105- 129.
[54] Genin A, Greene C, Haury L, Wiebe P, Gal G, Kaartvedt S, Meir E, Fey C, Dawson J. Zooplankton patch dynamics: daily gap formation over abrupt topography. Deep Sea Research Part I: Oceanographic Research Papers, 1994, 41(5/6): 941- 951.
[55] Hirch S, Martin B, Christiansen B. Zooplankton metabolism and carbon demand at two seamounts in the NE Atlantic. Deep Sea Research Part II: Topical Studies in Oceanography, 2009, 56(25): 2656- 2670.
[56] Franks P J S. Sink or swim: accumulation of biomass at fronts. Marine Ecology Progress Series, 1992, 82(1): 1- 12.
[57] Dower J, Freeland H, Juniper K. A strong biological response to oceanic flow past Cobb Seamount. Deep Sea Research Part A. Oceanographic Research Papers, 1992, 39(7/8): 1139- 1145.
[58] 张武昌, 于莹, 李超伦, 李学刚, 肖天. 海山区浮游生态学研究. 海洋与湖沼, 2014, 45(5): 973- 978.
[59] Hutchinson G E. Homage to Santa Rosalia or why are there so many kinds of animals?. The American Naturalist, 1959, 93(870): 145- 159.
[60] Pianka E R. Competition and niche theory // May R M, ed. Theoretical Ecology: Principles and Applications. 2nd ed. Oxford: Blackwell, 1981: 167- 196.
[61] 牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579- 593.
[62] Karl D M, Knauer G A, Martin J H. Downward flux of particulate organic matter in the ocean: a particle decomposition paradox. Nature, 1988, 332(6163): 438- 441.
[63] Maly E J, Maly M P. Dietary differences between two co-occurring calanoid copepod species. Oecologia, 1974, 17(4): 325- 333.
[64] Auel H, Harjes M, da Rocha R, Stübing D, Hagen W. Lipid biomarkers indicate different ecological niches and trophic relationships of the Arctic hyperiid amphipodsThemistoabyssorumandT.libellula. Polar Biology, 2002, 25(5): 374- 383.
[65] Nejstgaard J C, Frischer M E, Simonelli P, Troedsson C, Brakel M, Adiyaman F, Sazhin A F, Artigas L F. Quantitative PCR to estimate copepod feeding. Marine Biology, 2008, 153(4): 565- 577.
[66] Laakmann S, Auel H. Longitudinal and vertical trends in stable isotope signatures (δ13C and δ15N) of omnivorous and carnivorous copepods across the South Atlantic Ocean. Marine Biology, 2010, 157(3): 463- 471.
[67] Chikaraishi Y, Steffan S A, Ogawa N O, Ishikawa N F, Sasaki Y, Tsuchiya M, Ohkouchi N. High-resolution food webs based on nitrogen isotopic composition of amino acids. Ecology and Evolution, 2014, 4(12): 2423- 2449.
[68] Pakhomov E A, McClelland J W, Bernard K, Kaehler S, Montoya J P. Spatial and temporal shifts in stable isotope values of the bottom-dwelling shrimpNauticarismarionisat the sub-Antarctic archipelago. Marine Biology, 2004, 144(2): 317- 325.
[69] Laakmann S, Kochzius M, Auel H. Ecological niches of Arctic deep-sea copepods: vertical partitioning, dietary preferences and different trophic levels minimize inter-specific competition. Deep Sea Research Part I: Oceanographic Research Papers, 2009, 56(5): 741- 756.
[70] Losos J B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecology Letters, 2008, 11(10): 995- 1003.
[71] Peterson A T, Soberón J, Sánchez-Cordero V. Conservatism of ecological niches in evolutionary time. Science, 1999, 285(5431): 1265- 1267.
[72] Tingley M W, Monahan W B, Beissinger S R, Moritz C. Birds track their Grinnellian niche through a century of climate change. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(S2): 19637- 19643.
