
黄连幼苗期的解剖结构研究
2017-05-17 09:43杨再辉湖北省利川市农业局湖北利川445400
长江大学学报(自科版) 2017年6期
杨再辉 (湖北省利川市农业局,湖北 利川 445400)
周存宇,杨朝东 (湿地生态与农业利用教育部工程研究中心(长江大学) ,湖北 荆州 434025)
黄连幼苗期的解剖结构研究
杨再辉
(湖北省利川市农业局,湖北 利川 445400)
周存宇,杨朝东
(湿地生态与农业利用教育部工程研究中心(长江大学) ,湖北 荆州 434025)
利用普通光学和荧光显微镜对黄连(Coptischinensis)幼苗期地下部分进行了解剖结构和屏障结构研究,结果表明:黄连幼苗期不定根由二原型维管柱、内皮层、皮层、外皮层和表皮组成;胚轴由维管柱、内皮层、皮层、皮下层和表皮组成。黄连幼苗期不定根屏障结构为木质化和强烈栓质化的内皮层和外皮层;胚轴具木质化和强烈栓质化的内皮层,外侧具皮下层和角质层。
黄连(Coptischinensis);解剖结构;组织化学;幼苗期
黄连(Coptischinensis)属毛茛科药用植物,主要栽培于我国西南和中南的山区[1],具很高的医药和经济价值[2,3]。黄连栽培环境条件要求高,喜湿阴凉,不耐高温和强光;生长缓慢,通常要5~6a才能采收。因此,黄连的栽培区域受到气候环境的制约,采收周期长,影响农民收入和种植积极性。目前栽培黄连的解剖结构已有研究[4,5],黄连素在体内的分布规律也有报道[6~9],但缺乏对黄连幼苗期的形态学和解剖结构研究。因栓质化和木质化阻碍水和矿质离子等从植物质外体途径与环境发生交换,植物细胞壁中的凯氏带也称为质外体屏障结构[10]。近年来,常用硫酸消化法[11]和硫氢酸黄连素-苯胺兰对染法研究凯氏带[12];苏丹红7B染色法、蓝色激发光下不染色和硫氢酸黄连素-苯胺兰对染为研究细胞壁栓质层的3种方法[13~15];木质化细胞壁染色方法中硫氢酸黄连素-苯胺兰对染法比盐酸-间苯三酚染色法易显色观察[11,12]。本研究选用上述方法对黄连幼苗期地下部分的形态学和解剖结构进行了观察,试图找出育苗中的限制因素,并提出改进优化栽培措施。
1 材料与方法
黄连种子由湖北利川箭竹溪黄连专业合作社提供。2014年12月中旬,先将塑料盆中装入田园土和细沙混合土(二者比例4︰3)并浇透水,再将黄连种子播在表层下,置于荆州长江大学西校区植物园水杉林下,2015年3月中旬,种子萌发率达到90%,子叶出土,种子根长约10mm;4月初所有个体长出第一片真叶,不定根长约20mm;到8月底所有个体长出5片左右真叶,不定根长约50mm。从3月中旬开始取样完整植株,每2周取样1次,直到8月底。
