
跑台运动对肥胖大鼠比目鱼肌毛细血管生成及apelin表达的影响
2017-06-09 08:58:45张靓马谨陈雪飞董泽源
中国运动医学杂志 2017年5期
张靓 马谨 陈雪飞 董泽源
北京师范大学体育与运动学院(北京 100875)
跑台运动对肥胖大鼠比目鱼肌毛细血管生成及apelin表达的影响
张靓 马谨 陈雪飞 董泽源
北京师范大学体育与运动学院(北京 100875)
目的:观察跑台运动对高脂饮食诱导的肥胖大鼠骨骼肌毛细血管生成及apelin/APJ表达的影响。方法:5周龄健康雄性SD大鼠随机分成对照组和高脂喂养组,分别进行普通饲料和高脂饲料喂养,12周后筛选肥胖大鼠16只,随机分为高脂安静组和高脂运动组。跑台训练持续10周,每周5次,每次60分钟,跑速26米/分钟。测定大鼠体重、体脂重及血脂水平,免疫组化法观察比目鱼肌毛细血管密度和apelin蛋白表达水平,放免法测定比目鱼肌apelin含量,real-time PCR方法测定比目鱼肌apelin和APJ的mRNA表达。结果:与对照组相比,高脂饮食喂养大鼠体重和体脂显著增加(P<0.01),总甘油三酯、总胆固醇和低密度脂蛋白胆固醇水平显著升高,高密度脂蛋白胆固醇水平显著降低(P<0.05)。与对照组比,肥胖大鼠比目鱼肌毛细血管生成无显著变化,apelin和APJ mRNA表达无显著变化(P>0.05)。与单纯高脂饮食组大鼠相比,10周跑台运动大鼠体重显著降低(P<0.01),血脂改善;此外,比目鱼肌毛细血管密度显著升高,apelin和APJ mRNA表达显著上升(P<0.05),apelin蛋白表达水平显著升高(P<0.05)。结论:跑台运动能显著增加肥胖大鼠比目鱼肌毛细血管生成,其作用可能与运动诱导比目鱼肌apelin表达相关。
跑台运动;毛细血管密度;apelin;肥胖;比目鱼肌
骨骼肌毛细血管是骨骼肌发挥正常功能的关键组分,机体通过毛细血管,给肌纤维提供氧气和营养成分,并带走二氧化碳及代谢废物。骨骼肌重量占人体体重的40%,骨骼肌毛细血管的改变对机体的代谢、内分泌及运动能力均有着重要的影响[1]。骨骼肌毛细血管受多种生理及病理生理因素的影响,其中肥胖引起的骨骼肌毛细血管密度减少、毛细血管募集能力降低已被多篇文献所报道[2]。运动作为一种生理性因素,对骨骼肌毛细血管的生成有强大的促进作用。运动时骨骼肌毛细血管的血流量比安静时升高100倍,血管内皮承受的剪切力增加,骨骼肌反复收缩和舒张引起的牵拉等机械应力及运动诱导的缺氧是运动诱导骨骼肌毛细血管生成的主要原因[3]。上述因素作用于骨骼肌,诱导骨骼肌纤维合成和分泌多种促血管生成因子,如血管内皮生长因子(VEGF)等,促进血管生成[4,5]。
