
脉翅目细蛉科昆虫研究进展
2017-06-05 14:59师超凡
环境昆虫学报 2017年2期
师超凡,杨 强,任 东
(1. 中山大学地球科学与工程学院,广州 510275;2. 生物多样性演化与保护广东普通高校重点实验室,中山大学生物博物馆,中山大学生命科学学院,广州 510275;3. 首都师范大学生命科学学院,北京 100048;4. 河北地质大学地球科学博物馆,石家庄 050031)
脉翅目细蛉科昆虫研究进展
师超凡1,2,3*,杨 强2,3,4,任 东3
(1. 中山大学地球科学与工程学院,广州 510275;2. 生物多样性演化与保护广东普通高校重点实验室,中山大学生物博物馆,中山大学生命科学学院,广州 510275;3. 首都师范大学生命科学学院,北京 100048;4. 河北地质大学地球科学博物馆,石家庄 050031)
本文综述了脉翅目细蛉科化石与现生昆虫的分类学与系统发育学研究历史及进展。介绍了细蛉科成虫和幼虫的形态特征与生物学特性;分别回顾了现生与化石细蛉科昆虫的研究历史;分述了各地质历史时期细蛉科昆虫的属种及生物地理学分布;并介绍了近年细蛉科包括化石与现生属种的系统发育学研究进展,从细蛉科在脉翅目中的系统位置及细蛉科内亚科、属间的系统发育关系展开了讨论;并附世界化石与现生细蛉科昆虫的属种名录。
细蛉;化石;现生;分类学;系统发育
细蛉科Nymphidae是脉翅目蚁蛉亚目中一个较为古老的类群,最早的化石记录可追溯自中侏罗世。现生的细蛉科属种较少,包含8属33种;生物地理分布也比较局限,现生细蛉全部发现于澳大拉西亚。化石细蛉目前已发表15属24种,自中侏罗世至始新世都有化石记录,分布在欧亚大陆、北美洲和南美洲(Shietal., 2015;Myskowiaketal., 2016)。细蛉成虫体型中至大型;触角丝状,一般短于翅长的1/2;头部无单眼;前胸长大于宽,具翅胸节加宽;足中垫二裂;翅狭长或宽椭圆形,翅缘具缘饰,无翅疤,翅前缘域基部具有ScA脊,Sc与R1末端融合,亚前缘域横脉不完整;殖弧叶二分,背端部远离或靠近;幼虫触角刚毛状,上颚内侧中部有一个长齿,下唇须3节,中、后胸、腹节两侧有侧突(New, 1981;Shietal., 2015)。
中生代晚期是脉翅目适应辐射的重要时期,这一时期基本完成了科级阶元的分化。因此中生代晚期也是脉翅目中很多科的起源与早期演化的时期。我国东北中生代晚期陆相地层发育完好,主要包括燕辽生物群和热河生物群,发现有大量保存精美的脉翅目化石标本(Renetal., 2010;Wangetal., 2010;Shietal., 2011, 2012, 2013;Labandeiraetal., 2016)。其中细蛉科的化石标本数量虽然相对较少,却是该科在全世界的最早记录,其保存的特征是细蛉科早期演化、乃至脉翅目演化中的重要证据。
1 细蛉科形态特征和生物学
1.1 成虫
细蛉中至大形,头小至中形,无单眼,复眼大,触角丝状,绝大多数触角短于前翅长的1/2,仅中侏罗世的LiminymphaRenetEngel, 2007和现生的AustronymphesEsben-Petersen, 1914触角长于前翅长的1/2。前胸一般长大于宽,前胸窄于具翅胸节。足细长,步行足。胫节有或无端距。足中垫二分。
翅近等长,细长或宽圆,前翅翅长18-40 mm。现生细蛉很少有翅斑,化石细蛉多具翅斑,尤以白垩纪细蛉翅斑类型最为多样。具有缘饰,翅痣一般比较明显。前缘域宽度变化大,前缘横脉多,有时候分叉,前缘横脉间偶尔有纵向的小脉连接。ScA脊存在。中侏罗世和早白垩世的细蛉中偶见肩迴脉,现生的细蛉都不具有肩迴脉。Sc与R1末端融合,一般在翅端后与翅缘融合。亚前缘域基部横脉完全或较长但不完整,其余亚前缘横脉不完整,长度不等。Rs栉状分叉,Rs支脉间横脉众多,排列不规则,外阶脉明显。MA与Rs基部融合,后从Rs中分出。MP分叉位置多变,或早于MA分出处、或处于翅中部,在MA分出处之后、或在翅缘附近分叉,不易分辨出MP1和MP2。前翅CuA和后翅MP2一般在翅中部附近分叉,支脉较多,Nymphinae的前翅CuA和后翅MP2支脉间横脉较多,Myiodactylinae前翅CuA和后翅MP2支脉间无横脉或很少(图1)。
腹部细长,臀板简单,一般不特化,仅Osmylopssejunctus(Walker, 1853)种团的雄性臀板特化,较大,具有爪状修饰。臀胝明显。雌虫第七腹板后边缘多具修饰,产卵瓣凸,拱形,简单无装饰。亚生殖板显著。受精囊通常细长。雄虫第九腹板简单,圆拱形,第九背板有时向侧面延伸。殖弧叶存在,从中部分裂为两块,Nymphinae的两块殖弧叶互相远离,Myiodactylinae的殖弧叶背端靠近,有膜质相连。内突仅在Osmylopssejunctus种团存在。中突存在,有时很大,具有复杂的修饰;有时很小,位置多变。殖下片仅在部分属种中存在。下生殖板小。
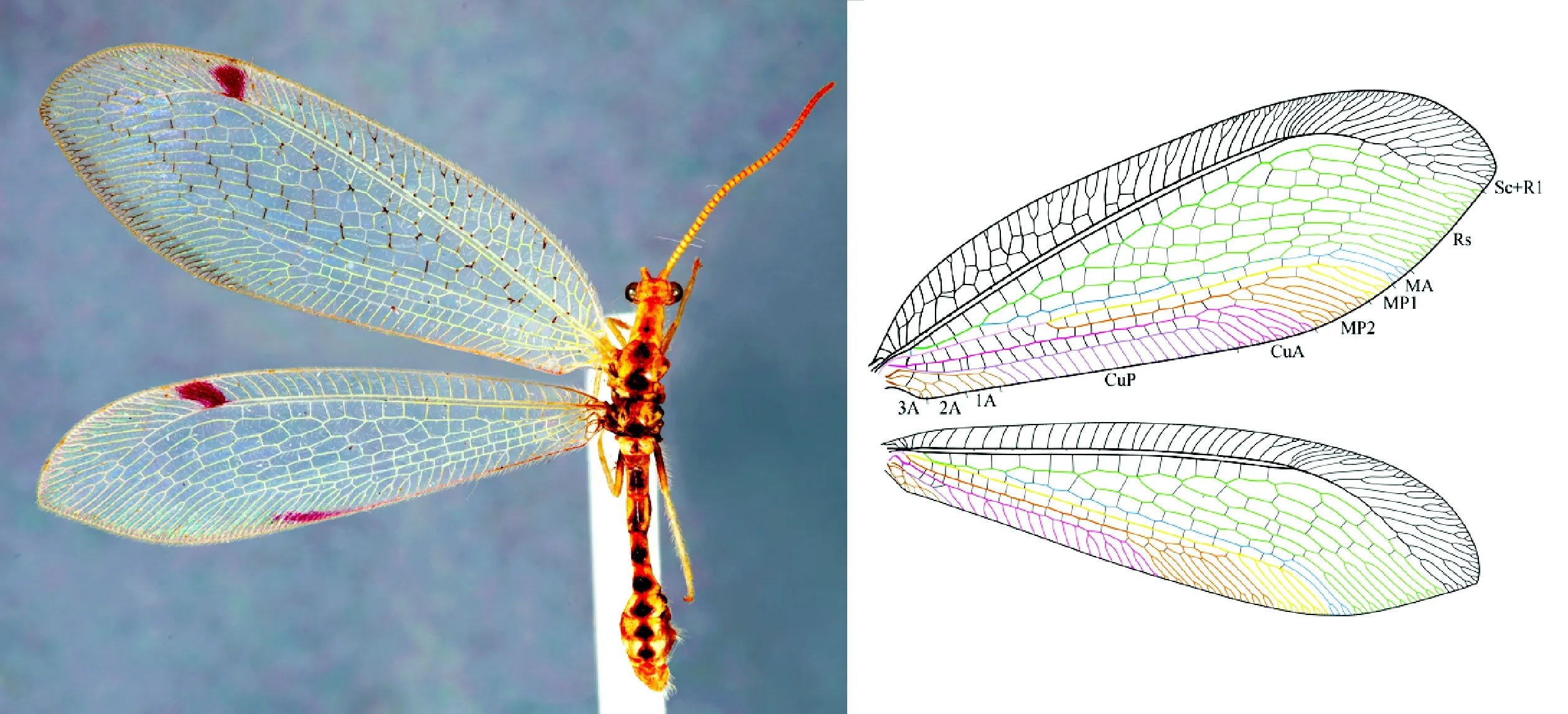
图1 细蛉科Norfolius howensis(Tillyard, 1917)照片和脉序图(仿Shi et al., 2015)Fig.1 Photo and venation drawing of Norfolius howensis (Tillyard, 1917) (Modified from Shi et al. 2015)
1.2 幼虫
幼虫头大,具有6个单眼,上颚常常分开,呈大于180度角。上颚内侧中部有一个大的齿,是细蛉科幼虫的明显特征。触角短于上颚的一半,触角鞭节明显比柄节和梗节细,下唇须3节。中、后胸、所有腹节都具有明显的侧突。
1.3 生物学
细蛉的卵聚产,成U-型排列,产卵时沿两个方向交替排卵30-40枚。细蛉的幼虫喜背负杂物,常在其长而卷曲的丝状背毛间夹杂垃圾碎片。NymphesLeach, 1814的老熟幼虫在沙石或土壤中作茧化蛹;OsmylopsBanks, 1913的老熟幼虫在植物或树皮中作茧化蛹(New, 1982, 1983a)。
2 细蛉科研究简史
2.1 现生细蛉科研究简史
现生细蛉科的研究始于1814年,英国动物学家William Elford Leach描述了细蛉科的第一个物种NymphesmyrmeleonoidesLeach, 1814,但未定科,这是细蛉科中最为常见的一个种。直到1842年,法国昆虫学家Jules Pierre Rambur依据该种建立了细蛉科Nymphides。此后,在十九世纪下半页和二十世纪初,Walker (1853)、Brauer(1866)、Mclachlan(1867-1877)、Gerstaecker(1885)、Banks(1910-1913)、Esben-Petersen(1914-1917)、Tillyard(1917-1926)、Navs(1921-1928)完成了细蛉科大量属种的描述。这一阶段,共建立7个现生属,但是对于属间的关系出现了不同的观点。
