
干湿度梯度及植物生活型对土壤氮磷空间特征的影响①
2017-05-15 07:38卢同平张文翔武梦娟林永静
土壤 2017年2期
卢同平,张文翔*,武梦娟,林永静
(1 云南师范大学高原湖泊生态与全球变化实验室,昆明 650500;2 云南师范大学旅游与地理科学学院,昆明 650500;3 云南师范大学云南省高原地理过程与环境变化重点实验室,昆明 650098)
干湿度梯度及植物生活型对土壤氮磷空间特征的影响①
卢同平1,2,3,张文翔1,2,3*,武梦娟1,2,3,林永静1,2,3
(1 云南师范大学高原湖泊生态与全球变化实验室,昆明 650500;2 云南师范大学旅游与地理科学学院,昆明 650500;3 云南师范大学云南省高原地理过程与环境变化重点实验室,昆明 650098)
通过对我国土壤氮磷含量及化学计量比值的分布特征研究,探讨了湿度梯度与植物生活型对土壤氮磷空间分布的影响。研究结果表明:中国陆地系统自然土壤氮、磷含量及氮磷比分别在0.02 ~ 8.78 g/kg,0.05 ~ 1.73 g/kg和0.06 ~ 9.85范围内变化,地形和气候因素对其空间变异性的影响均较为显著。干湿度梯度分带对土壤氮含量和氮磷比值的影响较磷含量更为明显,氮含量与氮磷比呈现出明显的梯度变化特征,表现出湿润区>半湿润区>干旱区>半干旱区的规律,而磷含量变化不显著。在不同生活型植被土壤中,森林土壤氮磷的变化规律较灌木和草本土壤更为复杂;其植物磷平均含量低于全国平均值,说明森林生态系统植物受磷元素的限制作用与其较低的土壤磷含量供给有关。土壤氮磷含量和氮磷比与年均降水量和年均温度的相关性及逐步回归分析表明,土壤氮磷及其化学计量的变异性主要是受降水的影响。
干湿度梯度;土壤氮磷;植物生活型;化学计量;回归分析
生态化学计量学是一门研究有机体主要组成元素碳(C)、氮(N)、磷(P)的养分化学计量关系的新科学[1–2]。而 N、P作为生物体遗传物质和蛋白质合成的主要生命元素,对生物体生长、发育、代谢等生命活动具有重要意义[3],同时也是自然陆地生态系统中主要的限制性元素,它们之间既相互独立,又相互作用[4],N∶P不仅是探讨植物生态系统结构和功能的重要指标[5],又是判断土壤对植物养分供应状况的指标[6–8]。土壤对植物的生长代谢提供必要的养分支持,在生物地球化学循环中扮演着十分重要的角色,其含量和分布状况对植物的生长发育产生着重要影响[9]。
自生态化学计量学的应用研究在国内外展开以来,运用生态化学计量学的方法对区域环境尺度上植物器官和土壤养分元素与地理环境因子、气候因子及人类干扰等化学计量特征的研究已取得了一系列成果[5,10–15],但研究结果存在差异。Reich和 Oleksyn等[16]在总结全球452个样点1 280种植物5 087个叶片的N、P分布格局时发现,随着纬度的增加,叶片N、P含量也在升高,而N∶P下降,且植物叶片及N∶P与土壤养分的有效利用紧密相关;He等[17]对阿拉善荒漠干旱区植物叶片与土壤的N、P及化学计量研究发现土壤全氮和全磷呈显著相关关系,并且土壤N、P随土壤层深度加深而减少,但土壤总N∶P并没有随深度变化而呈现出显著差异;而Manuel等[18]对全球干旱区土壤养分循环的研究显示,随着全球变化,干旱的加剧将降低干旱区土壤的 N浓度,但土壤P浓度会增加。区域尺度土壤N、P及化学计量的研究对于认识陆地生态系统空间格局的变化规律、未来变化趋势的预测及对全球变化的区域响应具有重要意义。然而到目前为止,对中国区域空间尺度上土壤N、P含量及化学计量特征的研究报道较少,尤其以地理和气候因素为空间界限划分的区域性研究更是很少。本研究以此研究格局为出发点,通过收集公开发表的大量文献资料,整理归类,提取关于研究土壤养分元素的文献数据,通过研究分析试图揭示以下几个问题:中国区域干湿度梯度(干旱、半干旱、半湿润、湿润)下土壤 N、P含量及化学计量的空间分布的特征;不同生活型植被下土壤N、P的养分状况的变化规律;气候因子(主要为降水和温度)与干湿度梯度下的土壤N、P的相关关系及影响土壤N、P空间变异的根本原因。本研究针对以上问题展开分析讨论,旨在丰富我国不同生活型土壤N、P含量及化学计量的研究以及在解决这些问题的同时为我国土壤养分研究和植被保护与恢复提供基础依据,同时以期能进一步推动我国该领域研究的广度和深度。
1 材料与方法
研究以陆生系统土壤为研究对象,从收集的数据所占空间区域分布将其归类于中国干湿度梯度分布的4个带,即干旱区、半干旱区、半湿润区和湿润区(以降水界限划分)。研究土壤为各带内受人类干扰较小的自然土壤,即荒漠草本土壤、草地土壤、灌木土壤和森林土壤。根据文献描述及中国土壤类型划分统计出研究样地土壤类型为:干旱区以灰漠土、灰棕漠土、棕漠土、棕钙土、栗钙土、风沙土、高山/亚高山草原土为主;半干旱区为褐土、棕钙土、盐碱土;半湿润区为棕钙土、草甸土、灰色森林土、黑垆土;湿润区为紫色土、黄壤、棕黄壤、红壤、砖红壤性红壤、潮土。土壤的植物生活型为草本、灌木和森林。最后归类出4个带内的研究样地分别为23、67、204、310个,总样地604个。