[73] Kelly M W, Sanford E, Grosberg R K. Limited potential for adaptation to climate change in a broadly distributed marine crustacean. Proceedings of the Royal Society of London B: Biological Sciences, 2012, 279(1727): 349- 356.
[74] Laakmann S, Auel H, Kochzius M. Evolution in the deep sea: biological traits, ecology and phylogenetics of pelagic copepods. Molecular Phylogenetics and Evolution, 2012, 65(2): 535- 546.
[75] Field C B, Behrenfeld M J, Randerson J T, Falkowski P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science, 1998, 281(5374): 237- 240.
[76] Sanders R, Henson S A, Koski M, De La Rocha C L, Painter S C, Poulton A J, Riley J, Salihoglu B, Visser A, Yool A, Bellerby R, Martin A P. The biological carbon pump in the North Atlantic. Progress in Oceanography, 2014, 129: 200- 218.
[77] Parekh P, Dutkiewicz S, Follows M, Ito T. Atmospheric carbon dioxide in a less dusty world. Geophysical Research Letters, 2006, 33(3): L03610.
[78] 黄邦钦, 胡俊, 柳欣, 王磊, 张彩云, 周理斌. 全球气候变化背景下浮游植物群落结构的变动及其对生物泵效率的影响. 厦门大学学报: 自然科学版, 2011, 50(2): 402- 410.
[79] 焦念志, 张传伦, 李超, 王晓雪, 党宏月, 曾庆璐, 张锐, 张瑶, 汤凯, 张子莲, 徐大鹏. 海洋微型生物碳泵储碳机制及气候效应. 中国科学: 地球科学, 2013, 43(1): 1- 18.
[80] 黄邦钦, 柳欣. 边缘海浮游生态系统对生物泵的调控作用. 地球科学进展, 2015, 30(3): 385- 395.
[81] 张武昌. 浮游动物的昼夜垂直迁移. 海洋科学, 2000, 24(11): 18- 21.
[82] Longhurst A R, Bedo A W, Harrison W G, Head E L H, Sameoto D D. Vertical flux of respiratory carbon by oceanic diel migrant biota. Deep Sea Research Part A. Oceanographic Research Papers, 1990, 37(4): 685- 694.
[83] Passow U, Carlson C A. The biological pump in a high CO2world. Marine Ecology Progress Series, 2012, 470: 249- 271.
[84] Takahashi K, Kuwata A, Sugisaki H, Uchikawa K, Saito H. Downward carbon transport by diel vertical migration of the copepodsMetridiapacificaandMetridiaokhotensisin the Oyashio region of the western subarctic Pacific Ocean. Deep Sea Research Part I: Oceanographic Research Papers, 2009, 56(10): 1777- 1791.
[85] Steinberg D K, Van Mooy B A S, Buesseler K O, Boyd P W, Kobari T, Karl D M. Bacterial vs. zooplankton control of sinking particle flux in the ocean′s twilight zone. Limnology and Oceanography, 2008, 53(4): 1327- 1338.
[86] Conover R J. Comparative life histories in the generaCalanusandNeocalanusin high latitudes of the northern hemisphere. Hydrobiologia, 1988, 167- 168(1): 127- 142.
[87] Dawson J K. Vertical distribution ofCalanushyperboreusin the central Arctic Ocean. Limnology and Oceanography, 1978, 23(5): 950- 957.
[88] Miller C B, Clemons M J. Revised life history analysis for large grazing copepods in the subarctic Pacific Ocean. Progress in Oceanography, 1988, 20(4): 293- 313.
[89] Kobari T, Steinberg D K, Ueda A, Tsuda A, Silver M W, Kitamura M. Impacts of ontogenetically migrating copepods on downward carbon flux in the western subarctic Pacific Ocean. Deep Sea Research Part II: Topical Studies in Oceanography, 2008, 55(14- 15): 1648- 1660.
[90] Lee R F, Hagen W, Kattner G. Lipid storage in marine zooplankton. Marine Ecology Progress Series, 2006, 307: 273- 306.