Handlirsch(1906-1908)将细蛉科中的部分属种分为一个独立的科Myiodactylidae,包括MyiodactylusBrauer, 1866,Osmylops。此后,Esben-Petersen(1914)、Comstock(1918)、Withycombe(1925)、Tillyard(1926)沿用这一分类系统,并多次讨论了各属的归属问题。
直到二十世纪八十年代,New(1981-1989)连续发表9篇关于细蛉的文章,首次对澳大利亚细蛉和东南亚的细蛉进行了全面的修订,描述了部分新属种,以及绝大部分已发表属种的生殖器结构并绘制了线条图,对部分细蛉的幼虫进行了描述。此后,Oswald(1997-1998)对Osmylops属进行了修订,重新描述4个种并描述了7个新种。将Osmylops分为2个种团:Osmylopssejunctusspecies group和Osmylopsarmatus(McLachlan, 1867)species group。Shi 等(2015)基于68个形态特征与4个基因的DNA分子序列分别进行了形态学和全证据的系统发育分析,将细蛉科分为2个亚科:Nymphinae,Myiodactylinae,并提出了细蛉科的科征、2个亚科的鉴定特征和现生属级检索表。
2.2 化石细蛉科研究简史
化石细蛉的研究也起步较早,但是分类系统一直存在争议。第一块化石细蛉Pronymphesmengeana(Hagen in Berendt, 1856)由Hagen于1854年首次提及,并于1856年描述,当时将该种归入现生属Nymphes(Hagen 1854, 1856)。Krüger(1923)年对该种进行了修订,建属PronymphesKrüger, 1923。这块标本是采集自始新世的波罗的海琥珀,Pronymphes也是化石细蛉中与现生细蛉最为接近的一个属。但是由于化石保存的极不完整,仅保存了前后翅翅基部的部分,翅端和身体的特征信息缺失。该种的模式标本有可能已在二战中损坏或丢失,Macleod(1970)依据前人的描述和线条图对该种进行了重新描述,并讨论了Pronymphes的分类位置和与现生属的系统关系。
Westwood也于1854年描述了采集自英国早白垩世的一块化石SialiumsipylusWestwood, 1854。这块标本仅保存了后翅的2/3翅端部分,由于特征信息的大量缺失,该种的分类位置被多次转移和修订(Westwood, 1854;Giebel, 1856;Hagen, 1862a, 1862b;Geinitz, 1884, 1887;Scudder, 1886, 1891;Handlirsch, 1906-1908, 1939;Martynova, 1949;Lambkin, 1988;Whalley, 1988;Jarzembowski, 1993;Jepsonetal., 2012)。依据后翅Rs,MA,MP1,MP2的分叉方式,本文认同Jepson等人(2012)的观点,将该种归入细蛉科。
Weyenbergh于1869年依据德国索罗霍芬晚侏罗世的一块标本描述Nymphitespriscus(Weyenbergh,1869),但将该种归入褐蛉科HemerobiusLinnaeus, 1758属。Haase(1890)依据该种与细蛉科多个现生属的相似性建属NymphitesHaase, 1890,并建立新组合Nymphitespriscus(Weyenbergh, 1869)。同时描述另一种NymphitesbraueriHaase, 1890,但Nymphitesbraueri后被Martins-Neto and Vulcano(1989)修正为Nymphitespriscus的异名。NymphitesbraueriHaase, 1890之后被认为是Nymphitespriscus(Weyenbergh, 1869)的异名(Shietal., 2013)。Handlirsch(1906-1908)依据该属建科Nymphitidae Handlirsch, 1908,认为Nymphitidae是与细蛉科亲缘关系相近的绝灭科。在此后的近一个世纪,Martynova(1949, 1958),Hong(1980),Makarkin(1990)等描述了该科的多个属种,并对该科的单系性及包含属进行了讨论。由于该科的模式属Nymphites是单模属,模式种Nymphitespriscus的正模标本仅保存了翅脉不完全的后翅,副模(原Nymphitesbraueri正模)也仅保存了后翅,因此造成了该科的系统位置一直存疑。直至Shi等2013年发现了我国中侏罗世地层中保存非常完整的该属新标本,对比模式种正模,对正模标本进行了重新描述,描述了该属的另一种,对属征进行了修订。依据修订后的属征,Nymphites应归属于细蛉科,因此也肯定了Martynova(1958),Makarkin and Menon(2005)等人的观点,Nymphitidae应该是细蛉科Nymphidae的异名。原被归属为Nymphitidae的Nymphites,Sialium,BaissoleonMakarkin,1990应归属为细蛉科(Jepsonetal., 2012;Makarkinetal., 2012)。
进入二十世纪,Carpenter(1929)、Panfilov(Dolinetal., 1980)、Makarkin(1990a, 1990b)、Ponomarenko(1992)描述了采集自欧洲细蛉科化石的4属7种,绝大多数属种采集自俄罗斯的白垩纪地层,此外还有德国和哈萨克斯坦晚侏罗世地层中的2个属Nymphites,MesonymphesCarpenter, 1929。这一时期描述的多数物种模式标本只是一个翅的一部分,残缺严重,如MesonymphesapicalisPonomarenko, 1992;DactylomyiusseptentrionalisMakarkin, 1990;此外,一些属种线条图不准确,描述不详尽,为后期细蛉科化石的分类造成了很大的困扰。
二十一世纪以来,随着古生物学的迅速发展,更多的学者对细蛉科化石进行了研究,细蛉科化石的研究更为全面和丰富。这一时期所描述的属种,在地质时期上,从中侏罗世至始新世;地理上,分布于中国、巴西、缅甸、美国、波罗的海等地(Martins-Neto, 2005;Menonetal., 2005;Ren and Engel,2007;Engel and Grimaldi, 2008;Archibaldetal., 2009;Shietal., 2013;Makarkinetal., 2013)。Shi 等(2015)对我国早白垩世地层发现的3属4种进行了描述,选取保存较好的化石细蛉属种与全部现生属,结合形态特征与现生属种的DNA分子序列分别进行了形态学与全证据的系统发育分析,确定了化石类群在细蛉科中的系统位置。Myskowiak 等(2016)利用Shi 等(2015)的矩阵,修改了部分化石特征后,进行了重新分析,获得了不同的系统发育关系。但是文中未指明重新分析所用矩阵为形态学矩阵还是全证据矩阵,也并未详述分析方法。 除作者提及的矩阵中化石特征改变外,尚有诸多可能影响系统发育树拓扑结构的原因,从文中却不得而知。因此,本文作者认同Shi等(2015)的系统发育分析结果。