Apelin是1998年发现的G蛋白偶联受体APJ的内源性配体,apelin/APJ广泛分布于体内多个组织和器官,发挥多种生物学效应,包括维持体液稳态、调节摄食、促进细胞增殖及改善机体糖代谢等[6]。近年来,大量临床和实验研究发现apelin具有明显的促进血管生成的作用[7]。在多种动物缺血/缺氧的疾病模型上,外源性给予apelin有显著的保护作用;在apelin敲除的动物,因缺血/缺氧诱导的组织损伤明显加重[8]。有研究发现apelin还是一种肌肉因子[9],在骨骼肌有丰富的apelin及其受体APJ的表达,并通过旁/自分泌的途径作用于骨骼肌自身,其在骨骼肌血管再生中的作用尚未见报道。本研究以高脂饮食喂养诱导的肥胖大鼠为模型,观察10周的跑台运动对肥胖大鼠比目鱼肌毛细血管密度的影响及比目鱼肌apelin/APJ表达的改变,探讨apelin在运动促进骨骼肌毛细血管增生中的作用。
1 材料与方法
1.1 实验动物与材料
本实验所有动物饲养和实验程序按中国《实验动物管理条例》的动物实验标准进行。5周龄雄性SD大鼠由北京大学医学部动物中心提供,实验动物生产许可证号:SCXK(京)2011-0012。动物实验于北京大学医学部动物中心完成,动物使用许可证号:SYXK(京)2011-0039。常规条件下饲养,室内温度为22±2oC,湿度为40%~60%,自由进食和饮水。血脂检测试剂盒由中生北控股份有限公司提供。高脂饲料购自北京科澳协力饲料有限公司,其中每1000 g高脂饲料由800 g基础饲料加200 g脂肪(170 g猪油 +30 g玉米油)与1% 胆固醇混合而成,脂肪供能占40%[10]。Apelin抗体购自Abcam公司(Cambridge,UK),CD31抗体购自武汉博士德生物公司,apelin放免试剂盒购自北京华英生物科技研究所,实验中所用其他试剂均为市售分析纯试剂。
1.2 动物分组与运动模型建立
5周龄健康雄性SD大鼠48只,随机分成对照组(n=16)和高脂喂养组(n=32),分别进行普通饲料和高脂饲料喂养,12周后,以体重≥对照组体重均值加1.4倍标准差为标准[11]筛选肥胖大鼠16只。将对照组大鼠随机分为对照安静组(Con组,n=8)和对照运动组(Con+Exe组,n=8);高脂喂养肥胖大鼠随机分为高脂安静组(HFD组,n=8)和高脂运动组(HFD+Exe组,n= 8)。运动组采用跑台运动,经1周适应,跑台速度逐渐增至26米/分钟[12],运动时间逐渐延长至60分钟。正式跑台训练持续10周,每周5次,每次60分钟。每次训练在下午3点至6点间进行,不使用声、光、电等刺激手段。
1.3 取材、检测
大鼠末次运动结束后,禁食过夜,自由饮水,腹腔注射乌拉坦(1 g/kg)麻醉。经腹主动脉取血,分别收取血浆和血清,血浆采用全自动生化分析仪进行总甘油三酯、总胆固醇、低密度脂蛋白胆固醇和高密度脂蛋白胆固醇水平测定。动物处死后,分离比目鱼肌、腹股沟脂肪(皮下脂肪)、附睾脂肪(内脏脂肪)分别称重,进行下述测定。
