
镍对蕹菜生长和营养成分的影响
2017-06-07 10:30:12王金生王业耀王学礼秦旭芝
广东农业科学 2017年3期
黎 宁,王金生,王业耀,3,王学礼,秦旭芝
(1.北京师范大学水科学研究院,北京 100875;2.广西壮族自治区环境监测中心站,广西 南宁 530028;3.中国环境监测总站,北京 100012;4.广西大学农学院,广西 南宁 530004)
镍对蕹菜生长和营养成分的影响
黎 宁1,2,王金生1,王业耀1,3,王学礼4,秦旭芝2
(1.北京师范大学水科学研究院,北京 100875;2.广西壮族自治区环境监测中心站,广西 南宁 530028;3.中国环境监测总站,北京 100012;4.广西大学农学院,广西 南宁 530004)
通过水培方式种植蕹菜,研究以尿素态氮作唯一氮源条件下,不同浓度镍对蕹菜生长、脲酶活力和某些营养成分的影响。结果表明:适量镍对蕹菜生长、脲酶活力和氨基酸合成有促进作用,但过量的镍产生毒害作用。施镍20 μg/L处理的蕹菜鲜重最高、比对照高12.1%,但镍浓度高于320 μg/L时蕹菜鲜重下降,施镍1 280 μg/L处理的蕹菜表现出受害症状;适量的镍(20~80 μg/L)可显著提高蕹菜叶片的脲酶活力,最高幅度达13.6%,脲酶含量也显著提高,最大增幅19.0%,但过量镍(大于640 μg/L)会抑制脲酶活力。研究足量镍(60 μg/L)影响蕹菜脲酶的作用机理,结果显示,镍可能先是提高脲酶比活力,然后参与脲酶合成而提高脲酶含量,从而更进一步提高脲酶活力。叶片中总氨基酸含量在镍0~40 μg/L范围内随镍浓度提高而升高,最高升幅44.7%,镍大于10 μg/L时效果极显著,之后保持稳定水平;叶片中可溶性糖含量在过高浓度镍处理(大于640 μg/L)时显著升高。
水培;蕹菜;施镍处理;脲酶;氨基酸;可溶性糖
镍在高浓度情况下会对生物产生毒害作用,通常被视为污染物,出于对环境保护的关注,近年来学者对镍的研究多集中在其污染、毒害作用方面,对于镍起到的积极作用研究相对较少。实际上,镍是人体必需的微量元素,也是植物体内脲酶和微生物某些酶(如氢化酶、甲基辅酶和一氧化碳脱氢酶)的组分,参与生理生化过程,一些植物在缺镍情况下难以完成生长周期,因此科学家们认为,镍是高等植物生长所必需的营养元素之一[1-3]。研究显示,缺镍情况下,水稻对尿素的利用率和脲酶活力低下,在以尿素态氮为氮源时出现缺素症状[4]。近年来,镍促进植物生长的研究不时见于报道[5],国外有学者认为目前镍在高等植物中仅限于参与脲酶的作用,对诸如自身免疫力等其他方面的作用仍知之甚少,且对镍和脲酶之间关系研究了解清楚的还仅限于大豆、马铃薯和拟南芥等少数植物,建议拓展到更多植物品种[6]。鉴于此,本试验选取蕹菜的一个品种白籽白梗为材料,研究镍对植物生长和脲酶的影响和作用机理。蕹菜(Ipomoea aquatica Forsk.)也称空心菜、通心菜、蓊菜等,喜高温多湿环境,在水源、阳光、肥料充足的土壤中生长茂盛,是我国各地尤其是南方农村普遍栽培的重要蔬菜,其栽培方式分旱栽和水植两种,北方以旱栽为主,南方旱栽与水植并存。蕹菜品种有很多,本试验选取的蕹菜品种白籽白梗经筛选试验表现对镍敏感。采用水培方法对蕹菜进行培育,观测镍对蕹菜植株生长的影响,分析脲酶活力以及可溶性糖、总氨基酸含量等指标,研究结果将进一步丰富镍元素影响植物生理生化过程的科学数据,也为合理施用微肥、提高蔬菜产量和品质、保护农业环境提供依据。