用自来水洗掉植株上的泥土,保持完整植株,放入FAA固定液中固定。将FAA固定材料置于蒸馏水中30min,在可调节放大倍数的解剖镜下,用双面刀片切片距根尖5、10、20 、30、40、50mm和胚轴中部分别切片。试验重复5次。采用0.1% (w/v) 硫氢酸黄连素和0.5% (w/v) 苯氨兰对染切片后观察初生壁凯氏带和次生壁木质化,在荧光显微镜下凯氏带呈现生动黄色,木质化细胞壁呈现呆滞黄色[12]。盐酸-间苯三酚染色切片的木质化胞壁为樱桃红色[11]。用0.1% (w/v)苏丹红7B溶液染色切片确定细胞壁栓质化呈现红色[13],并拍照记录观察结果。
陶瓷绘画工匠们在珐琅彩的基础上,发明了“玻璃白”这一不透明的乳白色颜料。在玻璃白表面绘制新彩颜料,经过烧制可以使颜色变淡雅。又能适应对鸟嘴,眼睛,抓等飞禽某个部位进行写实的表现。粉彩只有做到这样才有可能从色彩对比强烈的古彩基础上经过玻璃白的淡化演变为粉彩,这样整幅画色彩柔和,呈现细腻雅致的粉彩艺术风貌。可以看出,玻璃白的出现为粉彩的形成有着重要的作用。
2 结果与分析
2.1 黄连子叶期地下部分的解剖结构和组织化学特征
距黄连种子根根尖5mm切片,种子根为二原型,解剖结构由内而外依次为初生木质部、内皮层、皮层、外皮层和表皮;外皮层具凯氏带,轻微木质化和强烈栓质化,内皮层仅少数细胞具凯氏带,几乎无木质化和栓质化 (图1A、B、C)。距根尖10mm,具初生木质部,外皮层明显木质化和强烈栓质化;内皮层具凯氏带,轻微木质化和部分细胞强烈栓质化,具通道细胞;初生木质部具后生木质部,部分表皮细胞脱落(图1D、E、F)。胚轴解剖结构由内而外依次为初生木质部、内皮层、皮层和表皮;内皮层具凯氏带,轻微木质化和部分细胞强烈栓质化,具通道细胞;表皮细胞外具角质层(图1G、H、I)。
电压暂降的严重程度主要由以下条件制约:各网络节点和故障点的远近、各种变压器组的事故发生位置、连接方式、故障点周围有无电源、整个线路的阻抗情况、电压暂降前后故障点的电压情况、断路器通断的快慢、保险丝的熔断时间长短、各变压器组的失效率,还有系统规定的自动重合闸的重置次数等。
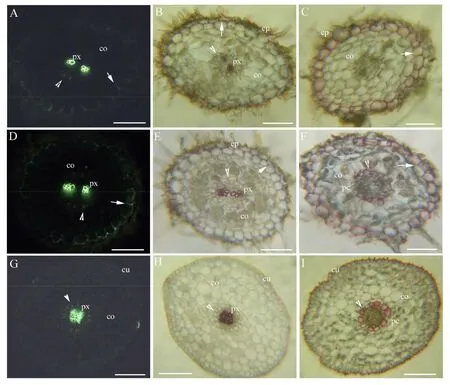
co:皮层;cu:角质层;ep:表皮;pc:通道细胞;px:初生木质部。A~C:距根尖5mm横切;D~F:距根尖10mm横切;G~I:胚轴横切。A.硫氢酸黄连素-苯氨兰染色,示初生木质部、皮层、内皮层(箭头)、外皮层(箭);B.盐酸-间苯三酚染色,示初生木质部、 皮层、内皮层(箭头)、外皮层(箭)、表皮细胞;C.苏丹红7B染色,示内皮层(箭头)、皮层、外皮层栓质化(箭)、 表皮细胞。D.硫氢酸黄连素-苯氨兰染色,示初生木质部、皮层、内皮层(箭头)、外皮层(箭);E.