1.3.1 免疫组化染色
比目鱼肌置于10%的多聚甲醛溶液中浸泡过夜,然后于20%蔗糖溶液中浸泡过夜,分别进行石蜡包埋和OCT包埋,用于石蜡切片和冰冻切片,切片厚度为5 µm。石蜡切片经脱蜡至水用于血小板内皮细胞粘附分子(Plateletendothelialcelladhesion molecule-1,PECAM-1/CD31)染色,冰冻切片经丙酮固定用于ape⁃lin染色。染色流程如下:磷酸盐缓冲液(PBS)洗3次后,过氧化氢孵育5~10分钟,消除内源性过氧化物酶的活性。蒸馏水冲洗后,PBS浸泡5分钟。经10%山羊血清封闭后冲洗,分别滴加1∶150 CD31或1∶30 apelin一抗、1∶500生物素标记二抗、适量辣根酶或碱性磷酸酶标记的链霉卵白素工作液37oC孵育10~30分钟后,PBS冲洗,显色剂显色。自来水冲洗,复染,脱水,透明,封片,观察结果。随机抽取各组动物比目鱼肌切片,在200倍视野下随机观察5个视野,确定CD31的表达改变。
1.3.2 放免测定法
新鲜分离的比目鱼肌100毫克,置于1.0 mol/L的醋酸溶液中剪碎,100oC水浴10分钟后取出,匀浆后离心取上清按试剂盒步骤进行放免测定。apelin放免试剂盒的标准曲线范围从0~400 ng/mL,相关系数为0.999。所有样品的浓度均在标准曲线的读数范围内。试剂盒检测的最小浓度为100 pg/mL。试剂盒的批内变异系数<7.2%,批间变异系数<10%。
1.3.3 Real-time PCR 测定比目鱼肌apelin、APJ 的mRNA水平
采用Trizol一步法提取骨骼肌总RNA,Promega逆转录系统(Promega,Madison,WI,USA)进行逆转录。real-time PCR反应体积共20 μl:Super Real Pre⁃Mix Plus(SYBR Green)体系(TianGen Biotech,Bei⁃jing)16.8 ml,10mmol/L的上下游引物各0.6 ml,cDNA模板2 ml。GAPDH的上游引物:5’-ACA GCA ACA GGG TGG TGG AC-3’;下游引物为:5’-TTT GAG GGT GCA GCG AAC TT-3’;apelin的上游引物:5’-GGC TAG AAG AAG GCA AC-3’;下游引物为:5’-CTG TGG AGT AAA GAG ACT GG-3’;APJ的上游引物:5’-GGT GCTTGC TGA GTT GCT-3’;下游引物为:5’-TAG ATG GCA GGG ATG AGA-3’。所有引物均由生工生物工程(上海)股份有限公司合成。经95oC 7分钟变性后,进行95oC 30s,60oC 30s,72oC 40s,热循环40次。Real-time PCR于Mx3000P多通道实时定量PCR仪(Stratagene,La Jolla,CA,USA)上进行,以GAPDH作为内参。