1 材料与方法
1.1 试验材料
供试蕹菜品种为白籽白梗,是经过筛选的对镍敏感品种,经检测,种子含镍量小于1 mg/kg。试验用镍源为NiSO4(AR),水培用去离子水的电导率低于0.5 μS/cm,配制水培营养液的化学试剂均为分析纯。
1.2 试验方法
试验在广西大学农学院玻璃网室布置水培试验。3月22日发芽,3月31日移栽,4月13日换营养液时进行镍处理,4月18日测定各项目。水培营养液配方参照华南农业大学的蕹菜专用配方[7],以尿素作唯一氮源,设含镍量分别为0(CK)、5、10、20、40、80、160、320、640、1 280 μg/L等10个处理,每个处理3次重复,共30盆(盆的规格为内径20 cm、深15 cm的椭圆收窄的塑料花盆,营养液装进聚乙烯塑料袋后放置在花盆中),每盆3.5 L营养液。
在单独准备的石英砂盘中常温下育苗至2~3叶时,取均匀健壮的蕹菜苗,根部裹着海绵后固定在有很多开孔(1 cm直径)的泡沫板上,每孔1株,一共23株,放置在已经灌满营养液的塑料盆中,蕹菜苗的根伸入营养液中,泡沫板用重物压着防止大风刮倒,置于露天玻璃棚屋中。每周更换营养液,移栽后7 d内营养液浓度取蕹菜专用配方的1/4浓度值,第8~14 d则取1/2浓度值,之后营养液浓度维持不变直至收获。移栽后第14 d更换营养液时进行施镍处理,施镍处理5 d后采集倒第4、5片成熟叶进行分析,测定脲酶活力、脲酶含量、脲酶比活力、总氨基酸和可溶性糖含量等,同时观察施镍后蕹菜的生长变化,包括植株叶色、长势等,移栽后第35 d采收,测定每盆蕹菜的地上、地下部的鲜重和干重、根冠比等指标。
同时设计另外一个水培试验,研究镍对蕹菜脲酶的作用机理。设计5组成对处理,每天布置一组处理,每组处理2盆,其中一盆不施镍(CK),另一盆施镍浓度60 μg/L。具体操作为:将蕹菜种子分成5组,每组60颗,自3月22日起,每天发芽一组,发芽后第5 d选择2叶苗移栽至带孔泡沫板上固定,置于两个容积1 L的塑料盆中水培种植,每盆16株,置于25℃带人工光照恒温恒湿条件下培养,每天更换营养液,培养6 d后进行施镍处理,第11 d后得到已经施镍处理1、2、3、4、5 d的蕹菜以及未施镍的5个对照,4月13日同一时间采集这10盆蕹菜叶片分别分析脲酶活力、脲酶含量、脲酶比活力等指标。
1.3 测定项目及方法
脲酶活力采用苯酚钠-次氯酸钠比色法[8-9]测定,以30℃下每克鲜叶每小时催化尿素产生的氨气(μmol)表示酶活力,单位为μmol/g·h。脲酶含量以每克蕹菜鲜叶中含脲酶这种蛋白质的量表示,采用考马斯亮蓝法[10]测定鲜叶脲酶提取液中的蛋白质含量作为脲酶含量指标,单位为mg/g。脲酶比活力即单位酶量的催化能力,以30℃下每毫克脲酶每小时产生的氨气(μmol)表示,单位为 μmol/mg·h。可溶性糖含量采用蒽酮比色法[12]测定,单位为%。总氨基酸含量采用茚三酮显色法[13]测定,单位为g/100g。
2 结果与分析

表1 不同浓度镍处理对蕹菜生长的影响