本研究数据主要通过收集公开发表的文献资料获得,主要渠道是以土壤氮(N)磷(P)、土壤养分和生态化学计量为关键词,通过CNKI数据库、万方数据库、维普期刊数据库及 Springer 电子期刊数据库获得相关文献数据,然后以文章是否满足研究土壤和植被N、P及N∶P为条件,剔除一些研究领域相关性较低、控制试验研究和耕作土壤研究等不符合条件的文献数据,最后整理所得研究所需文献数据。气候因子数据来自于原始文献数据,如果原始文献没有给出则通过其他文献查找或通过原始文献提及的经纬度及海拔等地理因子取研究样地所在中心位置气候因子作为参考。在对所获取的数据整理分类后,利用相关性分析和回归分析对其进行变量数据统计分析,并在统计前对数据进行正态分布检验,检验结果表明N、P含量及N∶P均呈正态分布。本文中土壤N(即全氮)、P(即全磷)含量及N∶P数据均为土壤0 ~ 20 cm层。
采用One-Way ANOVA进行不同干湿度梯度和不同生活型植物土壤N、P及N∶P的差异性比较,用最小显著差数法(LSD)进行显著差异性多重比较;相关性分析采用Spearman相关性分析法,土壤N、P与气候因子和元素间的回归分析采用多元线性逐步回归分析法。统计分析运用SPSS 19.0完成。
2 结果与分析
2.1 干湿度梯度下土壤氮磷含量及其化学计量特征
方差分析显示,在干湿度梯度下,土壤N、P含量及N∶P在干旱、半干旱、半湿润、湿润区的空间变异性存在较大差异(P<0.05):干旱区N含量与半干旱、半湿润和湿润区相比,存在显著差异性,其变异系数分别为 0.14、0.92、0.81、0.98,变异性最大者为湿润区(0.02 ~ 8.78 g/kg),而最小的干旱区为0.7 ~0.85 g/kg;但是N含量平均值最小的却是半干旱区,为0.51 g/kg,与土壤矿化速率和森林面积及氮沉降区域程度不同等有关,土壤N含量最大出现在湿润区。而且比较发现,土壤N含量的变化关系为湿润区(2.71 g/kg)>半湿润区(1.23 g/kg)>干旱区(0.77g/kg)>半干旱区(0.51 g/kg)(图1A)。此外,统计得出,全国自然土壤N平均值为1.41 g/kg。
土壤P含量的变化在干湿度带间无明显规律性。变异性最大的为干旱区和湿润区,变异系数为 0.84和0.69(图1B),半干旱和半湿润区则变异系数较小,为0.37和0.23。土壤P含量最大值与N含量类似,湿润区为1.73 g/kg,但最小值同样出现在该区域,为0.13 g/kg,平均值则过渡带的半湿润区最大(0.95 g/kg),半干旱区最小(0.54 g/kg),干旱区(0.70 g/kg)高于湿润区(0.61 g/kg),全国土壤P的平均值为0.66 g/kg。土壤P含量总体表现为半湿润区>干旱区>湿润区>半干旱区,说明我国半干旱区和湿润区土壤P含量低于全国平均水平。
N∶P的空间变异性与土壤N和P含量的变化密切相关,方差分析显示,干湿度对土壤 N∶P的影响极为显著(P<0.001)。与土壤N、P含量不同的是,N∶P的空间变异性在半湿润区和湿润区都较大;与N含量相同,N∶P的最小值和最大值都出现在湿润区,分别为0.06和9.85;4个区的土壤N∶P关系为湿润区(4.24)>干旱区(2.02)>半湿润区(1.48)>半干旱区(1.23);全国土壤N∶P为2.79(图1C)。
综上所述,我国干旱和半干旱区域土壤N含量低于全国平均水平,而P含量则湿润区和半干旱区低于全国平均水平,且生态脆弱区的半干旱区域是N、P都比较贫瘠区。方差分析显示:干湿度梯度对土壤 N含量的影响(P<0.01)要显著于土壤 P含量(P<0.05)。
2.2 不同生活型植物下土壤氮磷含量及其化学计量特征
对不同生活型植物的土壤N、P含量及N∶P的方差分析显示:不同生活型对土壤 N含量的影响大
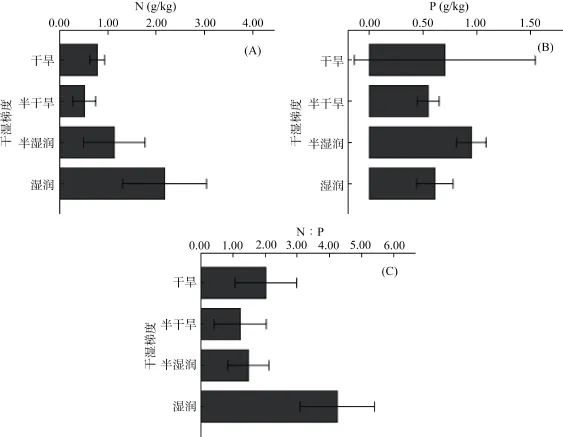
图1 干湿度梯度下土壤N、P含量及N∶P空间变化比较Fig. 1 Spatial variations of soil total nitrogen, total phosphorus contents and N∶P mass ratio under humidity/aridit gradient in China
于土壤P含量(P<0.05)。如图2所示,不同生活型植物土壤N含量的平均值以森林土壤最大(2.34 g/kg),灌木土壤最小(0.90 g/kg),但土壤P含量在3种生活型植被下的差异并不大(0.69、0.67、0.60 g/kg)。而森林土壤N、P含量变异性最大,草本土壤最小,且二者的最大值分别为森林土壤和灌木土壤,分别为8.78 g/kg和1.73 g/kg,但最小值分别为灌木和草本土壤,分别为0.02 g/kg和0.05 g/kg。

