
紫花苜蓿的抗菌肽基因Rev4遗传转化
2017-05-23 20:34武慧宋明霞宁思淇武翠玲周晓馥
江苏农业科学 2017年7期
武慧+宋明霞+宁思淇+武翠玲+周晓馥
摘要:抗菌肽是机体先天免疫的重要组成成分,是许多生物体抵抗外来致病菌侵袭的第1道屏障。Indolicidin是一个13氨基酸残基肽,从牛的嗜中性粒细胞的胞浆小颗粒分离而来,具有广谱抗菌性,Rev4是其人工设计的反向序列。利用经典叶盘法,以根癌农杆菌A.GV3850介导紫花苜蓿遗传转化,GUS瞬时表达检测转化体系建立成功;压力筛选后叶片基因组DNA的PCR成功检测到抗菌肽基因条带,PCR产物回收测序得到13株转化紫花苜蓿。
关键词:紫花苜蓿;抗菌肽;遗传转化
中图分类号: Q785文献标志码: A文章编号:1002-1302(2017)07-0044-03
紫花苜蓿(Medicago sativa L.)为四倍体异花授粉的多年生草本植物,属于豆科苜蓿属,基因组大,是很好的水土保持植物与绿肥植物,在世界范围内广泛种植,以“牧草之王”著称[1]。长期以来,人们围绕经济有效地提高紫花苜蓿产量和品质两大目标进行了不懈的努力[2-3],但由于传统的紫花苜蓿育种方法时间长、成本高且可利用的种质资源有限,基因工程的方法更具有科学性和准确性[4]。
抗菌肽(antimicrobial peptides,AMPs)是生物体天然免疫的重要组成部分,是机体抵御外源致病菌的第1道屏障,由于其独特且多样的生物学功能和作用机制,引起了科学界的广泛关注和研究[5]。抗菌肽是自然进化的产物,它可以干扰病原微生物,使其细胞膜结构发生改变,通透性增加,进而起到杀菌作用,又因病原体不易对其产生抗药性,因此成为新型高效的抗生素替代品[6]。目前,人们一方面研究抗菌肽在先天免疫中发挥作用的机制,一方面设计新型抗菌、抗感染的抗菌肽制剂[7],抗菌肽研究前景非常广阔。
Indolicidin是一个13残基肽(IL-PWKWPWWPWRR-NH2),从牛的嗜中性粒细胞的胞浆小颗粒分离而来,具有广谱的抗菌和抗病毒活性[8]。针对外源抗菌肽基因转入植物体内容易被消化分解的不足,有人设计了抗菌肽Indolicidin的反向序列Rev4,合成的反向肽仍然有高的抗菌活性,且相比其他的抗菌肽,对植物蛋白酶的稳定性更高[9]。本研究利用经典叶盘法,以根癌农杆菌A.GV3850介导紫花苜蓿遗传转化,GUS瞬时表达检测转化体系建立成功;压力筛选后叶片基因组DNA的PCR成功检测到抗菌肽基因条带,PCR产物回收测序得到13株转化紫花苜蓿。本研究提高紫花苜蓿自身抗病性,紫花苜蓿作为绿色饲料具有重大的生态和社会效益。
1材料与方法
1.1植物材料
供试紫花苜蓿种子为吉林省农业科学院惠赠,吉林师范大学资源科学与绿色生产吉林省重點实验室保存。
1.2菌种和质粒
根癌农杆菌菌株A.GV3850-pKLP36.PcRIL(149)的pKLP36质粒上含有抗菌肽基因,根癌农杆菌 A.GV3850-PKYLX35S2(194)中含有gus基因。以上2种菌株均由美国迈阿密大学惠赠。
1.3PCR扩增引物序列
以Primer 5.0软件设计正向引物AnF和反向引物AnR。具体序列如下:AnF:5′-CACGAAGCTTACCATGGGATTTTTTCTCTTTTCAC-3′;AnR:5′-GACTGGAGCTCTTAAATAAGAGGCCATTTC-3′。
1.4试验试剂