1.4 统计方法
2 结果
2.1 高脂饮食及跑台运动对大鼠体重、体脂重和血脂的影响
与正常饮食组大鼠相比,高脂饮食诱导大鼠出现显著肥胖,表现为体重显著增加(P<0.01)(图1),内脏脂肪和皮下脂肪与体重比显著增加(P<0.05),血总胆固醇(P<0.05)、总甘油三酯(P<0.01)、低密度脂蛋白胆固醇(P<0.01)显著升高,高密度脂蛋白胆固醇显著降低(P<0.01)(图2)。10周跑台运动显著逆转了高脂饮食诱导的上述变化,高脂运动组体重显著降低(P<0.01)(图1),体脂比显著下降,脂代谢各参数也都回归到正常水平(图2),提示运动减轻了大鼠肥胖,改善了肥胖大鼠的脂代谢。
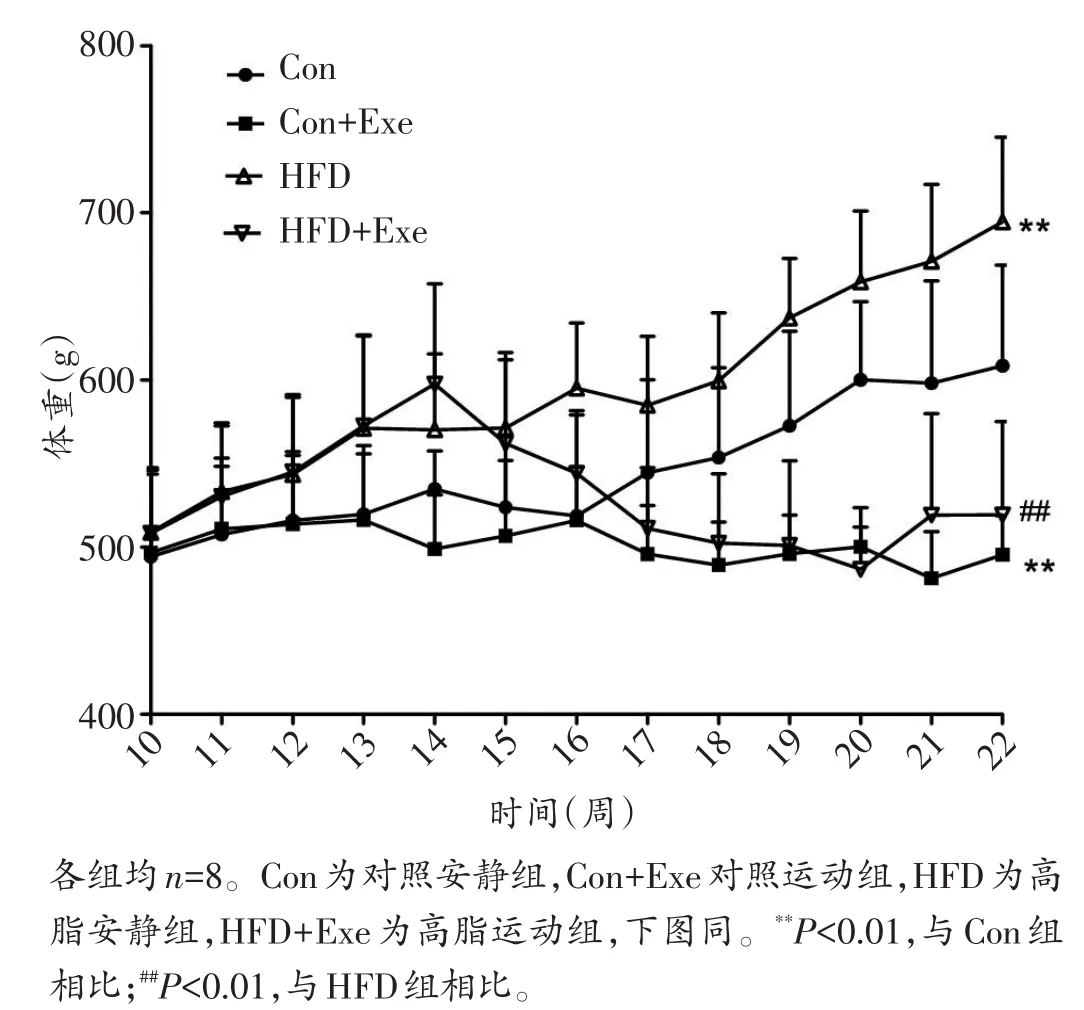
图1 各组大鼠体重变化比较
2.2 高脂饮食及跑台运动对大鼠比目鱼肌毛细血管生成的影响
血小板内皮细胞粘附分子(PECAM-1/CD31)在内皮细胞特异表达,通常作为毛细血管生成的标记蛋白。图3显示,与对照组相比,正常饮食运动组大鼠比目鱼肌CD31水平明显增多,提示毛细血管生成显著增加。而在高脂饮食诱导的肥胖大鼠比目鱼肌CD31的表达与对照组相比无显著变化,提示22周的高脂饮食并未影响比目鱼肌毛细血管的生成。高脂运动组大鼠比目鱼肌CD31的表达与单纯高脂饮食大鼠比显著提高,提示运动对肥胖大鼠骨骼肌毛细血管再生也有促进作用。
2.3 高脂饮食及跑台运动对大鼠比目鱼肌apelin/APJmRNA表达的影响
图4显示,与对照组相比,正常饮食运动组骨骼肌apelin/APJ的表达水平均显著升高(P<0.05),单纯高脂饮食组大鼠比目鱼肌apelin/APJ mRNA表达水平虽有上升,但无显著变化(P>0.05)。跑台训练则显著升高了肥胖大鼠比目鱼肌apelin/APJ mRNA的表达(P<0.05)。结果提示,运动对骨骼肌aplin/APJ的表达有显著的促进作用。
2.4 高脂饮食及跑台运动对大鼠比目鱼肌apelin蛋白表达水平的影响
比目鱼肌中apelin蛋白水平与apelin的mRNA表达结果一致。图5显示,与对照组相比,正常饮食运动组骨骼肌apelin的水平均显著升高(P<0.05),单纯高脂饮食组大鼠比目鱼肌apelin水平无显著变化(P>0.05)。跑台训练则显著升高了肥胖大鼠比目鱼肌ape⁃lin的蛋白表达(P<0.05)。结果提示,运动显著增加骨骼肌apelin的蛋白水平。
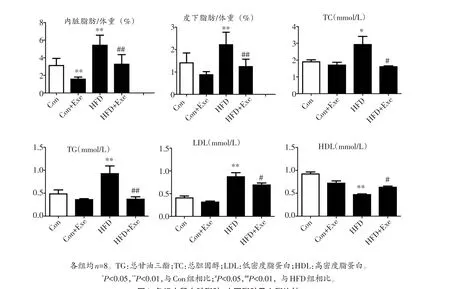