图2 不同生活型植物下土壤N、P含量(g/kg)及N∶P均值比较Fig. 2 Mean value comparison of soil N, P contents and N∶P mass ratio under different vegetation forms
不同生活型植物的土壤N、P含量及N∶P的差异不同,生活型间和同种生活型植物的变异性也存在差异。在草本、灌木和森林植物土壤之间,其N、P含量及 N∶P受生活型影响最为显著的是 N含量(P<0.001),其次为P含量和N∶P(P<0.05, P<0.05)。而在同种生活型植物内,土壤N含量及N∶P的变异性大小表现为灌木>草本>森林,变异系数分别为1.52、1.38、0.93和 1.33、1.09、0.56。但土壤P含量的变异性则表现为森林>灌木>草本,变异系数为0.67、0.55和0.49。
不同生活型植物土壤的N、P含量及N∶P与全国自然土壤平均值相比,灌木和草本 N含量均低于全国平均水平(1.41 g/kg)草本土壤 N含量平均值为1.15 g/kg;而 P含量则森林土壤低于全国平均水平(0.66 g/kg)。这说明我国以草本和灌木为主的区域植物对土壤N含量的吸收需求可能受到限制,而森林植被类型的植物群落可能受土壤P含量的限制较为明显。
2.3 土壤氮磷含量及其化学计量比与气候因子间的相关与回归分析
2.3.1 土壤氮磷含量及其化学计量比与气候因子间的相关性 相关性分析发现,土壤 N、P含量及N∶P均与年均降水存在显著相关关系,其中与N含量(rs=0.687,P=0.002)和N∶P(rs=0.550,P<0.001)为显著正相关,与 P含量呈显著负相关(rs=–0.410, P= 0.003);土壤N含量及N∶P与年均温度呈显著正相关,但P含量与年均温度无明显相关性;土壤N含量与P含量之间无明显的相关性,而N∶P则与N含量和P含量都存在显著相关性关系,与N含量呈显著正相关,与P含量呈显著负相关,但与N含量的显著性(rs=0.767, P<0.001)要高于 P含量(rs=–0.279,P<0.05)(表1,图3)。

表1 土壤N、P含量及N∶P与年均降水和年均温度间的Spearman相关性分析rs值Table 1 Spearman correlation coefficients (rs) among soil N, P contents, N∶P, mean annual precipitation and temperature
2.3.2 土壤氮磷含量及其化学计量比与气候因子间的回归分析 对土壤 N含量进行多元线性回归,以土壤N为因变量,土壤P含量和年均降水及年均温度为自变量,在α=0.05水平上进行多元线性逐步回归分析,见表2。其中年平均温度与土壤N含量无明显因果关系,因此未进入方程,回归方程为:YN=–0.908 + 2.056X1+ 0.001X2,R2= 0.228;其中,YN代表土壤N含量,X1代表P含量,X2代表年均降水。由此可以得出,土壤N含量主要受土壤P含量和年均降水的的影响,二者决定系数分别为 2.056和0.001,说明P含量的影响要高与年均降水。
同样,对土壤P含量进行多元逐步回归分析,以土壤P含量为因变量,土壤N含量、年均降水和年均温度为自变量,回归方法与土壤 N含量类似,结果显示,土壤P含量与年均温度无明显归因关系,与土壤N含量和年均降水归因关系较为明显(表2),回归方程为:YP= 0.769 – 0.0000324X1+ 0.086X2,R2= 0.273;其中,YP代表土壤P含量,X1代表年均降水,X2代表土壤N含量。由此可以得出,土壤P含量受年均温度的影响不明显(未进入方程),而主要受年均降水和土壤N含量的影响,决定系数分别为–0.0000324和0.086,即土壤P含量与年均降水呈反比关系,与土壤N含量呈正比关系,且与土壤P含量的相关关系要强于与年均降水的相关性。

表2 土壤N、P含量的多元线性逐步回归分析结果Table 2 Multiple linear stepwise regression of soil N and P contents
土壤N∶P比率受N、P含量的共同影响,但由于土壤N、P含量的差异较大,二者对其的影响程度也会不同。因此,通过 N∶P回归分析发现,N∶P与土壤N含量具有高度显著正相关关系(P<0.001),其值的变化主要由N元素决定;与土壤P含量虽呈负相关关系,但相关性较弱(P>0.05),说明P含量对N∶P的决定因素要远小于土壤N含量(图3,图4)。
3 讨论
3.1 土壤氮磷含量及其化学计量化与干湿度梯度和植物生活型的关系