3 细蛉科地质历史
3.1 侏罗纪
细蛉科的化石记录较为完整,最早的细蛉科化石发现于中国中侏罗世地层。目前已记录侏罗纪细蛉3属3种,分布于欧亚大陆。中侏罗世的细蛉已表现出了一定的多样性。LiminymphaRenetEngel, 2007触角长,翅狭长,前翅前缘域窄,具有肩迴脉,是细蛉科的根部类群。DaonymphesMakarkin,Yang, ShietRen, 2013的正模标本是一块仅保存了前翅翅基部2/3的标本,但保存了一部分很有意义的特征,如前缘域较宽,具有肩迴脉,因此Daonymphes有可能是细蛉科中Myiodactylinae的早期代表。NymphitesHaase, 1890是细蛉科中延续时间最久的1属,从中侏罗世至早白垩世的地层中都有Nymphites的发现。其中,NymphitesbimaculatusShi,MakarkinetRen, 2013是该属最早的代表。Nymphites翅狭长,前翅前缘域窄,后足胫节具有端距,可能是细蛉科中Nymphinae的早期代表。Nymphitespriscus是德国晚侏罗世地层中发现的,虽然模式标本只保存了不完整的后翅,但是根据后翅脉序的特征可以推断该种与中国中侏罗世的Nymphitesbimaculatus应属于同一属。该种的发现意味着Nymphites在中生代晚期是一个地理分布较广、在地质历史时期延续较久的属。在德国和哈萨克斯坦的晚侏罗世地层中还发现了同一属的2种MesonympheshageniCarpenter, 1929和MesonymphesrohdendorfiPanfilov 1980。Mesonymphes与Nymphites较为相似,仅在前缘横脉、MP2、CuA的分叉方式及支脉间横脉的特征上有差异。Mesonymphes也是一个广域、延续时长的属,除哈萨克斯坦和德国的晚侏罗世地层外,在俄罗斯的早白垩世地层中也有发现。鉴于形态特征相似、生物地理分布相近、存在时代比较一致,Mesonymphes与Nymphites可能是姐妹群。但是由于Mesonymphes的标本都保存不太完整,早期作者的描述和绘制线条图不够详尽准确。Shi 等(2015)为了避免因特征信息不准确导致系统发育分析结果有误,在其系统发育分析中,未将Mesonymphes列入。
3.2 白垩纪
白垩纪的细蛉类群丰富,分布更为广泛。目前已描述11属14种,分布于英国、俄罗斯、中国、缅甸、巴西(Westwood, 1854;Makarkin, 1990a, 1990b;Ponomarenko, 1992;Martins-Neto, 2005;Menonetal., 2005;Engel and Grimaldi, 2008;Shietal., 2015)。这一时期在古北区出现了侏罗纪的2属Mesonymphes与Nymphites的代表。此外,出现了大量的仅发现于白垩纪的属种。其中既包含Myiodactylinae的代表,如SpilonymphesShi,WintertonetRen, 2015,也包含大量Nymphinae的代表,如BaissoleonMakarkin, 1990a,SialiumWestwood, 1854。白垩纪时期也出现了2个广域属,Baissoleon在中国和俄罗斯都有发现,Sialium在中国和英国都有发现。其他属都是单模属。
3.3 古近纪
在美国和波罗的海的始新世的地层中发现了细蛉科2属3种(Pictet-Baraban and Hagen, 1856;Archibaldetal., 2009)。其中,发现于美国始新世早期的NymphesgeorgeiArchibaldetal., 2009被归入了现生属Nymphes,是该属的最早记录,也是现生细蛉属中唯一的1化石种。另外2种都采集自始新世中/晚期的波罗的海琥珀中,归属于化石属Pronymphes。该属仅在始新世的波罗的海琥珀中有所发现。2种的模式标本都是仅保存了翅的一部分,就其保存的翅脉特征而言,该属更接近于Nymphinae,但是由于保存特征太少,也未列入本文的系统发育分析中。
3.4 现代
现生的细蛉包含8属33种,主要分布于马来拉西亚区。历史上曾把现生的细蛉科分为2个独立的科Myiodactylidae和Nymphidae,但该分类系统自二十世纪八十年代以来就没有再使用过,但是Myiodactylidae和Nymphidae s. str.主要包含的属形成了细蛉科中的2个亚科,Nymphinae和Myiodactylinae。根据化石记录的情况来看,2个亚科的分支时间应该不晚于中侏罗世,与利用分子钟推测的分支时间基本一致(Wintertonetal., 2010;Shietal., 2015)。
4 系统发育学研究
Shi等(2015)根据形态学和全证据的系统发育分析,验证了细蛉科的单系性,其共有衍征包括:成虫足的中垫二裂,翅的亚前缘域具有不完整的横脉,幼虫具有6个单眼,触角鞭节较梗节细,下唇须3节,上颚内侧有一个很长的大齿,中、后胸和腹节具有侧突。尽管上述部分幼虫特征在蚁蛉科或蝶角蛉科中也有所发现,例如幼虫胸节和腹节的侧突,但是上述多个特征的同时存在显然只有在细蛉科中才能找到。
现生的细蛉明显地分为了2个亚科,即Nymphinae和Myiodactylinae(图2)。Nymphinae包括现生属Nesydrion,Austronymphes,Nymphes,其中Nesydrion是Austronymphes+Nymphes的姐妹群。该亚科的单系性得到了以下特征的支持:足的胫节具有端距,前翅的前缘域窄,CuA的支脉间具有多排横脉,1A短,后翅MP2的支脉间具有多排横脉,雄性生殖器中突具有大的侧叶,殖弧叶一分为二,互相远离。Austronymphes是单模属,而Nymphes和Nesydrion的单系性也得到了验证。
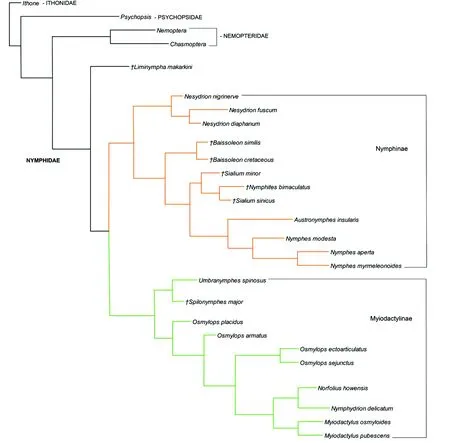
图2 细蛉科系统发育关系Fig.2 Phylogeny of extant and extinct Nymphidae注:†,为化石属种(仿Shi et al. 2015)。Note: †, indicates fossil species (Modified from Shi et al. 2015).