2.1 不同浓度镍处理对蕹菜生长的影响
水培蕹菜施镍试验结果(表1)显示,以尿素为氮源时,不同浓度镍处理的蕹菜总鲜重、地上部和地下部鲜重差异达到显著水平,低浓度镍处理对蕹菜的生长有促进作用,20 μg/L镍处理的蕹菜地上部鲜重最高、比对照高12.1%,其次是10 μg/L镍处理、比对照高10.7%,但5 μg/L镍处理对蕹菜的生长无明显影响。当镍处理浓度高于320 μg/L时,蕹菜鲜重开始下降,说明此浓度下蕹菜生长发育受到抑制,表现为生物量下降;镍处理浓度为640 μg/L时,蕹菜鲜重与最适浓度处理(10~20 μg/L)的植株鲜重相比差异显著;当镍处理浓度为1 280 μg/L时,植株鲜重显著低于对照。不同浓度镍处理蕹菜地上部干重的变化趋势与鲜重基本一致,根鲜重和鲜重根冠比也有类似规律、但不明显,根干重的规律性也不明显,干重根冠比则表现为对照最高。从以上几个指标的测定结果看,蕹菜地上部鲜重与镍浓度关系密切,说明施镍主要影响了蕹菜地上部的生长。观察还发现,镍1 280 μg/L处理蕹菜1周后出现明显受害症状,表现为叶片脉间失绿黄化、侧根粗大。
2.2 不同浓度镍处理对蕹菜叶片脲酶活力的影响
采集倒第4、第5片成熟叶片,测定脲酶活力和脲酶含量,并计算脲酶比活力。结果(图1)显示,经镍处理后蕹菜脲酶活力发生明显变化,其中20、40、80 μg/L镍处理极显著提高了脲酶活力。从变化趋势看,在镍处理浓度0~80 μg/L范围内,脲酶活力随镍浓度的提高而提高,从2.95 μmol/g·h增加到3.35 μmol/g·h,增幅13.6%,但在低浓度(≤10 μg/L)时增幅不明显,过量的镍(>80 μg/L)处理使脲酶活力下降至低于或接近对照水平。图2表明,蕹菜叶片中脲酶含量在低浓度镍范围内(≤80 μg/L)随镍浓度提高而逐步升高,从1.68 mg/g增加到2.00 mg/g,最大增幅19.0%,极显著高于对照和低浓度镍处理,在镍处理浓度大于80 μg/L时,脲酶含量开始下降,但仍一直高于对照水平。对脲酶比活力而言,只有20 μg/L和40 μg/L两个镍处理的脲酶比活力高于对照,

图1 蕹菜叶片脲酶活力随镍处理浓度的变化
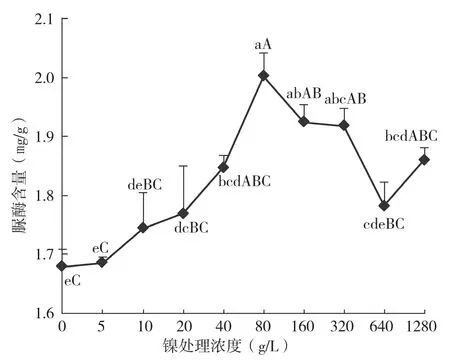
图2 蕹菜叶片脲酶含量随镍处理浓度的变化
其中20 μg/L镍处理的脲酶比活力最高为1.83 μmol/mg·h,比对照(1.75 μmol/mg·h)增加4.6%,且在镍处理浓度大于20 μg/L时比活力呈下降趋势,当镍处理浓度≥160 μg/L时,脲酶比活力极显著低于对照(图3)。

图3 蕹菜叶片脲酶比活力随镍处理浓度的变化
从脲酶3个指标的最大增幅看,镍处理对蕹菜脲酶含量的提高作用最明显,其次是脲酶活力,而对脲酶比活力的提升效果最弱。综合分析3个指标和蕹菜生长情况与镍浓度的关系可以发现,当镍浓度为5 μg/L时,脲酶活力、含量和比活力变化不明显,此时蕹菜生长量也无变化(表1);当镍浓度提高到10 μg/L时,脲酶含量首先提高,脲酶活力暂无变化,脲酶比活力表现为下降,蕹菜鲜重有所增加;随着镍浓度增加到20 μg/L,脲酶活力增幅较大,且高于脲酶含量增幅,导致脲酶比活力此时有一个最大增幅,可能此时脲酶被激活,此时蕹菜鲜重最高;随着镍处理浓度继续提高至80 μg/L,脲酶活力提高趋缓,但脲酶含量提高更快,比活力表现出逐步下降,直到镍浓度160 μg/L时3个脲酶指标都下降,说明80 μg/L是脲酶变化的拐点,镍浓度高于此值时蕹菜脲酶活力开始受到抑制。