植物地上部分元素特征通常与土壤养分含量具有一定的相关性[19–21]。温度变化对植物叶片N、P含量影响的研究结论比较一致:随着温度的升高,叶片N、P含量降低;而水分变化对植物叶片影响的研究报道较少,目前的结论是N、P含量随着经度的增加而增加[22]。本文综合温度与降水两个因素得出,土壤N含量及N∶P与年均温度显著正相关,但P含量与温度的相关性并不明显;且与降水的相关分析表明,土壤N含量和N∶P与年均降水间的相关性与温度的类似,不同的是土壤P含量随着年均降水的增加而减少,这与植物叶片的研究结果具有同样的变化规律[23]。本研究结果得出,干旱区的土壤N、P含量均值在4个带区中都不是最值,但N含量和 N∶P的变异系数最小,P含量的变异系数最大,而最值都发生在湿润区,其原因可能是干旱的气候条件下干旱区植物对土壤N、P的吸收效率具有重要影响[24],而且 P含量的变异系数虽大,但其含量在草地土壤和荒漠土壤中差异并不大,因此导致了N∶P的变异系数最小;湿润区N含量最高P含量最小的可能原因是其主要来源于矿物质分解和漫长的岩石风化过程[8],森林土壤有较为丰富的腐殖质层,腐殖质含量较高,进而导致 N含量明显高于其他 3个带区,P含量最小是因为湿润区较高的降水量增加了土壤的淋溶作用,可能导致本来就较低的P含量随水分发生了元素迁移,从而使得湿润区的植物叶片P含量也较低;根据N∶P的大小反映土壤对植物养分供应的能力,表明湿润区土壤较低的P含量导致了植物叶片受P的限制。土壤P含量的最大值在半湿润区,可能与在半湿润区分布有较为广泛的草地系统有关。
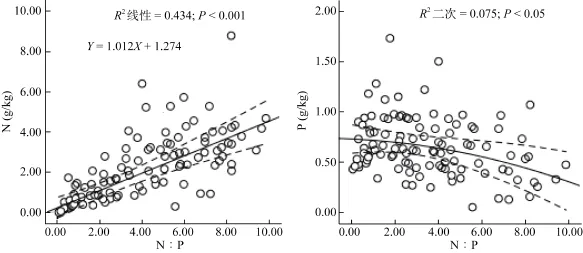
图4 土壤N、P含量与N∶P间的相关回归关系Fig. 4 Correlation regression among soil N, P contents and N∶P
本研究得出不同生活型植被类型对土壤N、P含量都存在重要影响,但对土壤N的影响大于P,而且森林土壤 N含量要明显高于其他生活型植被类型土壤,这与前人对黄土丘陵沟壑区不同植被区土壤研究得出土壤养分含量受植被类型及植被盖度的影响,而且森林区要大于其他植被区的部分结果一致[25]。本文结果中得出森林区土壤 N含量和 N∶P 最高,P含量最低(图2),这不仅反映了土壤N、P元素的来源及不同的土壤基质状况影响土壤养分含量,而且说明了土壤不同厚度腐殖质层和不同强度的年降雨量影响了土壤N、P含量及N∶P的格局,同时,也使得森林植物群落受养分限制的可能性增大,尤其在演替过程中这种限制作用更加明显[26–27],而且这种限制可能导致森林群落由丰富复杂向耐干旱和贫瘠的群落类型发展[28]。
3.2 土壤氮磷含量及其化学计量比与气候因子的相关与回归分析
本研究通过对影响土壤N、P含量的气候因子和元素间相互作用(N、P之间)的相关分析和回归分析发现,土壤N、P含量与年均降水和年均温度均存在显著相关性,但元素间的相关性不明显,但回归分析却发现土壤 N含量与年均温度无明显归因关系,而与土壤P含量和年均降水量具有明显相关性,土壤P含量也存在这种情况。其主要原因可能是土壤微生物繁育代谢时对养分含量的敏感性调控了这一关系,土壤与微生物之间复杂的交互作用为植物生长发育提供了营养保障[29],因为土壤微生物受不同植被非均质性土壤的影响十分显著[30],而且土壤微生物在受土壤基质、土壤养分状况和土壤元素化学计量比影响的同时也调控着土壤重要养分元素(C、N、P、K)的生物转化和土壤的理化结构[31–32]。然而,这方面的研究以后应该以时间尺度和植物–凋落物–土壤作为一个循环系统进行跟踪研究[33–34],同时从微观分子角度深入研究土壤微生物与土壤的关系,这能使整个生物地球化学循环的研究更深入本质化。
4 结论
中国土壤的N、P含量及N∶P的空间变异性均较高(P<0.05; P<0.05; P<0.01);在干湿度梯度下土壤N含量与N∶P的空间变化具有明显的梯度特征,表现为湿润区最高,半干旱区最小,总体变化趋势为湿润区>半湿润区>干旱区>半干旱区;土壤P含量在干湿梯度下无明显的变化规律,表现为半湿润区最高,半干旱区最低,但在湿润区的变异性较大;土壤N、P含量及化学计量特征的空间变化与植物的不同生活型有关,森林土壤与灌木和草本土壤相比,其N、P含量及N∶P与均表现出明显的变化特征,且森林土壤植物受P元素的限制作用极为明显。土壤N、P含量及 N∶P与年均降水均具有显著相关性(P<0.05),N含量和 N∶P与年均温度显著正相关(P<0.05),但P含量与年均温度无明显相关性(P<0.05);回归分析与相关分析结果不同,土壤N、P含量均与年均温度无显著的因果关系,而与年均降水和元素间的相互作用具有较为显著的相关性。
[1] Elser J J, Dobberfuhl D R, Mackay N A, et al. Organism size, life history, and N:P stoichiome[J]. BioScience, 1996, 46(9): 674–684