Myiodactylinae包括现生属UmbranymphesNew, 1987,Osmylops,NorfoliusNavs, 1922,Nymphydrion,Myiodactylus。支持该亚科的单系性的共有衍征包括:足的胫节无端距,前翅前缘域宽,CuA支脉间无横脉,1A长,后翅MP2支脉间横脉少,雄性生殖器中突无侧叶,殖弧叶一分为二,但在背侧互相靠近。Umbranymphes是单模属,是其他几个属的的姐妹群。在分析结果中,Osmylops是并系。Osmylops在早期的研究中被分为了2个种团:O.sejunctus种团和O.armatus种团(Oswald,1997,1998)。其中,O.sejunctus种团是单系群,而O.armatus种团仍是并系。Norfolius和Nymphydrion是单模属,互相成为姐妹群。此外,Myiodactylus的单系性也得到了很高的支持。
侏罗纪的Liminympha是其他所有细蛉的姐妹群。其余的化石细蛉分属于Nymphinae和Myiodactylinae 2个亚科。在Myiodactylinae中,早白垩世的单模属Spilonymphes与Umbranymphes是姐妹群,2属一起成为了其他属的姐妹群。其他的化石细蛉都被归入了Nymphinae中,Baissoleon是Nesydrion和其他属之间的过渡类群,而Sialium与Nymphites共同形成了一个单系,Sialium是一个并系。
5 讨论
尽管脉翅目的各科之间系统发育关系尚无定论,但是细蛉科在脉翅目中的系统位置还是相对比较稳定,是蚁蛉亚目Myrmeleontiformia中的一个基干类群(Aspöcketal., 2001, 2012; Aspöck, 2002; Wintertonetal., 2010)。蚁蛉亚目包括现生的蝶蛉科、细蛉科、蚁蛉科、蝶角蛉科和旌蛉科。蚁蛉亚目的系统发育关系主要有两种观点,主要的差异在于旌蛉科的系统位置。基于脉序、生殖器、幼虫上颚等的形态特征进行的分析认为细蛉科是蚁蛉科和蝶角蛉科的姐妹群,而蝶蛉科和旌蛉科共同与前述三科形成姐妹群(Aspöcketal., 2001, 2012; Aspöck, 2002; Engel and Grimaldi, 2008)。基于形态特征与DNA分子序列相结合的全证据系统发育分析得到的结果则认为细蛉科是蚁蛉科、蝶角蛉科、旌蛉科的姐妹群,而蝶蛉科是前述四科的姐妹群(New 1984; Oswald 1998; Wintertonetal., 2010)。在这些类群中,蝶蛉科无疑是最基干的类群,也是地质历史最久的一个科,自晚三叠世就有化石记录(Pengetal., 2010,2015; Yangetal., 2012)。细蛉科次之,其最早的化石记录是中侏罗世(Makarkinetal., 2013; Shietal., 2013)。其他类群的地质历史相对较短,最早的化石记录来自早白垩世(蚁蛉科和旌蛉科)甚至始新世(蝶角蛉科)(Martins-Neto, 2000; Grimaldi and Engel, 2005; Ren and Engel, 2008; Yangetal., 2012)。
在细蛉科的地质历史中,最早的代表是发现自中国中侏罗世地层(Makarkinetal., 2013; Shietal., 2013)。其中,Liminymphamakarkini是侏罗纪细蛉化石中唯一一个保存有触角的物种。触角长于前翅长的1/2(Ren and Engel, 2007),与现生的Nymphesaperta相似,而其他的现生细蛉触角较短。在侏罗纪和白垩纪的细蛉中还发现了肩迴脉,而在新生代以及现生的细蛉中却未发现,有可能是在晚期肩迴脉这一特征丢失了(Makarkinetal., 2013)。胫节的端距在中侏罗世和早白垩世的3个属中有所发现(Nymphites,Sialium,Baissoleon),这3个属归属于Nymphinae,是现生属Nymphes和Austronymphes的姐妹群。大多数的化石细蛉翅型狭长,类似于Nymphinae,但是从中侏罗世开始就出现了前翅前缘域较宽的类群,如Daonymphes前缘域宽,而Liminympha前缘域窄。所有的细蛉化石前翅MP在MA分出前双分叉,这一特征在现生细蛉中只有Nesydrion中有所发现,其他的现生细蛉MP分叉晚于MA分出处。其中的一些属种MP在近翅缘处分叉,很难辨认MP1和MP2,如Austronymphes,Myiodactylus,Umbranymphes和Nymphes的一些种。侏罗纪和部分早白垩世的细蛉,前翅MP2和CuA支脉数相近,这与现生的绝大多数Myiodactylinae以及Nesydrion相同。但是Myiodactylinae的前翅MP2和CuA与翅后缘平行,这一特征是仅这一亚科所有的。在Nymphinae和白垩纪的部分属(如Cretonymphes,Sialium)中,MP2和CuA的支脉间具有多排横脉,在其他的细蛉中,MP2和CuA支脉间的横脉很少或没有。Nymphes和Austronymphes的前翅CuA支脉众多,且支脉间有多排横脉,这一特征在蚁蛉科、蝶角蛉科、旌蛉科中也普遍存在。
现生的细蛉在历史中曾经被分为2个科Myiodactylidae和Nymphidae s. str.(Handlirsch, 1906-1908; Esben-Petersen, 1914; Withycombe, 1925; Tillyard, 1926)。其中,Myiodactylidae由相对原始的Myiodactylus和Osmylops组成。他们区别于Nymphidae s. str.(Nymphes,Austronymphes,Nymphidrion和Nesydrion)的特征在于翅型宽,前缘域宽,胫节无端距。Withytcombe(1925)甚至将Myiodactylidae归入溪蛉总科Osmyloidea,而Nymphidae归入蚁蛉总科Myrmeleontoidea。Shi等(2015)对所选类群的前翅的长宽比进行了测量和统计,发现在Myiodactylidae与Nymphidae s.str.之间并没有明显的差异。尽管Myiodactylus与Umbranymphes的长宽比明显小于Nymphidae s.str.,但是Osmylops各种之间的长宽比变化范围很大。因此,Myiodactylidae与Nymphidae s.str.就翅型上难以区分。胫节端距的存在与否和翅前缘域的宽度也不足以进行科级阶元的区分,如蚁蛉科中就既有有胫节端距的属,也有的属胫节无端距(New, 1985)。
尽管脉序与雄性外生殖器结构多样性较大,Nymphidae s.str.,包括Myiodactylidae在内,应该是一个单系。其共有衍征是足的中垫二分,ScA脊存在,亚前缘域横脉不完整,幼虫触角鞭节明显变细,上颚内侧中部具有一个大的齿,中、后胸和腹节有侧突。
Nymphinae包括现生属Nesydrion,Austronymphes,Nymphes和化石属Baissoleon,Sialium,Nymphites。其共有衍征是:至少后足胫节具有端距,翅基部窄,2/3处最宽,前缘域窄,前翅CuA支脉间有横脉,一对殖弧叶互相远离,中突有大的侧叶,与殖弧叶的背面顶端相邻。该亚科的现生属种都分布在澳大利亚。Nymphinae中包含中侏罗世的Nymphites,说明该亚科的起源不晚于中侏罗世。因此,Nymphinae和Myiodactylinae的分支时间也应不晚于中侏罗世,这一结论与依据分子数据推测的分支时间基本一致(Wintertonetal. 2010;Shietal. 2015)。