根据镍浓度大于80 μg/L后脲酶活力可能受抑制的试验结果,采用稍低浓度(60 μg/ L)镍处理水培蕹菜,研究脲酶活力在施镍后随时间的变化。结果(图4~图6)显示,经镍处理后,叶片脲酶活力随时间的延长而升高,从第1 d的2.89 μmol/g·h升高到第5 d的4.01 μmol/g·h,增幅38.8%;相比较而言,脲酶含量到第3 d才有所提高,第5 d升高至2.23 mg/g,增幅16.1%;脲酶比活力在镍处理后2 d有所上升,从1.51 μmol/mg·h提高至1.74 μmol/mg·h,增幅15.2%,随后升幅趋缓,到第5天提高至1.80 μmol/mg·h,增幅20%;不施镍对照处理的3个脲酶指标在生长过程中无太大变化。这说明适量镍处理既能提高脲酶活力,又能提高脲酶含量,在2 d内主要提高脲酶活力,此时脲酶含量暂未增加,从而表现为脲酶比活力提高,且脲酶比活力在提高后就基本保持稳定;随时间推移,脲酶含量逐步增加。据此,本研究推测镍的作用机理如下:适量镍在短期内(2 d)发挥了对脲酶的激活作用,提高了脲酶比活力并保持在一个稳定水平,随着试验时间延长,在比活力不再提高的情况下,镍通过参与脲酶合成,使脲酶含量继续提高,故施镍5 d后脲酶活力提高值是脲酶比活力和脲酶含量两方面提升的结果。

图4 镍处理蕹菜叶片脲酶活力随时间变化

图5 镍处理蕹菜叶片的脲酶含量随时间变化

图6 镍处理蕹菜叶片脲酶比活力随时间变化
2.3 不同浓度镍处理对蕹菜叶片氨基酸和可溶性糖含量的影响
通过分析氨基酸和可溶性糖含量,可了解镍对蕹菜碳氮代谢的影响情况。从图7、图8可以看出,蕹菜叶片中氨基酸含量在镍浓度≤40 μg/L时随镍浓度提高而快速提高,从每100 g含0.73 g升高至1.06 g,增幅高达44.7%。当镍浓度大于40 μg/L时,每100 g蕹菜叶片氨基酸含量维持在1.0 g左右。当镍浓度≤320 μg/L时,可溶性糖含量一直维持在0.32%左右,直至镍处理浓度为640 μg/L时可溶性糖含量才急速升高。据分析,这两个营养成分的变化趋势与其在植物代谢过程中所起的作用有关,适量浓度的镍处理提高了蕹菜脲酶活力和含量,脲酶转化尿素产生铵,参与植物对氮的同化过程,提高了植物对尿素的利用速度,体现为叶片中的氨基酸含量提高,说明镍通过提高脲酶转化尿素效率而参与到氮代谢过程中,这与刘国栋[14]的结果一致。
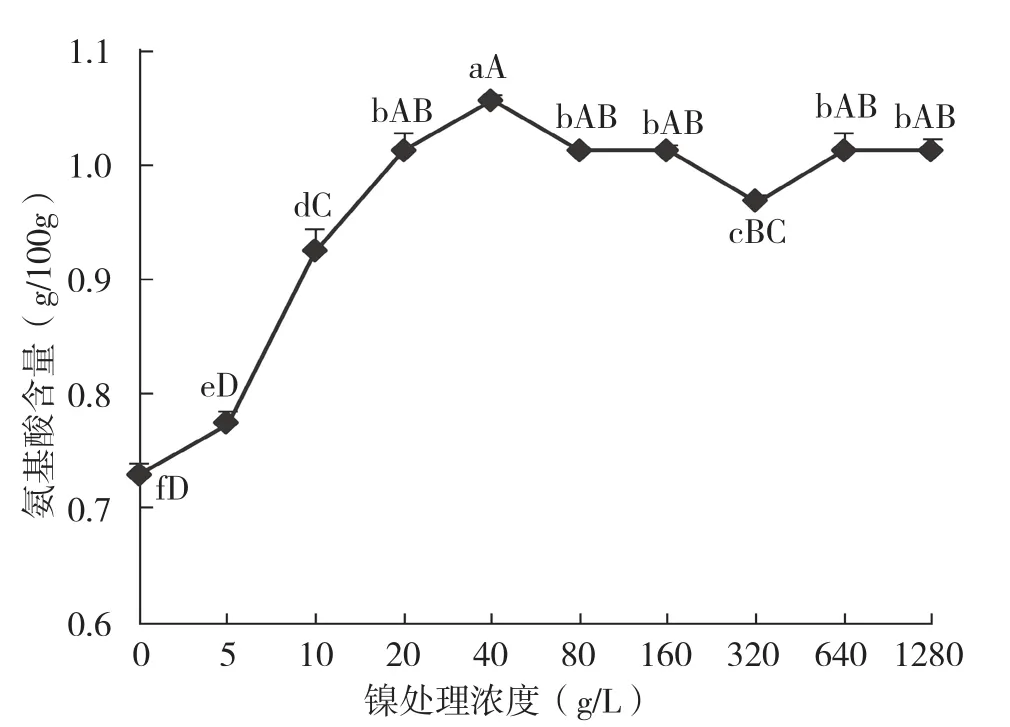
图7 蕹菜叶片氨基酸含量随镍处理浓度的变化

图8 蕹菜叶片可溶性糖含量随镍处理浓度的变化
3 结论与讨论
3.1 镍促进蕹菜生长的最佳浓度