图2 各组大鼠内脏脂肪、皮下脂肪及血脂比较
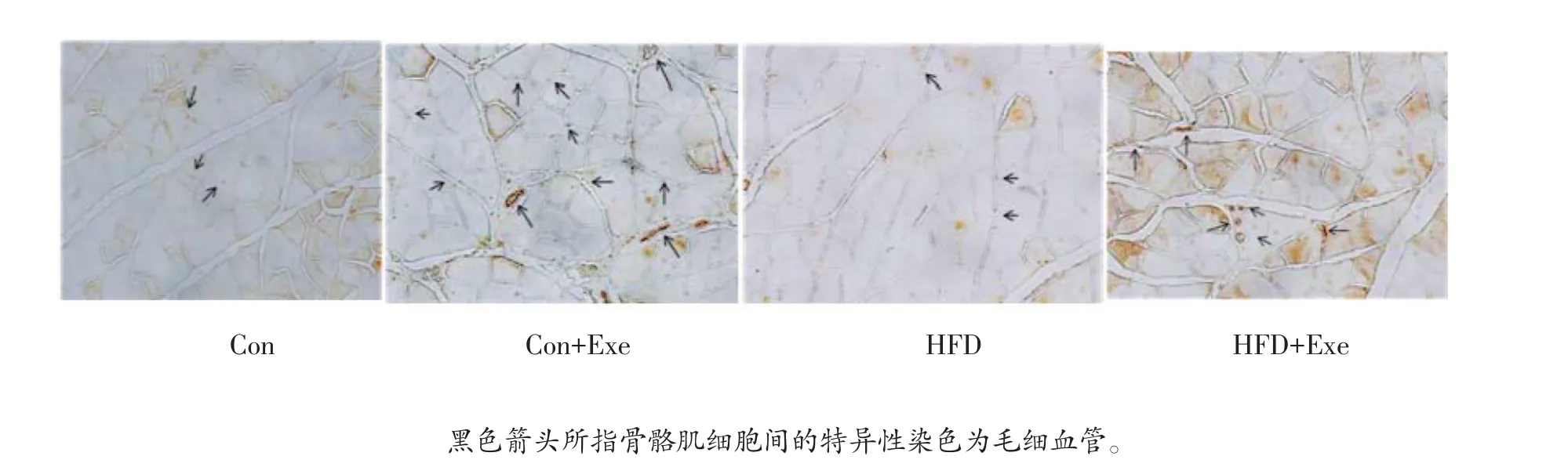
图3 比目鱼肌CD31免疫组化结果(200×)
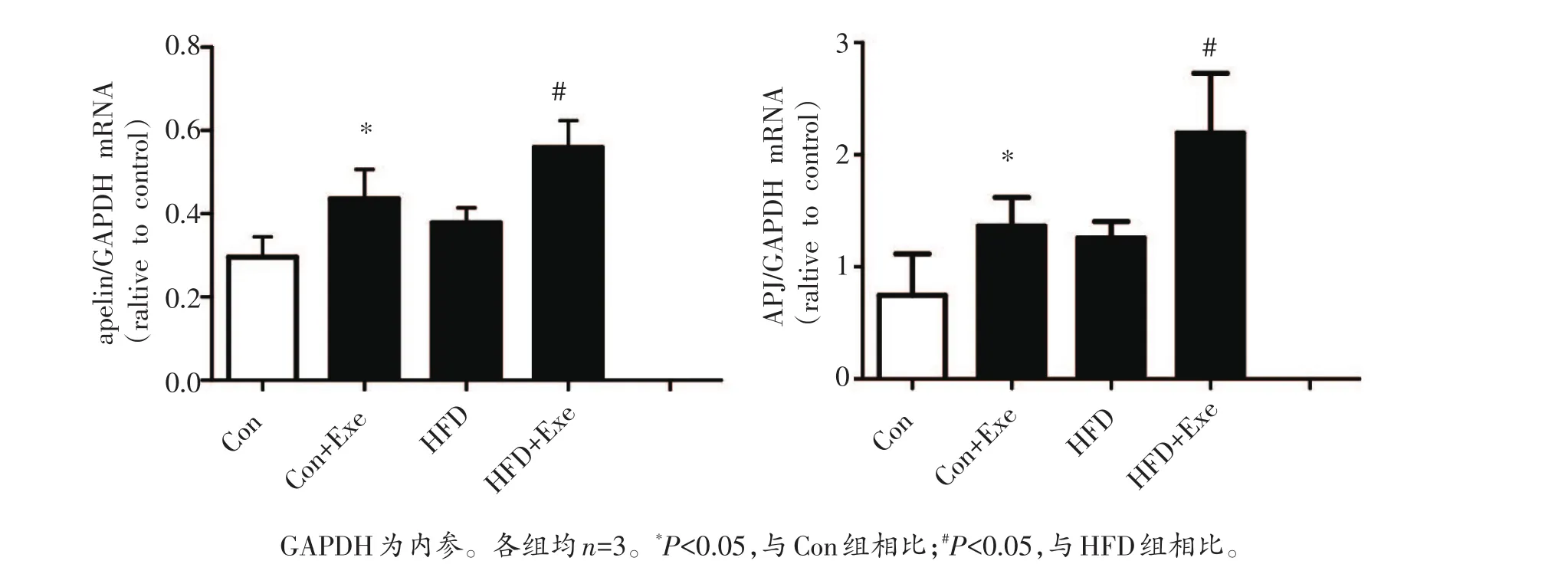
图4 各组大鼠比目鱼肌apelin/APJ mRNA表达比较
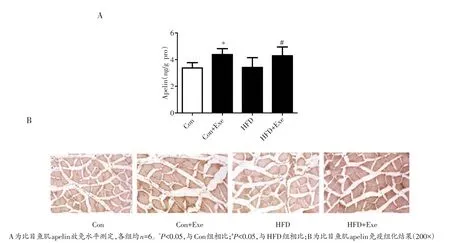
图5 各组大鼠比目鱼肌apelin水平比较
3 讨论
3.1 肥胖和运动对骨骼肌毛细血管生成的影响
毛细血管是直径小于5 μm的微血管,毛细血管重占骨骼肌重量的2%~3%[3]。在骨骼肌,毛细血管的走向与骨骼肌纤维平行,在毛细血管间有短小的分支相连,形成毛细血管网包裹在肌纤维四周。毛细血管管壁很薄,在0.2 μm左右[13],非常有利于组织对氧气和营养物质的吸收及代谢废物的排出。毛细血管与肌纤维的比例直接关系到骨骼肌运动能力及代谢功能。
肥胖时骨骼肌毛细血管密度改变,但在不同的肥胖模型,骨骼肌毛细血管密度的改变也不同。在ZDF(zucker diabetic fatty rats)肥胖及糖尿病大鼠,骨骼肌毛细血管标记物PECAM-1/CD31的表达显著下降[14],斜方肌毛细血管网分支下降37%,纵向毛细血管密度降低19%,血流量下降了近44%[15],提示骨骼肌毛细血管密度显著降低。但是,在高脂饮食喂养诱导的肥胖模型,3周[16]、8周[14]、12周[17]的高脂膳食喂养均未明显影响骨骼肌CD31的表达,提示高脂饮食喂养不影响骨骼肌毛细血管生成。在本研究中,40%高脂膳食喂养22周,大鼠比目鱼肌CD31的表达也没有出现显著变化,与以往研究结果相同。
运动对骨骼肌毛细血管生成有强大的促进作用,在本实验中,我们发现不管是正常饮食组还是高脂饮食组,运动都显著促进比目鱼肌CD31的表达,提示运动促进了骨骼肌毛细血管生成。运动时机械刺激和局部缺氧诱导骨骼肌细胞缺氧诱导因子(hypoxia induc⁃ ible factors,HIFs)激活[18],HIFs调节多种参与血管生成的靶基因的转录和转录后调节,如血管内皮生长因子A(vascular endothelial growth factor A,VEGFA)等,进而促进血管生成[19]。目前研究发现,apelin是HIF1α的下游靶基因之一[20],其在血管生成中的作用备受关注。
3.2 apelin/APJ的促血管生成作用