GUS染色液,FAA固定液,离心柱琼脂糖凝胶回收试剂盒,Premix Ex Taq Version 2.0为TaKaRa公司产品,低熔点琼脂糖为Sigma公司产品,X-Gluc、5-溴-4-氯-3-吲哚半乳糖苷(X-gal)试剂、异丙基-β-D-硫代半乳糖苷(IPTG)为BBI公司产品,其他国产试剂购自北京鼎国生物公司。
1.5主要仪器设备
德国QIAGEN高通量组织研磨仪,美国Quawell超微量紫外分光光度计(UV-Vis Spectrophotometer Q5000),HC-2518R高速冷冻离心机,紫外凝胶电泳成像系统,Agilent温度梯度PCR扩增仪,Nikon荧光体式显微镜等。
1.6试验方法
1.6.1根癌农杆菌A.GV3850介导的苜蓿遗传转化将苜蓿组培苗叶片预培养3 d后,分别转入D600 nm为0.5的根癌农杆菌149和194菌液中,26 ℃、166 r/min振荡培养 15 min;倒掉菌液,无菌水冲洗3次,置于共培养培养基,25 ℃黑暗条件下,共培养3 d。
叶片转接到Kan压力筛选培养基上培养30 d,获得抗性植株;待抗性植株长到2~3 cm,切段后置于1/2 MS培养基中扩繁,智能人工气候箱中培养。
1.6.2转化植株的GUS表达检测取194侵染后5 d的转化叶片制成徒手切片,非转化植株叶片作为对照;取1 mL染色液放入2 mL 离心管中,将准备好的叶片浸泡在染液中,抽真空5 min,37 ℃过夜;取出染液,加入800 μL FAA固定液,颠倒混匀,倒掉,重复2次;依次加入30%、75%、95%乙醇,进行脱色,至阴性对照呈黄白色,倒掉95%乙醇,加入FAA固定液;压片后以Nikon荧光体式显微镜观察并采集图像。
1.6.3转化植株的PCR分子检测取149转化的苜蓿叶片,研磨后加入250 μL CTAB提取缓冲液(2% CTAB,100 mmol/L Tris-HCl pH值为8.0,20 mmol/L EDTA,1.4 mol/L NaCl,3% PVP)和10 μL β-巯基乙醇,65 ℃温浴;加入体积比为25 ∶24 ∶1的酚、三氯甲烷、异戊醇,混匀后 4 ℃、12 000 r/min离心10 min;取上清,无水乙醇醇沉1 h后离心30 min,75%乙醇洗涤沉淀2次;水酶混合液消化RNA;琼脂糖凝胶电泳检测。
PCR反应为25 μL体系,其中含12.5 μL Premix Ex Taq,正向引物AnF 1 μL,反向引物AnR 1 μL,模板DNA 1 μL,ddH2O 9.5 μL;PCR程序:94 ℃ 10 min;94 ℃ 1 min,58 ℃ 1 min,72 ℃ 1 min,35个循环;72 ℃ 10 min。
1.6.4PCR产物测序对阳性质粒PCR产物和68个呈阳性的PCR产物进行切胶回收,送至北京六合华大基因科技有限公司进行测序。
2结果与分析
2.1转化植株的Gus表达检测
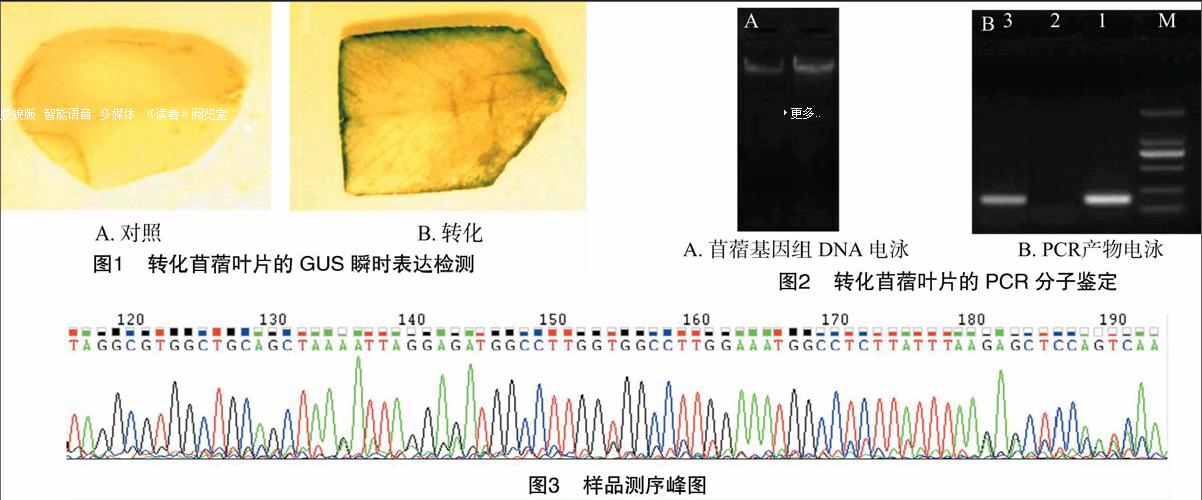
通过GUS组织化学定位法,对根癌农杆菌194菌液侵染的苜蓿组培苗叶片进行GUS瞬时表达检测,转化植株的GUS瞬时表达呈现出蓝色斑点(图1-B),而对照叶片无蓝色斑点(图1-A),初步证明转化体系的成功建立。
2.2转化植株的分子鉴定
本研究利用CTAB法分别提取对照和转化苜蓿组培苗的基因组DNA,琼脂糖凝胶电泳检测,结果如图2-A所示,原种与转化种苜蓿基因组DNA都较完整、浓度大、无脱尾。根癌农杆菌菌株149的pKLP36质粒上含有抗菌肽基因,包括烟草中PR-1b部分编码区及抗菌肽基因RIL,长约 200 bp;根据图2-B可知,阳性质粒PCR产物(泳道1)在 100~250 bp 处有条带,紫花苜蓿原种基因组DNA的PCR产物(泳道2)无目标条带,而转化植株基因组DNA的PCR产物有较为清晰的目标条带(泳道3)。