镍是植物和许多微生物的必需元素,缺镍会影响植物生长[15],低浓度的镍能促进植物生长[5],过量的镍对植物生长有毒害作用,影响养分吸收、营养平衡、叶绿素合成等,甚至土壤微小动物也会受到影响。学者们从不同角度研究了镍的作用,如低剂量镍能促进植物根生长,高浓度镍抑制并伤害蚯蚓[16];少量镍能促进小麦生长,但超出临界值则长势下降[17];低浓度镍促进植物细胞分裂,而浓度过高对细胞有毒害[18],过量的镍使大豆叶片叶绿素含量降低[19]。本试验结果表明,在水培条件下,以尿素为氮源时,20 μg/L镍处理可满足蕹菜白子白梗品种的生长需要,地上部鲜重最高;当镍处理浓度高于320 μg/L时,蕹菜生长发育受到抑制,表现为蕹菜鲜重下降,镍处理浓度再继续提高则使蕹菜出现受害症状,这与刘高琼[20]、黄锦孙[21]、武振中[22]和王兵[20-23]的研究结果相似。刘高琼试验结果显示,在以尿素为氮源的水培条件下,镍促进蕹菜生长的最佳浓度为上海矮抗青500 μg/L、南农矮脚黄1 000 μg/L[20]。这与本研究得到的促进蕹菜生长的最佳镍浓度20 μg/L相比差别很大,这说明蕹菜不同品种间对镍的需求有所不同,对白子白梗蕹菜品种而言,镍最佳刺激浓度为20 μg/L,该浓度即可满足蕹菜的生长需要,蕹菜鲜重可提高12.1%,脲酶比活力最高,脲酶活力增速最大。因此,实际农业生产中若需施用镍肥,必须考虑作物品种的需镍特性。
3.2 镍影响蕹菜脲酶活力的内在机理
Loe等[6]认为脲酶是高等植物中唯一把镍作为组成分的酶,即尚未发现镍是高等植物中其他酶的组分,但在微生物体内却是很多酶的组成分。对于高等植物,目前比较明确的是镍作为脲酶的组分参与到植物生理生化过程,其主要作用是激活脲酶而使尿素态氮水解变成铵态氮,进而参与到氨基酸、蛋白质和其他含氮化合物的合成,但也有研究显示,适量镍可增强苹果叶片谷氨酰胺合成酶(GS)和超氧化物歧化酶(SOD)活性,延缓叶片衰老[24],也有报道认为镍能体外激活乙二醛酶[25]。适量镍可以提高脲酶活性的相关报道多见于需镍较多的豆科作物、禾本科的水稻等[4,21,26-27],镍影响根际土壤中脲酶的报道也有不少[28-29]。然而,镍到底是通过提高脲酶含量还是脲酶比活力来提高脲酶活力,其机理如何,很少有详细分析。岛田典司等[27]研究了水稻叶片脲酶总活力随处理时间的变化趋势,发现脲酶总活力在48 h内迅速上升,1周后趋势减缓,但到底是脲酶含量还是比活力所导致的变化,未见明确解释。
本研究设计了两个试验,一个是不同浓度镍处理对脲酶的作用,另一个是在足够镍量处理下,脲酶活力随时间的变化。前者结果显示,在镍浓度≤5 μg/L时,3个脲酶活力指标还没有变化;当镍浓度为10 μg/L时,脲酶活力没有变化,但脲酶含量有所增加,相应的比活力下降;当镍浓度提高到20 μg/L时,3个指标值均有提高,其中脲酶活力有较大变化,比活力有所提高,脲酶含量有一定增加;随着镍处理浓度的继续提升,脲酶含量继续提高,但脲酶活力增幅不大,相应的比活力开始下降。说明外源镍太少对脲酶无影响,需要达到一定的量才起作用,对蕹菜来说存在一个最低激活浓度值,约为10 μg/L。从脲酶活力随时间的变化结果看,镍处理在2 d内主要作用是提高脲酶比活力,之后基本保持稳定,此时镍起到激活脲酶的作用,4~5d后,脲酶含量才大幅提高,此时镍的作用应该是参与并加快了脲酶合成。综上分析,本研究推测镍对脲酶的作用机理如下:适量镍进入植物体内后首先激发脲酶活力,体现在提高脲酶比活力,但并不随镍浓度的继续提高而提高,而是保持一个稳态;之后镍作为脲酶的组分参与脲酶的合成,使脲酶含量继续升高,使脲酶总活力进一步提升。因植物生理生化活动是个复杂过程,本研究推测的镍作用机理还需更多实验来加以证实。