在多种以缺氧为特征的疾病,如缺氧性肺动脉高压、缺血性心力衰竭、缺血性脑损伤、急性肾损伤、早产儿视网膜疾病及肥胖时,均发现内源性的apelin表达升高,提示apelin参与了缺氧性疾病的发展[8],其病理生理学作用如何?在动物缺氧性肺动脉高压模型,外源性给予apelin均显著改善疾病的发展,而apelin敲除小鼠,肺动脉高压的症状加重[21];此外在缺血心衰模型等其他缺氧性疾病模型观察到相同的变化[22],提示apelin对缺血性疾病有显著的保护作用。进一步研究提示,apelin通过促进内皮型一氧化氮合酶的表达,激活PI3K/Akt通路,促进细胞存活、增殖和迁移,上调血管生长因子的表达等途径,参与病理性或生理性的血管生成[8]。
Yamamoto等发现在高脂饮食喂养的apelin转基因小鼠,骨骼肌中促血管新生蛋白因子(angiopoietin-1,Ang 1)及其受体内皮特异性受体酪氨酸激酶2(endo⁃thelium-specific receptor tyrosine kinase 2,Tie2)的表达显著增强,同时骨骼肌中CD31的表达增强,毛细血管与肌纤维比值显著增加,提示apelin有强大的促进骨骼肌毛细血管生成的作用[23]。骨骼肌能合成和分泌apelin,其以旁/自分泌的方式,作用于骨骼肌局部,发挥生物学效应[9]。在本研究中我们发现运动显著上调比目鱼肌apelin/APJ的表达,其表达与比目鱼肌毛细血管密度改变相一致,提示apelin可能参与了运动诱导的骨骼肌毛细血管生成。
3.3 肥胖和运动对骨骼肌apelin/APJ表达的调节
多数研究报道,肥胖时循环中apelin水平显著升高,而运动或减脂手术均显著下调血浆apelin水平[9,24],血浆中apelin水平与肥胖呈正相关[25]。apelin在机体的多种组织表达,肥胖和运动对apelin表达的调节具有显著的组织特异性。研究证实,循环中apelin的主要来源为脂肪组织,骨骼肌合成和分泌的apelin主要以旁分泌的方式作用于骨骼肌自身。
Besse-Patin等[9]研究发现,8周的耐力运动显著增加了11名男性肥胖受试者骨骼肌中apelin的mRNA表达,并提示其变化与运动时细胞内钙离子、环腺苷酸等第二信使的改变有关。在本研究中,高脂饮食对骨骼肌apelin的表达无显著影响,运动则显著诱导了正常饮食组和高脂饮食组大鼠骨骼肌apelin的mRNA、蛋白表达,提示运动能特异性地上调骨骼肌apelin的表达,发挥其旁/自分泌作用。
4 总结
10周的跑台运动显著增加体重正常和肥胖大鼠比目鱼肌毛细血管生成,显著上调骨骼肌apelin的mRNA、蛋白表达,提示apelin可能参与了运动诱导的骨骼肌毛细血管生成。
[1] Olfert IM,Baum O,Hellsten Y,et al.Advances and challenges in skeletal muscle angiogenesis.Am J Physiol Heart Circ Physiol,2016,310(3):H326-336.