[2] 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索[J]. 植物生态学报, 2005, 29(6): 1007–1019
[3] Elser J J, Sterner R W, Gorokhova E, et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000b, 3, 540–550
[4] Han W X, Fang J Y, Guo D, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377–385
[5] Elser J J, Fagan W F, Denno R F, et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 2000, 408(6812): 578–580
[6] Wassen M J, Olde V, Harry G M, et al. Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems[J]. Journal of Vegetation Science, 1995, 6(1): 5–16
[7] Aerts R, Chapin F S Ⅲ. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns[J]. Advances in Ecological Research, 2000, 30: 1–67
[8] 刘兴诏, 周国逸, 张德强, 等. 南亚热带森林不同演替阶段植物与土壤中 N、P 的化学计量特征[J]. 植物生态学报, 2010, 34(1): 64–71
[9] 刘兴华, 陈为峰, 段存国, 等. 黄河三角洲未利用地开发对植物与土壤碳、氮、磷化学计量特征的影响[J]. 水土保持, 2013, 27(2): 204–208
[10] Thompson K, Parkinson J A, Band S R, et al. A comparative study of leaf nutrient concentrations in a regional herbaceous flora[J]. New Phytologist, 1997, 136(4): 679–689
[11] Reich P B, Ellsworth D S, Walters M B, et al. Generality of leaf trait relationships: a test across six biomes[J]. Ecology, 1999, 80(6): 1955–1969
[12] 郑淑霞, 上官周平. 黄土高原地区植物叶片养分组成的空间分布格局[J]. 自然科学进展, 2006, 16(8): 965–973
[13] He J S, Fang J Y, Wang Z H, et al. Stoichiometry and largescale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006, 149(1): 115–122
[14] Braakhekke W G, Hooftman D A P. The resource balance hypothesis of plant species diversity in grassland[J]. Journal of Vegetation Science, 2009, 10( 2): 187–200
[15] 杨文, 周脚根, 王美慧, 等. 亚热带丘陵小流域土壤碳氮磷生态计量特征的空间分异性[J]. 土壤学报, 2015, 52(6): 1336–1344
[16] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences, 2004, 101(30): 11001–11006
[17] He M Z, Dijkstra F A, Zhang K, et al. Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability[J]. Scientific Reports, 2014, 6392(4): 1–7
[18] Manuel D B, Maestre F T, Gallardo A, et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands[J]. Nature, 2013, 502(31): 672–676