Myiodactylinae包括现生属Umbranymphes,Osmylops,Myiodactylus,Norfolius,Nymphydrion和白垩纪的化石属Spilonymphes。该亚科的现生属种的地理分布较Nymphinae更广。Osmylops和Myiodactylus在澳大利亚以外的新几内亚群岛也有发现。Umbranymphes仅分布于巴布亚。这一亚科的共有衍征是:足的胫节无端距;翅基部宽,前缘域宽,前缘横脉多分叉;前翅MP,CuA,CuP和后翅MP1,MP2,CuA直,与翅后缘平行;前翅CuA间无横脉;殖弧叶二分,背侧顶点互相靠近;中突小,或成球形,或腹端具有三齿结构,无明显的侧叶。
在Nymphinae中,Nesydrion属的共有衍征是翅痣不明显,在近翅端处具有色斑,前翅MP在MA分出前分叉,阳基侧突末端有齿,具有一对殖下片。Nesydrion是Nymphinae 其他属的姐妹群。该亚科其他的现生属和白垩纪的Baissoleon都具有明显的翅痣。此外其他的现生属前翅MP在MA分出后分叉。
白垩纪的Baissoleon属的共有衍征是前缘域在翅端处不变宽,前翅CuP双分叉,前翅CuA的支脉间具有一排横脉,这个特征在侏罗纪的细蛉中也可发现。Baissoleon是化石属Sialium和Nymphites,现生属Nymphes和Austronymphes的姐妹群。上述4个属的共有衍征是后翅CuA支脉间具有多排横脉,后翅CuP栉状分叉。
现生的Austronymphes和Nymphes是姐妹群,他们的共有衍征是前翅CuA支脉众多。Austronymphes的独征是前翅2A和3A闭合形成一个圆环状结构。Nymphes在翅端处具有玫瑰色或棕色的色斑,前翅CuP与后缘之间具有两排翅室。
Nymphinae的各属间形态差异明显,易于区别,而Myiodactylinae的各属间形态较为相似,早期用于区别属的特征变化较大,也不易于用于区别属,如利用前缘域基部的宽度来区分Myiodactylus和Osmylops,前缘阶脉的存在与否将Umbranymphes和Norfolius与其他属进行区分(New,1981,1987)。Norfolius属的雄性外生殖器结构特征独特,可以很容易的从其他属中区分出来,如阳基侧突末端特化成三指的结构,殖下片存在,中突下方有一对膜质的突出结构。Nymphidrion是单模属,模式标本只有一个被严重损坏的雌虫标本,该属与Myiodactylus与Norfolius在脉序上非常相似,但是其前翅CuA非常的长。Osmylops是细蛉科中物种最多的一个属,共有11个种,分为2个种团O.sejunctus种团和O.armatus种团(Oswald, 1997, 1998)。其中O.sejunctus种团的共有衍征是臀板特化,在末端形成了一个爪状结构,具有内突,在阳基侧突下方具有一对很长的膜质突出结构,中突腹面呈球形,具齿。O.armatus种团与O.sejunctus种团相比特征不太明显。O.armatus种团的2个种中突小,腹面成三齿状结构,但是这一特征在Norfolius也有发现。Myiodactylinae中存在Osmylops的并系性、部分属的共有衍征不够突出等问题。因此,今后还需要对该亚科的各属的标本进行重新的检视,进行进一步的研究,从而明确该亚科内的系统发育关系。
References)
Adams PA. Studies in the Neuroptera, with Special Reference to Wing Structure and Evolution in the Osmyloidea. [D]. Cambridge: Harvard University, 1958:120.
Archibald SB, Makarkin VN, Ansorge J. New fossil species of Nymphidae (Neuroptera) from the Eocene of North America and Europe [J].Zootaxa, 2009, 2157: 59-68.
Aspöck U, Haring E, Aspöck H. The phylogeny of the Neuropterida: Long lasting and current controversies and challenges (Insecta: Endopterygota) [J].ArthropodSystematics&Phylogeny, 2012, 70 (2): 119-129.
Aspöck U. Phylogeny of the Neuropterida (Insecta: Holometabola) [J].ZoologicaScripta, 2002, 31 (1): 51-55.
Aspöck U, Plant JD, Nemeschkal HL. Cladistic analysis of Neuroptera and their systematic position within the Neuropterida (Insecta: Holometabola: Neuropterida: Neuroptera) [J].SystematicEntomology, 2001, 26 (1): 73-86.
Banks N. Some Neuroptera from Australia [J].Psyche, 1910, 17 (3): 99-105.
Banks N. Synopses and descriptions of exotic Neuroptera [J].TransactionsoftheAmericanEntomologicalSociety, 1913, 39 (3/4): 201-242.
Brauer F. Zusätze und Berichtigungen zu Hagen’s Hemerobidarum Synopsis Synonymica und Beschreibung einer neuen Nymphiden-gattung: Myiodactylus osmyloides aus Australien[J].VerhandlungenderKaiserlich-KöniglichenZoologisch-BotanischenGesellschaftinWien, 1866, 16: 983-992.
Carpenter FM. A Jurassic neuropteran from the lithographic limestone of Bavaria [J].Psyche, 1929, 36 (3): 190-194.
Comstock JH. The Wings of Insects [M]. New York: Comstock Publishing Co., 1918: xviii , 430.
Engel MS, Grimaldi DA. Diverse Neuropterida in Cretaceous amber, with particular reference to the paleofauna of Myanmar (Insecta) [J].NovaSupplementaEntomologica, 2008, 20: 1-86.
Esben-Petersen P. Australian Neuroptera. Part I [J].ProceedingsoftheLinneanSocietyofNewSouthWales, 1914, 39: 635-645.
Esben-Petersen P. Australian Neuroptera. Part III [J].ProceedingsoftheLinneanSocietyofNewSouthWales, 1917, 42 (1): 203-219.