[2] 张靓,刘小园,唐朝枢.肥胖与骨骼肌重塑.生理科学进展,2012,43(6):405-410.
[3] Haas TL,Nwadozi E.Regulation of skeletal muscle cap⁃illary growth in exercise and disease.Appl Physiol Nutr Metab,2015,40(12):1221-1232.
[4] Gustafsson T.Vascularremodelling in human skeletal muscle.Biochem Soc Trans,2011,39(6):1628-1632.
[5] Gorman JL,Liu STK,Slopack D,et al.Angiotensin II evokes angiogenic signals within skeletal muscle through co-ordinated effects on skeletal myocytes and endothelial cells.PloS ONE,2014,9(1):e85537.
[6] Hu H,He L,Li L,et al.Apelin/APJ system as a ther⁃apeutic target in diabetes and its complications.Mol Gen⁃et Metab,2016,119(1-2):20-27.
[7] Yang X,Zhu W,Zhang P,et al.Apelin-13 stimulates angiogenesis by promoting crosstalk between AMP-activat⁃ed protein kinase and Akt signaling in myocardial micro⁃vascularendothelialcells.MolMed Rep,2014,9(5): 1590-1596.
[8] He L,Xu J,Chen L,et al.Apelin/APJ signaling in hy⁃poxia-related diseases.Clin Chim Acta,2015,451(Pt B): 191-198.
[9] Besse-Patin A,Montastier E,Vinel C,et al.Effect of endurance training on skeletal muscle myokine expres⁃sion in obese men:identification of apelin as a novel myokine.Int J Obes(Lond),2014,38(5):707-713.
[10]Kim YJ,Park T.Genes are differentially expressed in the epididymal fat of rats rendered obese by a high-fat diet.Nutr Res,2008,28(6):414-422.
[11]孙长颢,刘荣,李颖,等.饮食诱导肥胖抵抗大鼠激素敏感脂肪酶基因的表达.中华预防医学杂志,2004,38(5): 339-341.
[12]Gauthier MS,Couturier K,Charbonneau A,et al.Ef⁃fects of introducing physical training in the course of a 16-weekhigh-fatdietregimen on hepaticsteatosis,adipose tissue fat accumulation,and plasma lipid profile.Int J ObesRelatMetabDisord,2004,28(8):1064-1071.
[13]Simionescu M,Simionescu N,Palade GE.Morphometric data on the endothelium of blood capillaries.J Cell Biol,1974,60(1):128-152.
[14]Roudier E,Chapados N,Decary S et al.Angiomotin p80/ p130 ratio:a new indicator of exercise-induced angiogen⁃ic activity in skeletalmusclesfrom obese and nonobese rats?J Physiol,2009,587(Pt 16):4105-4119.
[15]Benedict KF,Coffin GS,Barrett EJ,et al.Hemodynam⁃ic systems analysis of capillary network remodeling dur⁃ing the progression of type 2 diabetes.Microcirculation,2011,18(1):63-73.
[16]Thomas MM,Trajcevski KE,Coleman SK,et al.Early oxidative shifts in mouse skeletalmuscle morphology with high-fat diet consumption do not lead to functional improvements.Physiol Rep,2014,2(9):e12149.