[91] Jónasdóttir S H, Visser A W, Richardson K, Heath M R. Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(39): 12122- 12126.
[92] Hirst A G, Kiørboe T. Mortality of marine planktonic copepods: global rates and patterns. Marine Ecology Progress Series, 2002, 230: 195- 209.
[93] Sameoto D, Jaroszynski L O, Fraser W B. BIONESS, a new design in multiple net zooplankton samplers. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(4): 722- 724.
[94] Wiebe P H, Morton A W, Bradley A M, Backus R H, Craddock J E, Barber V, Cowles T J, Flierl G R. New development in the MOCNESS, an apparatus for sampling zooplankton and micronekton. Marine Biology, 1985, 87(3): 313- 323.
[95] Weikert H, John H C. Experiences with a modified Bé multiple opening-closing plankton net. Journal of Plankton Research, 1981, 3(2): 167- 176.
[96] 孙晓霞, 孙松. 海洋浮游生物图像观测技术及其应用. 地球科学进展, 2014, 29(6): 748- 755.
[97] Heywood K J, Scrope-Howe S, Barton E. Estimation of zooplankton abundance from shipborne ADCP backscatter. Deep Sea Research Part A. Oceanographic Research Papers, 1991, 38(6): 677- 691.
[98] Postel L, da Silva A J, Mohrholz V, Lass H U. Zooplankton biomass variability off Angola and Namibia investigated by a lowered ADCP and net sampling. Journal of Marine Systems, 2007, 68(1/2): 143- 166.
[99] 陈建芳, 郑连福. 沉积物捕获器与全球变化研究. 海洋通报, 1996, 15(1): 41- 47.
[100] Ivory J A, Tang K W, Takahashi K. Use of Neutral Red in short-term sediment traps to distinguish between zooplankton swimmers and carcasses. Marine Ecology Progress Series, 2014, 505: 107- 117.
[101] Matsuno K, Yamaguchi A, Fujiwara A, Onodera J, Watanabe E, Harada N, Kikuchi T. Seasonal changes in the population structure of dominant planktonic copepods collected using a sediment trap moored in the western Arctic Ocean. Journal of Natural History, 2015, 49(45- 48): 2711- 2726.
[102] 陈清潮, 张谷贤, 陈柏云. 西沙、中沙群岛周围海域浮游动物的平面分布和垂直分布 // 中国科学院南海海洋研究所. 我国西沙、中沙群岛海域海洋生物调查研究报告集. 北京: 科学出版社, 1978: 75- 80.
[103] 陈瑞祥, 蔡秉及, 林茂, 戴燕玉, 林景宏, 连光山. 南海中部海域浮游动物的垂直分布. 海洋学报, 1988, 10(3): 337- 341.
[104] 陈柏云. 西沙、中沙群岛海洋浮游桡足类的种类组成和分布. 厦门大学学报: 自然科学版, 1982, 21(2): 209- 217.
[105] 林景宏, 戴燕玉, 张金标, 林茂, 杨清良, 林更铭. 夏季楚科奇海浮游动物的生态特征. 极地研究, 2001, 13(2): 107- 116.
[106] 林景宏, 戴燕玉, 林茂, 杨清良, 张金标, 林更铭. 夏季白令海浮游动物的分布. 极地研究, 2002, 14(2): 126- 135.
[107] 杨光, 李超伦, 张永山, 刘群. 南极夏季南设得兰岛海域浮游动物群落垂直分布. 极地研究, 2015, 27(1): 17- 24.
[108] Hwang J S, Dahms H U, Tseng L C, Chen Q C. Intrusions of the Kuroshio Current in the northern South China Sea affect copepod assemblages of the Luzon Strait. Journal of Experimental Marine Biology and Ecology, 2007, 352(1): 12- 27.
[109] Koppelmann R, Böttger-Schnack R, Möbius J, Weikert H. Trophic relationships of zooplankton in the eastern Mediterranean based on stable isotope measurements. Journal of Plankton Research, 2009, 31(6): 669- 686.