3.3 镍对相关营养成分的影响
植物对氮的吸收形式要经过铵态氮形态,镍激活脲酶而使尿素水解变成NH4+,再与植物体内的有机物结合成氨基酸、蛋白质和其他含氮化合物。可溶性糖是光合同化物与能量的运输和贮藏形式,为植物的生长发育提供能量和代谢中间产物。有研究表明,镍过量会影响植物对其他元素(如铁元素)的吸收,影响叶绿素合成[30]或抑制蛋白质合成[31]。在尿素作为唯一氮源的情况下,外源镍激活了植物体内脲酶活力,并提高了脲酶含量,使尿素的水解利用速度加快,这较好地解释了本研究观察到的叶片总氨基酸随镍处理浓度提高而迅速提高的现象,至于可溶性糖含量在合适镍处理浓度范围基本保持稳定,应该是植物生长发育需要碳水化合物而使可溶性糖被迅速利用而不会积累。
高浓度镍处理时蕹菜叶片中可溶性糖含量升高的原因较为复杂,可能是由于高浓度镍对植物生长造成损害,使植物无法利用光合作用的产物,造成蕹菜叶片中可溶性糖累积,也可能是重金属胁迫造成植物体内可溶性糖含量提高[32-35]。从本试验结果看,当镍处理浓度在640~1 280 μg/L时,蕹菜生长量低于对照,脲酶活力和比活力均比对照要低,说明植株生长和脲酶活性受到抑制,且镍浓度为1 280 μg/L时,可溶性糖含量从0.3%提高到0.5%,增幅极显著,说明第一种可能性更大。
[1]Patrick H B,Ross M W,Earle E C. Nickel:a micronutrient essential for higher plants[J]. Plant Physiology,1987,85(3):801-803.
[2]Patrick H B,Ross M W,James T M. Effect of nickel deficiency on soluble anion,amino acid,and nitrogen level in barley[J]. Plant and Soil,1990,125(1):19-27.
[3]张西科,张福锁,李春俭. 植物生长必需的微量营养元素—— 镍[J]. 土壤,1996(4):167-179.
[4]朱祝军,Gere. 水稻缺镍症状和生理研究[J].植物营养与肥料学报,1997,9(3):224-228.
[5]Fernando G de M,Joana D B,Elcio F S,et al. Nickel availability in soil as influenced by liming and its role in soybean nitrogen metabolism[J]. Frontiers in Plant Science,2016,7(1358):1-12.
[6]Joe C P,Paulo M,Tiago T. Opinion - nickel and urease in plants:still many knowledge gaps[J]. Plant Science,2013,2(199-200):79-90.