Geinitz FE. Beitrag zur Geologie Mecklenburgs [J].ArchivedesVereinsderFreundederNaturgeschichteinMecklenburg, 1887, 41: 143-216.
Geinitz FE. Ueber die Fauna des Dobbertiner Lias [J].ZDGG, 1884, 36: 566-583.
Gerstaecker CEA. Zwei fernere decaden Australischer Neuroptera Megaloptera [J].MitteilungenausdemNaturwissenschaftlichenVereinfürNeu-VorpommernundRugen, 1884 (1885), 16: 84-116.
Giebel CG. Fauna der Vorwelt mit Steter BerüCksichtidung der Lebenden Thiere Gerstaecker. Monographisch dargestellt von Dr. C. G. Giebel [M]. Leipzig : Zweiter Band: Gliederthiere. Erste Abtheilung: Insecten und Spinnen, 1856:xviii ,511 .
Grimaldi DA, Engel MS. Evolution of the Insects [M]. Cambridge: Cambridge University Press, 2005, xv,755.
Haase E. Bemerkungen zur Palaeontologie der Insecten [J].NeuesJahrbuchfürMineralogie,GeologieundPalaeontologie,Stuttgart, 1890, 1890 (2): 1-33.
Hagen HA. A comparison of the fossil insects of England and Bavaria [J].Entomologist’sAnnual, 1862b, 1-10.
Hagen HA. Die im Bernstein befindlichen Neuropteren der vorwelt bearbeitet von F. J. Pictet-Baraban [sic; for Pictet-del la Rive], und Dr. H. Hagen. In: Berendt GC, ed. Die im Bernstein Befindlichen Organischen Reste der Vorwelt Gesammelt, in Verbindung mit Mehreren Bearbeitet und Herausgegeben von Dr. Georg Carl Berendt. Bd. 2 [C]. Berlin: Nicholaischen Buchhandlung, 1856, 41-125.
Hagen HA. Ueber die Neuropteren aus dem lithographischen Schiefer in Bayern [J].Palaeontographica, 1862a, 10 (2): 96-145.
Hagen HA. Ueber die Neuropteren der Bernsteinfauna [J].VerhandlungendesZoologisch-BotanischenVereinsinWien, 1854, 4: 221-232.
Handlirsch A. Die Fossilen Insekten und die Phylogenie der Rezenten Formen. Ein Handbuch fur Palaontologen und Zoologen, Nine parts [M]. Leipzig: W. Engelmann, 1906-1908.
Handlirsch A. Neue Untersuchungen über die fossilen Insekten mit Ergän-zungen und Nachträgen sowie Ausblicken auf phylogenetische, palaeogeographische und allgemein biologische Probleme. II. Theil [J].AnnalendesNaturhistorischenMuseumsinWien, 1939, 49: 1-240.
Hong YC. Mesozoic Stratigraphy and Palaeontology of Basins of Shanxi, Gangsu and Ningxia. Part 2. [M]. Beijing: Geological Publishing House, 1980, 111-114. [洪友崇. 陕甘宁盆地中生代地层古生物. II. [M]. 北京: 地质出版社, 1980, 111-114]
Jarzembowski EA. A provisional checklist of fossil insects from the Purbeck Beds of Dorset [J].ProceedingsoftheDorsetNaturalHistoryandArchaeologicalSociety, 1993, 114: 175-179.
Jepson JE, Makarkin VN, Coram RA. Lacewings (Insecta: Neuroptera) from the Lower Cretaceous Purbeck Limestone group of southern England [J].CretaceousResearch, 2012, 34 (2012): 31-47.
Krüger L. Neuroptera succinica baltica. Die im baltischen Bernstein eingeschlossenen Neuroptera des Westpreussischen Provinzial-Museums (heute Museum für Naturkunde und Vorgeschichte) in Danzig [J].StettinerEntomologischeZeitung, 1923, 84: 68-92.
Labandeira CC, Yang Q, Santiago-Blay JA,etal. The evolutionary convergence of mid-Mesozoic lacewings and Cenozoic butterflies [J].ProceedingsoftheRoyalSocietyofLondonB, 2016, 283 (1824): 20152893.
Lambkin KJ. A re-examination of Lithosmylidia Riek from the Triassic of Queensland with notes on Mesozoic 'osmylid-like’ fossil Neuroptera (Insecta: Neuroptera) [J].MemoirsoftheQueenslandMuseum, 1988, 25: 445-458.
Leach WE. Zoological Miscellany; Being Descriptions of New, or Interesting Animals. Vol. 1 [M]. London: B. McMillan for E. Nodder & Son, 1814.
MacLeod EG. The Neuroptera of the Baltic Amber. I. Ascalaphidae, nymphidae, and psychopsidae [J].Psyche, 1970, 77 (2): 147-180.
Makarkin VN.Baissoleoncretaceusgen. and sp. nov. Fossil Neuroptera from the Lower Cretaceous of Baisa, East Siberia. 2. Nymphitidae [J].AnnalesdelaSociétéEntomologiquedeFrance(N.S.), 1990a, 26 (1): 125-126.
Makarkin VN, Menon F. New species of the Mesochrysopidae (Insecta, Neuroptera) from the Crato Formation of Brazil (Lower Cretaceous), with taxonomic treatment of the family [J].CretaceousResearch, 2005, 26 (2005): 801-812.
Makarkin VN. Novye setchatokrylye (Neuroptera) iz verkhnego mela Azii [New lacewings (Neuroptera) from the Upper Cretaceous of Asia]. In: Akimov IA, ed. Novosti Faunistiki i Sistematiki. Sbornik nauchnyhk trudov [News of Faunistics and Systematics. Collected Scientific Papers] [C]. Kiev: Naukova Dumka Publ., 1990b: 63-68.
Makarkin VN, Yang Q, Shi CF,etal. The presence of the recurrent veinlet in the Middle Jurassic Nymphidae (Neuroptera): A unique character condition in Myrmeleontoidea [J].Zookeys, 2013, 325: 1-20.
Makarkin VN, Yang Q, Peng YY,etal. A comparative overview of the neuropteran assemblage of the Lower Cretaceous Yixian Formation (China), with description of a new genus of Psychopsidae (Insecta: Neuroptera) [J].CretaceousResearch, 2012, 35(2012): 57-68.
Martins-Neto RG. New Neuroptera from Crato Formation, Lower Cretaceous, Araripe Basin, northeast Brazil [J].GAEA, 2005, 1 (1): 5-10.
Martins-Neto RG. Remarks on the neuropterofauna (Insecta, Neuroptera) from the Brazilian Cretaceous, with keys for the identification of the known taxa [J].ActaGeológicaHispanica, 2000, 35: 97-118.
Martins-Neto RG, Vulcano MA. Neurópteros (Insecta, Planipennia) da Formacçaão Santana (Cretaceo Inferior), bacia do Araripe, nordeste do Brasil. II. Superfamília Myrmeleontoidea [J].RevistaBrasileiradeEntomologia, 1989, 33 (2): 367-402.
Martynova OM. Mesozoic lacewings (Neuroptera) and their bearing on concepts of phylogeny and systematics of the order [J].TrudyPaleontologicheskogoInstituta,1949, 20: 150-170.
Martynova OM. Novye nasekomye iz Permskikh i Mezozoiskikh otlozhenii SSSR [New insects from the Permian and Mesozoic deposits of the USSR] [J].MaterialykOsnovamPaleontologii, 1958, 2: 69-94.