盐酸-间苯三酚染色,示初生木质部、皮层、内皮层 (箭头)、外皮层(箭)、表皮细胞;F.苏丹红7B染色,示初生木质部、皮层、内皮层 (箭头)、通道细胞、外皮层 (箭)。G.硫氢酸黄连素-苯氨兰染色,示初生木质部、内皮层(箭头)、皮层、角质层;H.盐酸-间苯三酚染色,示初生木质部、内皮层(箭头)、皮层、角质层;I.苏丹红7B染色,示内皮层(箭头)、通道细胞、皮层、角质层。图1 黄连子叶期种子根和胚轴的显微初生结构(标尺= 50 μm)
3.1 黄连幼苗期的解剖结构
黄连幼苗期可分为子叶期和真叶期,其地下部分由种子根、不定根和胚轴组成,种子根仅在子叶期发育,随后在真叶期停止生长或者部分脱落,不定根和胚轴进一步生长发育,而胚轴不再伸长。黄连子叶期解剖结构为初生结构,种子根和不定根为二原型维管柱、内皮层、皮层、外皮层和表皮;胚轴由初生木质部、内皮层、皮层和表皮组成;真叶期不定根和胚轴具次生木质部,不定根仅表皮脱落,胚轴具木质化皮下层。因此,黄连幼苗期不定根由二原型维管柱、内皮层、皮层、外皮层和表皮组成;胚轴由维管柱、内皮层、皮层、皮下层和表皮组成。
黄连不定根初生结构和组织化学特征与上述种子根的相同,故省略。距不定根尖20mm,木质部具次生结构,内皮层仍轻微木质化和强烈栓质化,并具通道细胞;外皮层明显木质化和强烈栓质化;表皮细胞几乎脱落(图2A、B、C)。距根尖40mm,次生木质部接近圆形,内皮层具凯氏带,仍轻微木质化和强烈栓质化,仅有少数通道细胞;皮层和外皮层保持完整结构,外皮层木质化略增强和强烈栓质化(图2D、E、F)。胚轴木质部具次生结构并扩大,内皮层具凯氏带,轻微木质化和强烈栓质化,具少数通道细胞;皮下层部分细胞木质化 (图2G、H、I)。
本研究中的黄连不定根结构与前人研究的黄连或其他植物不定根[4,5]比较,其初生结构都相似。常见双子叶植物不定根中的次生结构发育后,皮层通常脱落,由维管柱鞘细胞发育而来的木栓层代替皮层的保护功能;而黄连不定根的内皮层、皮层和外皮层继续保持完好状态。黄连胚轴具凯氏带并强烈栓质化的内皮层,而水稻(Oryzasativa)胚轴为散生维管束鞘细胞具凯氏带、第一节茎的内皮层和其厚壁组织内散生维管束鞘细胞具凯氏带[16];水生毛茛(Ranunculustrichophyllus)的茎具内皮层,叶柄散生维管束鞘细胞具凯氏带[17],香蒲属(Typha)植物根茎和天胡荽(Hydrocotylesibthorpioides)匍匐茎也具内皮层[18,19]。
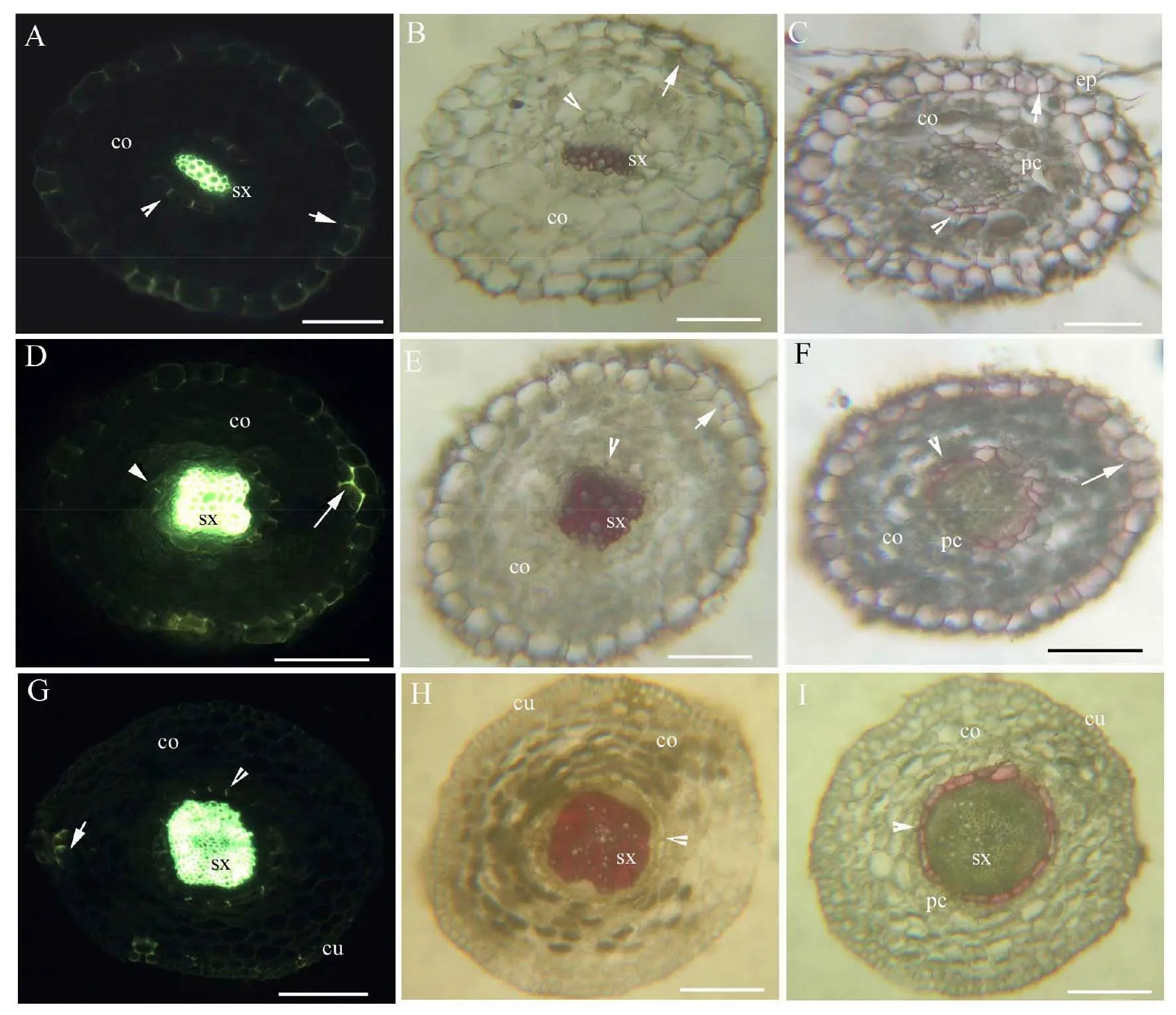