[19] Aerts R, Chapin F S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30: 1–67
[20] Hedin L O. Global organization of terrestrial plant-nutrient interactions. Proceedings of the National Academy ofSciences of the United States of America[J], 2004, 101( 30): 10849–10850
[21] Liu X Z, Zhou G Y, Zhang D Q, et al. N and P stoichiometry of plant and soil in lower subtropical forest successional series in southern China[J]. Acta Phytoecologica Sinica, 2010, 34( 1): 64–71
[22] 丁小慧, 罗淑政, 刘金魏, 等. 呼伦贝尔草地植物群落与土壤化学计量特征沿经度梯度变化[J]. 生态学报, 2012, 32(11): 3467–3476
[23] 任书杰, 于贵瑞, 陶波, 等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 2007, 28(12): 2665–2673
[24] Seligman N G, Sinclair T R. Global environment change and simulated forage quality of wheat II. Water and nitrogen stress[J]. Field Crops Research, 1995, 40(1): 29–37
[25] 朱秋莲, 邢肖毅, 张宏, 等. 黄土丘陵沟壑区不同植被区生态化学计量特征[J]. 生态学报, 2013, 33(15): 4674–4682
[26] 陈伏生, 曾德慧, 何兴元. 森林土壤氮素的转化与循环[J].生态学杂志, 2004, 23(5): 126–133
[27] 庞圣江, 张培, 贾宏炎, 等. 桂西北不同森林类型土壤生态化学计量特征[J]. 中国农学通报, 2015, 31(1): 17–23
[28] Gang W, Jing W, Hong B D, et al. Nutrient cycling in an Alpine tundra ecosystem on Changbai Mountain,Northeast China[J]. Applied Soil Ecology, 2006, 32(2): 199–209
[29] 雷海迪, 尹云锋, 刘岩, 等. 杉木凋落物及其生物炭对土壤微生物群落结构的影响[J]. 土壤学报, 2016, 53(3): 790–799
[30] Mclntosh A C S, Macdonald S E, Quideau S A. Linkages between the forest floor microbial community and resource heterogeneity within mature lodgepole pine forests[J]. Soil Biology and Biochemistry, 2013, 63: 61–72
[31] 马瑞丰, 刘金福, 张广帅, 等. 格氏栲林土壤生态化学计量和微生物群落特征及其关联性分析[J]. 植物资源与环境学报, 2015, 24(1): 19–27
[32] 党亚爱, 王立青, 张敏. 黄土高原南北主要类型土壤氮组分相关关系研究[J]. 土壤, 2015, 47(3): 490–495
[33] 卢同平, 史正涛, 牛洁, 等. 我国陆生生态化学计量学应用研究进展与展望[J]. 土壤, 2016, 48(1): 29–35
[34] 孔涛, 张德胜, 寇涌苹, 等. 浑河上游典型植被河岸带土壤有机碳、全氮和全磷分布特征[J]. 土壤, 2014, 46(5): 793–798
Effects of Humidity/Aridity Gradient and Vegetation Form on Spatial Characteristics of Soil Nitrogen and Phosphorus in China
LU Tongping1,2,3, ZHANG Wenxiang1,2,3*, WU Mengjuan1,2,3, LIN Yongjing1,2,3