[7]连兆煌. 无土栽培原理与技术[M]. 北京:中国农业出版社,1994:187-190.
[8]哈兹耶夫. 土壤酶活性[M]. 郑洪元,周礼恺,张德生译.北京:科学出版社,1980:111-113.
[9]礼恺. 土壤酶学[M]. 北京:科学出版社,1987:275-276.
[10]张龙翔,张庭芳,李令媛. 生化实验方法和技术[M]. 北京:人民教育出版社,1981:66-68.
[11]曹志华,罗静波. 鱼类脲酶比活力研究[J]. 湖北农学院学报,2001,21(1):39-42.
[12]林炎坤. 常用的几种蒽酮比色定糖法的比较与改进[J]. 植物生理学通讯,1989(4):53-55.
[13]李合生. 植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:192-194.
[14]刘国栋. 植物营养元素—— Ni[J]. 动植营养与肥料学报,2001,7(1):103-108.
[15]Mustapha A,Laëtitia J,Philippe L,et al. A physiological and molecular study of the effects of nickel deficiency and phenylphosphorodiamidate(PPD) application on urea metabolism in oilseed rape(Brassica napus L.)[J]. Plant and Soil,2013,362(1):79-92.
[16]张露,段碧华,苑亚会,等. 北京褐潮土中镍对植物根伸长和蚯蚓生殖的毒性效应研究[J].北京农学院学报,2015,30(2):35-39.
[17]白英,刘铮,刘占刚,等. 外源Ni在干旱区绿洲土壤—小麦系统中的迁移及其生物有效性[J].环境科学学报,2014,34(7):1801-1807.
[18]郭爱华,辛高伟,任静宇. 重金属铅、镍对苦菜根尖细胞分裂的影响[J]. 天津农业科学,2015,21(12):21-23.
[19]Geetika S,Mudaser A M,Elsayed F Abd-Allah,et al. Jasmonic acid modulates the physiobiochemical attributes,antioxidant enzyme activity,and gene expression in Glycine max under nickel toxicity[J]. Frontiers in Plant Science,2016,7(347):1-12.
[20]刘高琼,芮三亚,陶功俊,等. 镍对水培叶菜尿素同化的效应[J]. 植物生理学通讯,1992,28(4):252-354.
[21]黄锦孙,韦东普,郭雪雁,等. 田间土壤外源铜镍在小麦中的累积及其毒害研究[J]. 环境科学,2012,33(4):1369-1375.
[22]武振中,杨启良,贾维兵,等. 不同浓度镍胁迫对小桐子光合特性、形态特征和水分利用的影响[J]. 生态与农村环境学报,2015,31(5):767-773.
[23]王兵,马芳. 镍胁迫对十字花科5种植物生长及生理特性的影响[J]. 农业科技通讯,2014(12):87-88.
[24]孙凤金,郑孝胜,姜远茂,等. 不同供镍水平对苹果植株15N-尿素吸收分配的初步研究[J].园艺学报,2016,43(6):1141-1147.
[25]Ananda M,Ajit G,Tripathi A K,et al. A unique Ni2+-dependent and methylglyoxal-inducible rice glyoxalaseI possesses a single active site and functions in abiotic stress response[J]. Plant Journal for Cell & Molecular Biology,2014,78(6):951-63.
[26]岛田典司,安藤孝之. 镍对黄瓜、大麦生育及尿素同化的影响[J]. 日本土壤肥料科学杂志,1980,51(6):493-496.
[27]岛田典司,松尾明彦. 镍对尿素同化的影响[J]. 日本土壤肥料科学杂志,1986,57(3):257-263.
[28]姜凡琦,杨靖民,刘金华. 施镍对玉米苗期氮、磷吸收的影响[J]. 吉林农业大学学报,2013,35(5):568-572.
[29]罗虹,刘鹏,宋小敏. 重金属镉、铜、镍复合污染对土壤酶活性的影响[J]. 水土保持学报,2006,20(2):94-96.
[30]张忠庆,李韶山,刘金华,等. 外源NO供体硝普钠和镍对玉米生长及抗氧化酶活性的影响[J]. 西北农林科技大学学报(自然科学版),2015,43(8):79-85.