co:皮层;cu:角质层;ep:表皮;pc:通道细胞;sx:次生木质部。A~C:距根尖20mm横切;D~F:距根尖40mm横切;G~I:胚轴横切。A.硫氢酸黄连素-苯氨兰染色,示次生木质部、皮层、内皮层(箭头)、外皮层(箭);B.盐酸-间苯三酚染色,示次生木质部、内皮层 (箭头)、皮层、外皮层 (箭);C.苏丹红7B染色,示内皮层 (箭头)、通道细胞、皮层、外皮层 (箭)、 表皮细胞。D.硫氢酸黄连素-苯氨兰染色,示次生木质部、内皮层(箭头)、皮层、 外皮层(箭);E.盐酸-间苯三酚染色,示次生木质部、内皮层 (箭头)、皮层、外皮层 (箭);F.苏丹红7B染色,示内皮层 (箭头)、通道细胞、皮层、外皮层 (箭)。G.硫氢酸黄连素-苯氨兰染色,示次生木质部、内皮层(箭头)、皮层、皮下层(箭)、角质层;H.盐酸-间苯三酚染色,示次生木质部、内皮层(箭头)、皮层、角质层;I.苏丹红7B染色,示次生木质部、内皮层(箭头)、通道细胞、皮层、角质层。图2 黄连真叶期不定根和胚轴的显微次生结构(标尺= 50 μm)
黄连不定根屏障结构在真叶期为具凯氏带,轻微木质化和强烈栓质化的内皮层和外皮层组成;胚轴屏障结构在真叶期为具凯氏带,轻微木质化和强烈栓质化的内皮层,表皮外角质层和木质化皮下层组成[12]。
3 讨论
黄连种子根屏障结构在子叶期为具凯氏带,轻微木质化和强烈栓质化的内皮层和外皮层组成;胚轴屏障结构在子叶期为具凯氏带,轻微木质化和强烈栓质化的内皮层,表皮外角质层组成。
2.2 黄连真叶期地下部分的解剖结构和组织化学特征
清朝统治者经常举行木兰行围和大型宴会,借以彰显国力强盛、民生富庶。管世铭以诗歌描绘皇帝出巡狩猎、祭祀祖庙等活动,表达了对乾隆盛世的颂扬之情。《清实录》载:“乾隆五十四年己酉,八月己巳,上自避暑山庄启銮幸木兰。”[7]391管世铭有《己酉仲秋扈跸秋猎》,诗云:
(一)受制于学时的限制,只能节选部分统计方法进行讲解,但讲解方法明显偏少或代表性不足,不利用学生统计思维的全面塑造。
以上贵州喀斯特石林有关资料,源自于贵州山水旅游资源勘察开发设计院的有关研究成果,是创建“多彩贵州风,山地公园省”的重要科学依据。
3.2 黄连幼苗期的屏障结构
黄连子叶期质外体屏障结构,种子根和不定根为具凯氏带,木质化和强烈栓质化的内皮层和外皮层;胚轴为具凯氏带,木质化和强烈栓质化的内皮层,表皮外角质层。真叶期不定根仍具内皮层和外皮层,胚轴具内皮层和外侧木质化皮下层和角质层。黄连幼苗期不定根屏障结构为木质化和强烈栓质化的内皮层和外皮层;胚轴具木质化和强烈栓质化的内皮层,外侧具皮下层和角质层。
黄连不定根内皮层和外皮层的组织化学特征与陆生植物茶[20]的相似,内皮层和外皮层都具凯氏带且强烈栓质化,但弱木质化,外皮层木质化比内皮层略强烈。湿地植物如双穗雀稗(Paspalumdistichum)等[14,15,21],内皮层和外皮层都具凯氏带且强烈栓质化和木质化。胚轴木质化皮下层与水芹不定根木质化皮下层类似[22],而花椰菜(Brassicaoleracea)的不定根木质化皮层具一定程度忍耐水和离子的胁迫能力[23,24]。