[17]Li Y,Hazarika S,Xie D,et al.In mice with type 2 diabetes,a vascular endothelial growth factor(VEGF)-activating transcription factor modulates VEGF signaling and induces therapeutic angiogenesis after hindlimb isch⁃emia,Diabetes.2007,56(3):656-665.
[18]Lindholm ME,Rundqvist H.Skeletal muscle hypoxia-in⁃ducible factor-1 and exercise.Exp Physiol,2016,101(1):28-32.
[19]Zimna A,Kurpisz M.Hypoxia-Inducible Factor-1 in Physiological and Pathophysiological Angiogenesis:Appli⁃cations and Therapies.Biomed Res Int,2015,2015: 549412.
[20]M Eyries,G Siegfried,M Ciumas,et al.Hypoxia-in⁃duced apelin expression regulates endothelial cell prolifer⁃ation and regenerative angiogenesis.Circ Res,2008,103(4):432-440.
[21]SM Chandra,H Razavi,J Kim,et al.Disruption of the apelin-APJ system worsens hypoxia-induced pulmo⁃nary hypertension. Arterioscler Thromb Vasc Biol,2011,31(4):814-820.
[22]XJ Zeng,LK Zhang,HX Wang,et al.Apelin protects heartagainstischemia/reperfusion injury in rat.Pep⁃tides,2009,30(6):1144-1152.
[23]Yamamoto T,Habata Y,Matsumoto Y,et al.Apelintransgenic mice exhibit a resistance against diet-induced obesity by increasing vascularmassand mitochondrial biogenesis in skeletal muscle. Biochim Biophys Acta,2011,1810(9):853-862.
[24]Sheibani S,Hanachi P,Refahiat MA.Effect of Aerobic Exercise on Serum Concentration of Apelin,TNFα and Insulin in Obese Women.Iran J Basic Med Sci,2012,15(6):1196-1201.
[25]Liao YC,Chou WW,Li YN,et al.Apelin gene poly⁃morphism influences apelin expression and obesity pheno⁃types in Chinese women.Am J Clin Nutr,2011,94(3):921-928.
Effects of Treadmill Running on Capillary Density and Apelin Expression in Soleus Muscle of High-fat Diet Rats
Zhang Jing,Ma Jin,Chen Xuefei,Dong Zeyuan School of Physical Education and Sports Science,Beijing Normal University,Beijing 100875,China Corresponding Author:Zhang Jing,Email:zhangjing@bnu.edu.cn
ObjectiveTo observe the alteration of capillary density and apelin/APJ expression in sole⁃us muscles of high-fat diet rats.MethodsMale 5-week-old Sprague-Dawley rats were randomly divid⁃ed into a control group and a high-fat diet group.After 12 weeks of high-fat diet,16 rats were se⁃lected and randomly divided into a sedentary group and a treadmill running group.The exercise rats underwent 60-minute treadmill running at 26 m/min 5 days a week for 10 weeks.The body weight,body fat and blood lipid level were measured for all rats.The protein expression of Soleus CD31 and apelin was determined using immunohistochemical staining,soleus apelin content was determined using the radioimmunoassay,and the mRNA expression of apelin/APJ was detected using real-time PCR.Re⁃sultsCompared with the control group,significant increase was observed in the body weight,body fat and the level of total triglyceride,total cholesterol and low density lipoprotein cholesterin,but signifi⁃cant decrease was found in the high density lipoprotein cholesterin in the high-fat diet group.There were no significant differences in the capillary density and mRNA levels of apelin/APJ between the two groups.Compared with the sedentary high-fat diet group,significant improvement was observed in the body weight and blood lipid level of the treadmill running group.Moreover,significant increase was also observed in the capillary density,the expression of apelin/APJ mRNA,as well as that of ape⁃lin protein in the treadmill running group(P<0.05).ConclusionThe treadmill running can significant⁃ly increase capillary density of obese rats,as it may activate the expression of Apelin/APJ.
treadmill running,capillary density,apelin,obesity,soleus muscle
2017.01.05
国家自然科学基金(31471137)
张靓,Email:zhangjing@bnu.edu.cn
猜你喜欢
中国典型病例大全(2022年7期)2022-04-22 02:19:54
中华养生保健(2020年5期)2020-11-16 01:44:36
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
医学研究杂志(2015年12期)2015-06-10 06:57:46
医学研究杂志(2015年5期)2015-06-10 06:43:26
中华皮肤科杂志(2014年3期)2014-12-19 12:54:56
中国火炬(2014年8期)2014-07-24 14:30:21
食品工业科技(2014年5期)2014-03-11 18:14:30