[31]王敬华,张立芳,施国新,等. 镍添加对水鳖(Hydrocharis dubia)叶片镍和营养元素含量以及氧化胁迫和抗氧化系统的影响[J]. 湖泊科学,2016,28(3):599-608.
[32]冷天利,蒋小军,杨远祥,等. 锌铬复合污染对水稻根系可溶性糖代谢的影响[J]. 生态环境学报,2007,16(4):1088-1091.
[33]张志雯,秦素平,陈于和,等. 硅对铬、铜胁迫下小麦幼苗生理生化指标的影响[J]. 华北农学报,2014(S1):229-233.
[34]孟衡玲,张薇,卢丙越,等. 铜胁迫对金银花幼苗生理指标的影响[J]. 北方园艺,2015(5):167-169.
[35]赵宇飞,沙文沛,任诗语,等. 铅胁迫对小叶黄杨幼苗生理生化指标的影响[J]. 贵州农业科学,2014(9):68-71.
(责任编辑 张辉玲)
Effects of nickel on growth and nutritional ingredients of water spinach (Ipomoea aquatica Forsk.)
LI Ning1,2,WANG Jin-sheng1,WANG Ye-yao1,3,WANG Xue-li4,QIN Xu-zhi2
(1. College of Water Sciences,Beijing Normal University,Beijing 100875,China;2. Guangxi Zhuang Autonomous Region Environmental Monitoring Centre,Nanning 530028,China;3. China Nationnal Environmental Monitoring Centre,Beijing 100012,China;4. College of Agriculture,Guangxi University,Nanning 530004,China)
The paper investigated the effects of various concentrations of nickel on growth,urease activity and some nutritional ingredients of hydroponic water spinach by supplying urea nitrogen as the only nitrogen source. The results indicated that appropriate quantity of nickel could enhance the growth,urease activity and amino acids synthesis of water spinach,while excessive nickel was toxic to water spinach. The treatment of 20 μg/L nickel resulted in the highest fresh weight of water spinach,12.1% higher than that in control,while significant reduction of water spinach fresh weight was detected with nickel supply higher than 320 μg/L,and significant plant damage symptoms were observed with 1 280 μg/L nickel treatment. This results also showed that appropriate concentrations of nickel (20-80 μg/L) could significantly increase the urease activity up to 13.6%,while the urease content also raised up to 19.0%,but excessive nickel supply (more than 640 μg/L) inhibited the urease activity. Physiological analysis demonstrated that adequate nickel concentration (60 μg/L) firstly enhanced the specific activity of urease,then participated in the urease synthesis to improve the urease yield,and ultimately improved the overall ureaseactivity. This results also identified that the total amino acids content increased simultaneously as the concentrations of nickel raised within the range of 0-40 μg/L with a highest increase of 44.7%,this effect became significant and stable with nickel over 10 μg/L;the content of soluble carbohydrates in leaves significantly increased with treatments of over-concentrated nickel (more than 640 μg/L).
hydroponic;water spinach;nickel treatment;urease;amino acid;soluble carbohydrates
S636.9
A
1004-874X(2017)03-0052-09
2016-11-28
广西科学研究与技术开发计划项目(桂科合14125008-2-30);广西农业重点科技计划项目(201519);环境保护部公益项目 (201309032);广西土壤环境污染与生态修复人才小高地项目;广西西江流域生态环境与一体化发展协同创新中心项目
黎宁(1972-),男,壮族,博士,教授级高级工程师,E-mail:348427557@qq.com
黎宁,王金生,王业耀,等.镍对蕹菜生长和营养成分的影响[J].广东农业科学,2017,44(3):52-60.
猜你喜欢
生物技术通报(2020年12期)2020-12-21 09:20:00
腐植酸(2020年1期)2020-11-29 00:15:44
家庭医药·快乐养生(2019年4期)2019-04-23 02:50:48
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07 11:14:32
中成药(2018年8期)2018-08-29 01:28:34
长江蔬菜(2016年10期)2016-12-01 03:05:34
恋爱婚姻家庭·养生版(2016年8期)2016-08-23 08:05:24
医学研究杂志(2015年9期)2015-07-01 17:28:24
生物灾害科学(2015年3期)2015-03-11 16:38:37
中国当代医药(2015年9期)2015-03-01 02:02:13