2.3PCR产物测序
对阳性质粒及PCR呈阳性的产物进行回收后测序,发现测序片段与149质粒对比结果一致,信号值基本在正常范围内,即150~500,测序片段质量较高,峰图噪音较低,模板纯度较高。由图3可知,149质粒和13个样品PR-1b编码区部分基因以及抗菌肽基因RIL区段均无突变碱基,即测序成功。
3讨论
3.1利用GUS瞬时表达检测转化体系
Jefferson等首次提出β-葡糖苷酸基因GUS可作为植物遗传转化的报告基因之后,在许多国家都得到了广泛应用,其方法操作简单,灵敏度高,在基因工程领域得到广泛地应用。可以用于瞬时表达检测的报告基因还有绿色荧光蛋白基因(greenfluorescent protein,GFP),但由于有些植物组织中有自发荧光,会影响GFP检测的准确性,产生弱荧光背景,检测弱启动子驱动的GFP活性较难,致使转基因植物的筛选受到限制[10]。GUS酶有产物的放大作用,可以通过GUS染色来进行检测,而未转化植株无GUS活性,因此GUS瞬时表达检测对遗传转化的检测更加简便、可靠[11]。因此,GUS是目前转基因植物中应用最为广泛的报道基因。关于植物叶片的脱色问题,Bowling等通过30%、75%、95%对拟南芥进行脱色,结果对照呈白色,脱色效果好[12]。因此,本试验中脱色步骤借鉴此方法,且脱色效果很好,便于结果观察。
3.2改良的CTAB法提取苜蓿基因组DNA
苜蓿叶片中常含有糖类等物质,使其基因组DNA的提取受限。试验结果表明:经典的CTAB提取缓冲液法最大的优点是能除去多糖,可用于草本植物基因组DNA提取。本研究在传统CTAB法的基础上加入了PVP,可以有效地裂解植物组织细胞,保证核酸完整性。同时,PVP可以与酚类产生络合物,防止多酚对DNA的污染,还可与多糖结合,除去糖类物质。此外,加入抗氧化剂β-巯基乙醇,可有效地防止酚氧化成醌,避免褐变,使酚容易去除。利用改良法提取的苜蓿基因组DNA完整性好、亮度高、浓度较大,可满足PCR检测要求。
3.3抗菌肽基因的遗传转化前景
抗菌肽分子较小,动物源抗菌肽在植物体内易被蛋白酶快速降解,成为外原抗菌肽在植物体内表达的限制因素。但在作物中表达动物源与植物源的肽是可能的,可通过消除蛋白酶敏感位点来修饰肽,或設计反向类似物,从而实现对病原抗性的提高[13]。Xing等采用来自异源物种中的肽来抵抗植物病原,研究了来自动物源的肽,筛选了肽对植物蛋白酶的敏感性,结果发现,基于反向序列的肽拥有特别强的对植物叶片中蛋白酶的抵抗能,并同时拥有抗菌活性[14]。本研究以根癌农杆菌介导抗菌肽基因Rev4的遗传转化并获得成功,转化植株作为绿色饲料具有重大的生态和社会效益。
参考文献:
[1] Samac D A,Bucciarelli B,Miller S S,et al. Transgene silencing of sucrose synthase in alfalfa (Medicago sativa L.) stem vascular tissue suggests a role for invertase in cell wall cellulose synthesis[J]. BMC Plant Biology,2015,15(1): 283.