McLachlan R. Description of a new neuropterous insect from New Guinea, belonging to the genusMyiodactylus, Brauer [J].Entomologist'sMonthlyMagazine, 1877, 14: 85-86.
McLachlan R. New genera and species, &c., of neuropterous insects; and a revision of Mr. F. Walker's British Museum Catalogue of Neuroptera, part ii. (1853), as far as the end of the genus Myrmeleon [J].JournaloftheLinneanSocietyofLondon,Zoology, 1868, 9 (37): 230-281.
Menon F, Martins-Neto RG, Martill DM. A new Lower Cretaceous nymphid (Insecta, Neuroptera, Nymphidae) from the Crato Formation of Brazil [J].GAEA, 2005, 1 (1): 11-15.
Myskowiak J, Huang DY, Azar D,etal. New lacewings (Insecta, Neuroptera, Osmylidae, Nymphidae) from the Lower Cretaceous Burmese amber and Crato Formation in Brazil [J].CretaceousResearch, 2016, 59 (2016): 214-227.
New TR. A revision of the Australian Nymphidae (Insecta: Neuroptera) [J].AustralianJournalofZoology, 1981, 29 (6), 707-750.
New TR. A revision of the Australian Osmylidae: Kempyninae (Insecta: Neuroptera) [J].AustralianJournalofZoology, 1983b, 31 (3): 393-420.
New TR. A second species of Nymphidae (Neuroptera) from Papua New Guinea [J].NeuropteraInternational, 1985, 3: 187-189.
New TR. An oviruptor inNymphesLeach (Neuroptera: Nymphidae) [J].NeuropteraInternational, 1989a, 5 (4): 239-240.
New TR. Intergeneric relationships in recent Nymphidae. In: Gepp J, Aspöck H, Hölzel H, eds. Progress in World’s Neuropterology [C]. Graz: Proceedings of the 1st International Symposium on Neuropterology, 1984: 125-131.
New TR, Lambkin KJ. The larva ofNorfolius(Neuroptera: Nymphidae) [J].SystematicEntomology, 1989, 14: 93-98.
New TR. Nymphidae (Insecta: Neuroptera) from New Guinea. Invertebr [J].Taxon, 1987, 1 (8): 807-815.
New TR. Planipennia, Lacewings. Handbuch der Zoologie, Vol. 4 (Arthropoda: Insecta), Part 30 [M]. Berlin: Walter de Gruyter, 1989b, 132.
New TR. Some early stages ofOsmylops(Neuroptera: Nymphidae) [J].SystematicEntomology, 1983a, 8: 121-126.
New TR. The larva ofNymphesLeach (Neuroptera, Nymphidae) [J].NeuropteraInternational, 1982, 2 (2): 79-84.
Oswald JD. Review of thesejunctusspecies group of the split-footed Lacewing genusOsmylopsBanks (Neuroptera: Nymphidae), with remarks on the functional morphology of teminalic coupling [J].AustralianJournalofEntomology, 1997, 36: 351-358.
Oswald JD.OsmylopsBanks (Neuroptera: Nymphidae): Generic review and revision of thearmatusspecies group [J].JournalofNeuropterology, 1998, 1: 79-108.
Panfilov DV. New representatives of lacewings (Neuroptera) from the Jurassic of Karatau. In: Dolin VG, Panfilov DV, Ponomarenko AG,etal. eds. Fossil Insects of the Mesozoic [C]. Kiev: Naukova Dumka, 1980: 82-111.
Peng YY, Makarkin VN, Ren D. Diverse new Middle Jurassic Osmylopsychopidae (Neuroptera) from China shed light on the classification of psychopsoids [J].JournalofSystematicPalaeontology, 2015, 14 (4): 261-295.
Peng YY, Makarkin VN, Yang Q,etal. A new silky lacewing (Neuroptera: Psychopsidae) from the Middle Jurassic of Inner Mongolia, China [J].Zootaxa, 2010, 2663: 59-67.
Pictet-Baraban FJ, Hagen HA. Die im Bernstein befindlichen Neuropteren der Vorwelt. In: Berendt GC, ed. Die im Bernstein Befindlichen Organischen Reste der Vorwelt Gesammelt, in Verbindung mit Mehreren Bearbeitet und Herausgegeben von Dr. Georg Carl Berendt. Band 2, Abtheilung 2 [C]. Berlin: Nicolaische Buchhandlung, 1856: 41-125.
Ponomarenko AG. Setchatokrylye (Insecta, Neuroptera) iz nizhnego mela Zabaykal'ya [Neuroptera (Insecta) from the Lower Cretaceous of Transbaikalia] [J].PaleontologicalJournal, 1992, 1992 (3): 43-50.
Rambur JP. Histoire Naturelle des Insectes, Névroptères. Librairie Encyclopédique de Roret [M]. Paris: Fain et Thunot, 1842: xviii, 534.
Ren D, Engel MS. A split-footed lacewing and two epiosmylines from the Jurassic of China (Neuroptera) [J].AnnalesZoologici, 2007, 57 (2): 211-219.
Ren D, Shih CK, Gao TP,etal. Silent Stories-Insect Fossil Treasures from Dinosaur Era of the Northeastern China [M]. Beijing: Science Press, 2010, 1-409.
Scudder SH. A review of mesozoic cockroaches [J].MemoirsoftheBostonSocietyofNaturalHistory, 1886, 3 (13): 439-485.
Scudder SH. Index to the known fossil insects of the world including myriapods and arachnids [J].BulletinoftheUnitedStatesGeologicalSurvey, 1891, 71: 1-744.
Shi CF, Béthoux O, Shih CK,etal.Guyilingjianbonigen. et sp.n., an antlion-like lacewing, illuminating homologies and transformations in Neuroptera wing venation [J].SystematicEntomology, 2012, 37 (3): 617-631.
Shi CF, Makarkin VN, Yang Q,etal. New species ofNymphitesHaase (Neuroptera: Nymphidae) from the Middle Jurassic of China, with a redescription of the type species of the genus [J].Zootaxa, 2013, 3700 (3): 393-410.
Shi CF, Wang YJ, Ren D. New species ofGrammolingiaRen, 2002 from the Middle Jurassic of Inner Mongolia, China (Neuroptera: Grammolingiidae) [J].FossilRecord, 2013, 16 (2): 171-178.
Shi CF, Winterton S, Ren D. Phylogeny of split-footed lacewings (Neuroptera, Nymphidae), with descriptions of new Cretaceous fossil species from China [J].Cladistics, 2015, 31 (2015): 455-490.
Shi CF, Yang Q, Ren D. Two new fossil lacewing species from the Middle Jurassic of Inner Mongolia, China (Neuroptera: Grammolingiidae) [J].ActaGeologicaSinica-EnglishEdition, 2011, 85 (2): 482-489.
Tillyard RJ. Odonata, Planipennia, and Trichoptera from Lord Howe and Norfolk Islands [J].ProceedingsoftheLinneanSocietyofNewSouthWales, 1917, 42 (3): 529-544.
Tillyard RJ. The Insects of Australia and New Zealand (1stEdition) [M]. Sydney: Angus and Robertson, 1926: xi ,60.
Walker F. List of the Specimens of Neuropterous Insects in the Collection of the British Museum. Part II.(Sialides-Nemopterides) [M]. London: British Museum, 1853:iii,193-476.
Wang YJ, Liu ZQ, Wang X,etal. Ancient pinnate leaf mimesis among lacewings [J].ProceedingsoftheNationalAcademyofSciences, 2010, 107 (37): 212-215.