本研究实验材料栽培过程中,子叶出土,随后分枝在地面以上,分枝着生不定根易干枯,不能吸收水分和矿质而影响黄连生长,产量低,故幼苗需要移栽,将第2年及以后的分枝需埋入土中。黄连不定根距根尖5mm,外皮层即有凯氏带和强烈栓质化,阻碍根尖吸收水分和矿质,即根尖吸收面积很小,这可能是其生长缓慢和适应阴生环境的原因之一;建议苗床土壤疏松,在移苗时尽量不损伤根系,以提高成活率。
致谢:感谢湖北省利川市农业局和利川市箭竹溪黄连科技开发有限公司的支持。
[1]赵楠,李隆云,白志川.中药材黄连的研究现状与展望[J].重庆理工大学学报(自然科学版),2015,29(1):53~58.
[2]Kamath S,Skeels M,Pai A.Significant difference in alkaloid content ofCoptischinensis(Huanglian),from its related American species[J].Chinese Medicine,2009,17 (4):1~4.
[3]王宪楷,杨培全,陈新民.黄连研究第五报,石柱黄连根茎中生物碱的研究[J].药学学报,1964,11(6):389~392.
[4]杨建民,唐红光.黄连(CoptischinensisFranch)根和根状茎的解剖[J].武汉植物学研究,1994,14 (3):164~168.
[5]袁王俊,张维瑞,尚富德.黄连营养器官解剖结构与其阴生环境相关性研究[J].河南大学学报(自然科学版),2007,37(2):184~186.
[6]王宪楷,李美蓉,涂茂利.川产黄连的形态组织及组织中小蘖碱分布的研究[J].四川医学院学报,1959,(1):1~15.
[7]鲁高莲,胡正海.黄连根茎中小蘖碱积累的组织化学研究[J].西北植物学报,1994,14(3):164~168.
[8]Greathouse G A,Watkins G M.Berberine as a factor in the resistance ofMahoniatrifoliolataandM.swaseyato phymatotrichum root[J].American Journal of Botany,1938,25:743~748.
[9]Cromwell B T.Experiments on the Origin and Function of Berberine inBerberisdarwinii[J].Biochemical Journal,1933,27:863.