[2]Aung B,Gruber M,Amyot L,et al. MicroRNA156 as a promising tool for alfalfa improvement[J]. Plant Biotechnology Journal,2015,13(6): 779-790.
[3]Zhang Y M,Liu Z H,Wen Z Y,et al. The vacuolar Na+-H+ antiport gene TaNHX2 confers salt tolerance on transgenic alfalfa (Medicago sativa)[J]. Functional Plant Biology,2012,39(8): 708-716.
[4]Zhang Y,Zhang H,Liu Z,et al. The wheat NHX antiporter gene TaNHX2 confers salt tolerance in transgenic alfalfa by increasing the retention capacity of intracellular potassium[J]. Plant Molecular Biology,2015,87(3): 317-327.
[5] Anupa G,Bhat M A,Srivastava A K,et al. Cationic antimicrobial peptide,magainin down-regulates secretion of pro-inflammatory cytokines by early placental cytotrophoblasts[J]. Reproductive Biology and Endocrinology,2015,13(1): 121.
[6]Urfer M,Bogdanovic J,Lo Monte F,et al. A peptidomimetic antibiotic targets outer membrane proteins and disrupts selectively the outer membrane in escherichia coli[J]. The Journal of Biological Chemistry,2016,291(4): 1921-1932.
[7]Chou S,Shao C,Wang J,et al. Short,multiple-stranded β-hairpin peptides have antimicrobial potency with high selectivity and salt resistance[J]. Acta Biomaterialia,2016,30: 78-93.
[8]Selsted M,Novotny M,Morris W,et al. Indolicidin,a novel bactericidal tridecapeptide amide from neutrophils[J]. The Journal of Biological Chemistry,1992,267(7): 4292-4295.
[9]Li Q,Lawrence C B,Davies H M,et al. A tridecapeptide possesses both antimicrobial and protease-inhibitory activities[J]. Peptides,2002,23(1): 1-6.
[10]Zhang Q,Walawage S,Tricoli D,et al. A red fluorescent protein (DsRED) from Discosoma sp. as a reporter for gene expression in walnut somatic embryos[J]. Plant Cell Reports,2015,34(5): 861-869.
[11]Liu X,Lang S,Su L,et al. Improved agrobacterium-mediated transformation and high efficiency of root formation from hypocotyl meristem of spring Brassica napus ‘Precocity cultivar[J]. Genetics and Molecular Research,2015,14(4): 16840-16855.
[12]Bowling S,Guo A,Cao H,et al. A mutation in Arabidopsis that leads to constitutive expression of systemic acquired resistance[J]. The Plant Cell,1994,6(12): 1845-1857.
[13]Anantharaman A,Sahal D. Reverse engineering truncations of an antimicrobial peptide dimer to identify the origins of potency and broad spectrum of action[J]. Journal of Medicinal Chemistry,2010,53(16): 6079-6088.
[14]Xing H,Lawrence C,Chambers O,et al. Increased pathogen resistance and yield in transgenic plants expressing combinations of the modified antimicrobial peptides based on indolicidin and magainin[J]. Planta,2006,223(5): 1024-1032.
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13
潍坊学院学报(2020年6期)2020-11-22
养殖与饲料(2019年7期)2019-02-25
广东饲料(2016年5期)2016-12-01
国外医药(抗生素分册)(2016年5期)2016-07-12
湖北畜牧兽医(2015年11期)2016-01-11
特产研究(2014年4期)2014-04-10