Westwood JO.Contributions to fossil entomology [J].QuarterlyJournaloftheGeologicalSocietyofLondon, 1854, 10: 378-396.
Weyenbergh H Jr. Sur les insectes fossiles du calcaire lithographique de la Bavière, qui se trouvent au Musée Teyler [J].ArchivesduMuséeTeyler, 1869, 2 (1): 247-294.
Whalley PES. Mesozoic Neuroptera and Raphidioptera (Insecta) in Britain [J].BulletinoftheBritishMuseumofNaturalHistory(Geology), 1988, 44(1): 45-63.
Winterton SL, Hardy NB, Wiegmann BM. On wings of lace: Phylogeny and Bayesian divergence time estimates of Neuropterida (Insecta) based on morphological and molecular data [J].SystematicEntomology, 2010, 35 (3): 349-378.
Withycombe CL. Some aspects of the biology and morphology of the Neuroptera with special reference to the immature stages and their possible phylogenetic significance [J].TransactionsofTheAmericanEntomologicalSociety, 1925, 1924: 303-411.
Yang Q, Makarkin VN, Winterton SL,etal. A remarkable new family of Jurassic Insects (Neuroptera) with primitive wing venation and its phylogenetic position in Neuropterida [J].PLoSONE, 2012, 7 (9): e44762.
Research progress of split-footed lacewings (Neuroptera, Nymphidae)
SHI Chao-Fan1,2,3*, YANG Qiang2,3,4, REN Dong3
(1. School of Earth Sciences and Engineering, Sun Yat-sen University, Guangzhou 510275, China; 2. Key Laboratory of Biodiversity Dynamics and Conservation of Guangdong Higher Education Institute, The Museum of Biology, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China; 3. College of Life Sciences,Capital Normal University,Beijing 100048,China; 4. Geoscience Museum, Hebei GEO University, Shijiazhuang 050031, China)
The morphology and biology of adult and larva of Nymphidae are described. The taxonomic history of fossil and extant nymphids are summarized. Nymphids from each period of the geological history are presented with biogeographic distribution. The newest phylogenic study including fossil and extent nymphids are referred to, with discussion of the phylogenetic relationships between Nymphidae subfamilies and among genera, as well as the relationship of Nymphidae and other lacewings. A checklist of extinct and extant Nymphidae is provided.
Nymphids; fossil; insects; taxonomy; phylogeny

附录:细蛉科属种名录
续上表

种Species产地Locality层位/时代Horizon/AgeMesonymphessibiricaPonomarenko,1992RussiaZazaFm/EarlyCretaceousMyiodactyluslyriformisNew,1987NewGuineaHolocene(extant)MyiodactylusmaculatusNew,1986AustraliaHolocene(extant)MyiodactylusnebulosusMcLachlan,1877NewGuineaHolocene(extant)MyiodactylusosmyloidesBrauer,1866AustraliaHolocene(extant)MyiodactylusparanebulosusNew,1987NewGuineaHolocene(extant)MyiodactyluspubescensBanks,1910AustraliaHolocene(extant)MyiodactylusroseistigmaEsben⁃Petersen,1917AustraliaHolocene(extant)MyiodactylussorongensisNew,1987NewGuineaHolocene(extant)MyiodactylusstriatusNew,1982AustraliaHolocene(extant)NesydriondiaphanumGerstaecker,1885AustraliaHolocene(extant)NesydrionfuscumGerstaecker,1885AustraliaHolocene(extant)NesydrionnigrinerveEsben⁃Petersen,1914AustraliaHolocene(extant)NesydrionpallidumBanks,1913AustraliaHolocene(extant)Norfoliushowensis(Tillyard,1917)AustraliaHolocene(extant)NymphesapertaNew,1982AustraliaHolocene(extant)NymphesmodestaGerstaecker,1885AustraliaHolocene(extant)NymphesmyrmeleonoidesLeach,1814AustraliaHolocene(extant)NymphesnigrescensNew,1982AustraliaHolocene(extant)NymphesparamyrmeleonidesNew,1982AustraliaHolocene(extant)†NymphesgeorgeiArchibald,MakarkinetAnsorge,2009USAEarlyEoceneNymphitespriscus(Weyenbergh,1869)GermanyLateJurassicNymphitesbimaculatusShi,MakarkinetRen,2013ChinaJiulongshanFm/MiddleJurassicNymphydriondelicatumBanks,1913AustraliaHolocene(extant)OlindanymphesmakarkiniMartins⁃Neto,2005BrazilCratoFm/EarlyCretaceousOsmylopsarmatus(McLachlan,1867)AustraliaHolocene(extant)OsmylopsbispinosusOswald,1998AustraliaHolocene(extant)OsmylopsclavatusOswald1998AustraliaHolocene(extant)OsmylopscruxOswald,1998AustraliaHolocene(extant)OsmylopsectoarticulatusOswald,1997AustraliaHolocene(extant)OsmylopshalberdisOswald,1998AustraliaHolocene(extant)OsmylopshirsutusNew,1987NewGuineaHolocene(extant)OsmylopsmonofoveatusOswald,1998NewGuineaHolocene(extant)OsmylopsnesosOswald,1998NewGuineaHolocene(extant)Osmylopsplacidus(Gerstaecker,1885)AustraliaHolocene(extant)Osmylopssejunctus(Walker,1853)AustraliaHolocene(extant)Pronymphesmengeana(Hagen,1856)BalticMiddle/LateEocenePronympheshoffeinsorumArchibald,MakarkinetAnsorge,2009BalticMiddle/LateEoceneRafaelnymphescratoensisMyskowiaketal,2016BrazilCratoFm/EarlyCretaceousSantananymphesponomarenkoiMartins⁃Neto,2005BrazilCratoFm/EarlyCretaceousSialiumminorShi,WintertonetRen,2015ChinaYixianFm/EarlyCretaceousSialiumsinicusShi,WintertonetRen,2015ChinaYixianFm/EarlyCretaceousSialiumsipylusWestwood,1854EnglandLulworthFm/EarlyCretaceousSpilonymphesmajorShi,WintertonetRen,2015ChinaYixianFm/EarlyCretaceousUmbranymphesspinosusNew,1987NewGuineaHolocene(extant)
国家科技基础条件平台工作重点项目(2005DKA21402);国家标本平台教学标本子平台(http://mnh.scu.edu.cn/);国家自然科学基金(31501881, 41602014);高校基本科研业务费(15lgpy28);中国博士后科学基金(2016M592570);河北省自然科学基金(C2015403012)
师超凡,女,1986年生,讲师,研究方向为昆虫分类与系统演化
*通讯作者Author for correspondence,E-mail: shichf5@mail.sysu.edu.cn
Received: 2017-02-21;接受日期Accepted: 2017-03-13
Q961
A
1674-0858(2017)02-0263-12
师超凡,杨强,任东.脉翅目细蛉科昆虫研究进展[J].环境昆虫学报,2017,39(2):263-274.
猜你喜欢
内蒙古科技与经济(2022年9期)2022-06-24
中国金属通报(2021年11期)2021-11-02
中国金属通报(2020年4期)2020-12-20
民用飞机设计与研究(2020年1期)2020-05-21
科技创新导报(2016年20期)2016-12-14
南方文学(2016年4期)2016-06-12
中国海上油气(2015年1期)2015-07-01
小说月刊(2015年2期)2015-04-23
小学科学(2015年2期)2015-03-11
小学科学(2015年1期)2015-03-11