[10]杨朝东,张霞,刘国锋,等.植物根中质外体屏障结构和生理功能研究进展[J].植物研究,2013,33(1):114~119.
[11]Jensen W A.Botanical histochemistry-principles and practice[M].San Francisco,CA:Freeman W H,1962:1~117.
[12]Brundrett M C,Enstone D E,Peterson C A.A berberine-aniline blue fluorescent staining procedure for suberin,lignin and callose in plant tissue[J].Protoplasma,1988,146:133~142.
[13]Brundrett M C,Kendrick B,Peterson C A.Efficient lipid staining in plant material with Sudan red 7B or Fluorol yellow 088 in polyethylene glycol-glycerol[J].Biotechnic and Histochemistry,1991,66:111~116.
[14]杨朝东,张霞.双穗雀稗(Paspalumdistichum) 通透性生理和茎解剖结构补充研究[J].植物研究,2013,33(5):564~568.
[15]Yang C,Zhang X,Li J,etal.Anatomy and histochemistry of roots and shoots in wild Rice (ZizanialatifoliaGriseb.) [J].Journal of Botany,2014,Article ID 181727.
[16]Watanabe H,Saigusa M,Morita S.Identification of Casparian bands in the mesocotyl and lower internodes of rice (OryzasativaL.) seedlings using fluorescence microscopy[J].Plant Production Science,2006,9:390~394.
[17]Vecchia F D,Cuccato F,Rocca N L,etal.Endodermis-like sheaths in the submerged freshwater macrophyteRanunculustrichophyllusChaix[J].Annals of Botany,1999,83:93~97.
[18]Mc Manus H A,Seago Jr J L,Marsh L C.Epifluorescent and histochemical aspects of shoot anatomy ofTyphalatifoliaL.,TyphaangustifoliaL.andTyphaglaucaGodr[J].Annals of Botany,2002,90:489~493.
[19]杨朝东,李守峰,姚兰,等.天胡荽(Hydrocotylesibthorpioides)的解剖和屏障结构特征研究[J].草业学报,2015,24(7):139~145.
[20]胡露洁,杨朝东,袁龙义,等.湖北‘利川红’茶不定根的解剖和屏障结构时空发育特征研究[J].湖北农业科学,2016,55(7):3662~ 3665.
[21]Yang C D,Zhang X,Zhou C Y,etal.Root and stem anatomy and histochemistry of four grasses from the Jianghan floodplain along the Yangtze River,China[J].Flora,2011,206:653~661.
[22]张霞,胡露洁,周存宇,等.水芹(Oenanthejavanica)适应湿地环境的解剖和屏障结构特征研究[J].中国蔬菜,2016,(7):52~58.
[23]López-Pérez L,Fernández-García N,Olmos E,etal.The phi thickening in roots of broccoli plants.An adaptation mechanism to salinity[J].International Journal of Plant Sciences,2007,168:1141~1149.
[24]Fernandez-Garcia N,Lopez-Perez L,Hernandez M,etal.Role of phi cells and the endodermis under salt stress inBrassicaoleracea[J].New Phytologist,2009,181:347~360.
[编辑] 余文斌
2016-12-02
湿地生态与农业利用教育部工程研究中心(长江大学)开放基金项目(KF201603)。
杨再辉(1976-),男,农艺师,现从事药用植物研究。通信作者:杨朝东,chaodongyang@aliyun.com。
Q944.54;Q949.746.5
A
1673-1409(2017)06-0054-05
[引著格式]杨再辉,周存宇,杨朝东.黄连幼苗期的解剖结构研究[J].长江大学学报(自科版),2017,14(6):54~58.
猜你喜欢
作物研究(2022年1期)2022-11-27
作物学报(2022年10期)2022-07-21
现代农业研究(2021年10期)2021-11-02
上海农业学报(2021年4期)2021-08-28
植物研究(2021年2期)2021-02-26
中国稻米(2019年6期)2019-12-05
天然产物研究与开发(2018年4期)2018-05-07
上海农业学报(2017年4期)2017-04-10
江西农业学报(2017年12期)2017-02-03
天津农业科学(2015年11期)2015-12-03