(1 Key Laboratory of Plateau Lake Ecology and Global Change, Yunnan Normal University, Kunming 650500, China; 2 College of Tourism and Geography Science, Yunnan Normal University, Kunming 650500, China; 3 Key Laboratory of Plateau Geographical Process and Environmental Change of Yunnan Province, Yunnan Normal University, Kunming 650098, China)
In this paper, the impact of humidity/aridity gradient and vegetation form on the spatial distribution on soil nitrogen (N) and phosphorus (P) contents and N∶P ratio in China were discussed. The results showed that N and P contents and N∶P ratio of natural soil of terrestrial ecosystems in China were 0.02–8.78 g/kg, 0.05–1.73 g/kg and 0.06–9.85, respectively. The spatial variabilities of N and P contents and N∶P ratio were further significantly influenced by topography and climate. Humidity/ aridity gradient had greater effect on N content and N∶P than on P content. N content and N∶P showed obvious gradient change characteristics with an order of humid region > semi-humid region > arid region > semi-arid region, while P content change was not significant. The variations of N and P in forest soil were more complex than those in shrub and herb. The average content of plant phosphorus was lower than the national average, may indicating forest ecosystem limited by P is attributed to low soil P content. Correlation and stepwise regression analyses proved precipitation controls the variation of N and P contents and N∶P ratios.
Humidity/aridity gradient; Soil N and P; Vegetation form; Ecological stoichiometry; Regression analysis
P9351.1;P595;Q148
A
10.13758/j.cnki.tr.2017.02.023
国家自然科学基金项目(41461015)、云南省中青年学术技术带头人后备人才项目(2015HB029)和云南省水利厅项目(2014003)资助。
* 通讯作者(wenxiangzhang@gmail.com)
卢同平(1988—),男,甘肃定西人,硕士研究生,主要研究方向为环境地球化学。E-mail: tongpinglu2014@sina.com
猜你喜欢
农业灾害研究(2022年11期)2023-01-09
考试与评价·七年级版(2021年5期)2021-08-14
科学(2020年6期)2020-02-06
中央民族大学学报(自然科学版)(2018年2期)2018-11-09
儿童故事画报·自然探秘(2017年2期)2017-09-26
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
现代农业科技(2016年22期)2017-03-24
学周刊·下旬刊(2015年10期)2015-07-22
植物营养与肥料学报(2012年5期)2012-10-26