[110] Hannides C C S, Popp B N, Landry M R, Graham B S. Quantification of zooplankton trophic position in the North Pacific Subtropical Gyre using stable nitrogen isotopes. Limnology and Oceanography, 2009, 54(1): 50- 61.
[111] Bell J D, Ganachaud A, Gehrke P C, Griffiths S P, Hobday A J, Hoegh-Guldberg O, Johnson J E, Le Borgne R, Lehodey P, Lough J M, Matear R J, Pickering T D, Pratchett M S, Gupta A S, Senina I, Waycott M. Mixed responses of tropical Pacific fisheries and aquaculture to climate change. Nature Climate Change, 2013, 3(6): 591- 599.
[112] Doney S C, Ruckelshaus M, Duffy J E, Barry J P, Chan F, English C A, Galindo H M, Grebmeier J M, Hollowed A B, Knowlton N, Polovina J, Rabalais N N, Sydeman W J, Talley L D. Climate change impacts on marine ecosystems. Annual Review of Marine Science, 2012, 4: 11- 37.
A review of open ocean zooplankton ecology
SUN Dong1, WANG Chunsheng1,2,*
1LaboratoryofMarineEcosystemandBiogeochemistry,SecondInstituteofOceanography,StateOceanicAdministration,Hangzhou310012,China2StateKeyLaboratoryofSatelliteOceanEnvironmentDynamics,SecondInstituteofOceanography,StateOceanicAdministration,Hangzhou310012,China
The open-sea ecosystem is usually divided into several subsystems: epipelagic, mesopelagic, bathypelagic, abyssopelagic zones, etc. Many findings indicated that plankton communities showed significant temporal and spatial variations in the epipelagic zone, mainly because of the variations of light, temperature, ice, nutrient supplements, etc. However, in the mesopelagic and deeper zones, more stable physical and chemical environments with increased depths resulted in weaker temporal and spatial variations of ecosystems than those in the epipelagic zone. Besides, our knowledge about the general patterns of the temporal and spatial variation of deep-sea ecosystems has been seriously restricted by limited investigation technology and funding. With the development of marine science and the implementation of marine strategies, the focus of our marine research has evolved from the limited neritic area to coordination of neritic, marginal, and open seas. However, China′s scientific contribution to deep-sea ecology, especially zooplankton ecology of the mesopelagic, bathypelagic, and abyssopelagic zones, has been more limited than that of the neritic zone in the past several decades. The present paper provides an overview of the current progress of deep-sea zooplankton ecology, including the vertical distribution and variation of deep-sea zooplankton communities, interspecific relationships, and niche differentiation, and their roles in carbon deposition and the biogeochemical cycle, etc. Furthermore, the investigation technology of deep-sea zooplankton ecology has developed remarkably in recent years.
plankton ecosystem; vertical distribution; hypoxia; monsoon; seamount; biogeochemical cycle
国家重点基础研究发展计划(2015CB755902);国家重点研发计划 (2016YFC0304105); 国家自然科学基金(41406116)
2016- 03- 06; 网络出版日期:2017- 02- 17
10.5846/stxb201603060393
*通讯作者Corresponding author.E-mail: wangsio@sio.org.cn
孙栋,王春生.深远海浮游动物生态学研究进展.生态学报,2017,37(10):3219- 3231.
Sun D, Wang C S.A review of open ocean zooplankton ecology.Acta Ecologica Sinica,2017,37(10):3219- 3231.
猜你喜欢
潍坊学院学报(2020年2期)2021-01-18
Plasma Science and Technology(2020年9期)2020-09-14
空间科学学报(2020年2期)2020-04-01
流行色(2019年10期)2019-12-06
37°女人(2019年4期)2019-04-20
作品(2016年10期)2016-12-06
大江南北(2016年3期)2016-11-22
环境科技(2016年2期)2016-11-08
现代企业(2015年6期)2015-02-28
华东师范大学学报(自然科学版)(2014年4期)2014-03-11
