
崖柏属植物的核型分析
2017-05-12 07:55:01倪妍妍杨文娟刘建锋江泽平
林业科学研究 2017年2期
倪妍妍, 杨文娟, 刘建锋, 江泽平
(中国林业科学研究院林业研究所, 国家林业局林木培育重点实验室, 北京 100091)
崖柏属植物的核型分析
倪妍妍, 杨文娟, 刘建锋*, 江泽平
(中国林业科学研究院林业研究所, 国家林业局林木培育重点实验室, 北京 100091)
[目的]为了从细胞学角度探讨崖柏属植物分类与进化问题,[方法]采用根尖压片法对崖柏属5种植物进行核型研究。[结果]表明:崖柏和日本香柏的核型公式为2n=2x=22=18m(2SAT)+4sm,朝鲜崖柏、北美乔柏和北美香柏的核型公式为2n=2x=22=20m(2SAT)+2sm,5种植物的核型均属于1A类型。[结论]该属在柏科中可能处于比较进步的进化地位;通过进化趋势图分析发现,崖柏在该属中分化最晚,而朝鲜崖柏则较原始。
核型;崖柏;朝鲜崖柏;日本香柏;北美乔柏;北美香柏
崖柏属(ThujaL.)为柏科(Cupressaceae)的一个小属群,属下仅有5个现存种,间断分布在东亚和北美东西部。东亚3个种分布范围均较狭窄,如崖柏(T.sutchuenensisFranch)为我国特有的极度濒危物种,仅见于渝东北的石灰岩山地(城口县和开县交界处);朝鲜崖柏(T.koraiensisNakai)分布于我国长白山地区和朝鲜半岛,日本香柏(T.standishiiL.)则为日本所特有。北美2个种分布较广,北美乔柏(T.plicataD. Don)分布可北至亚北极泰加林,南至落叶林区;北美香柏(T.occidentalisL.)分布于北美西部太平洋海岸山地与落基山脉[1]。
崖柏属植物的细胞学研究国内外已有一些报道[2-3],本文对属内5种植物(崖柏、朝鲜崖柏、日本香柏、北美乔柏和北美香柏)进行了核型比较分析,其中,崖柏和朝鲜崖柏的核型均为首次报道,旨在为崖柏属的系统发育研究及资源利用提供细胞学资料。
1 材料与方法

核型分析的项目、标准和方法参照李懋学等[4]的核型分析标准。臂比(AR)=按长臂/短臂;染色体长度比(LR)=最长染色体长度/最短染色体长度;核型不对称系数(As.K.C)=(长臂总长/全组染色体总长)×100%;染色体总形态百分比(TM)=(短臂总长/全组染色体总长)×100%;染色体相对长度指数(I.R.L)=染色体长度/全组染色体平均长度。
2 结果与分析
2.1 染色体及核型分析
分别从崖柏、朝鲜崖柏、日本香柏、北美乔柏和北美香柏的5个细胞分裂相确定体细胞染色体数目,5个树苗的体细胞染色体数目均为2n=22,染色体基数x=11,未见染色体非整倍性变异和加倍现象。核型特征和数据结果见图1和表1。

图1 崖柏属植物染色体形态Fig.1 Chromosome morphology in species of genus Thuja L.
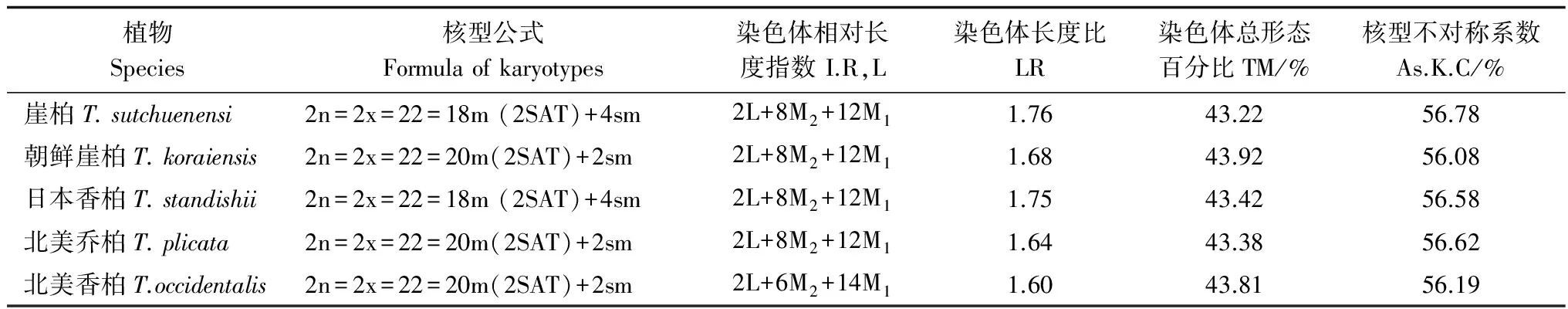
表1 5种崖柏属植物的核型参数
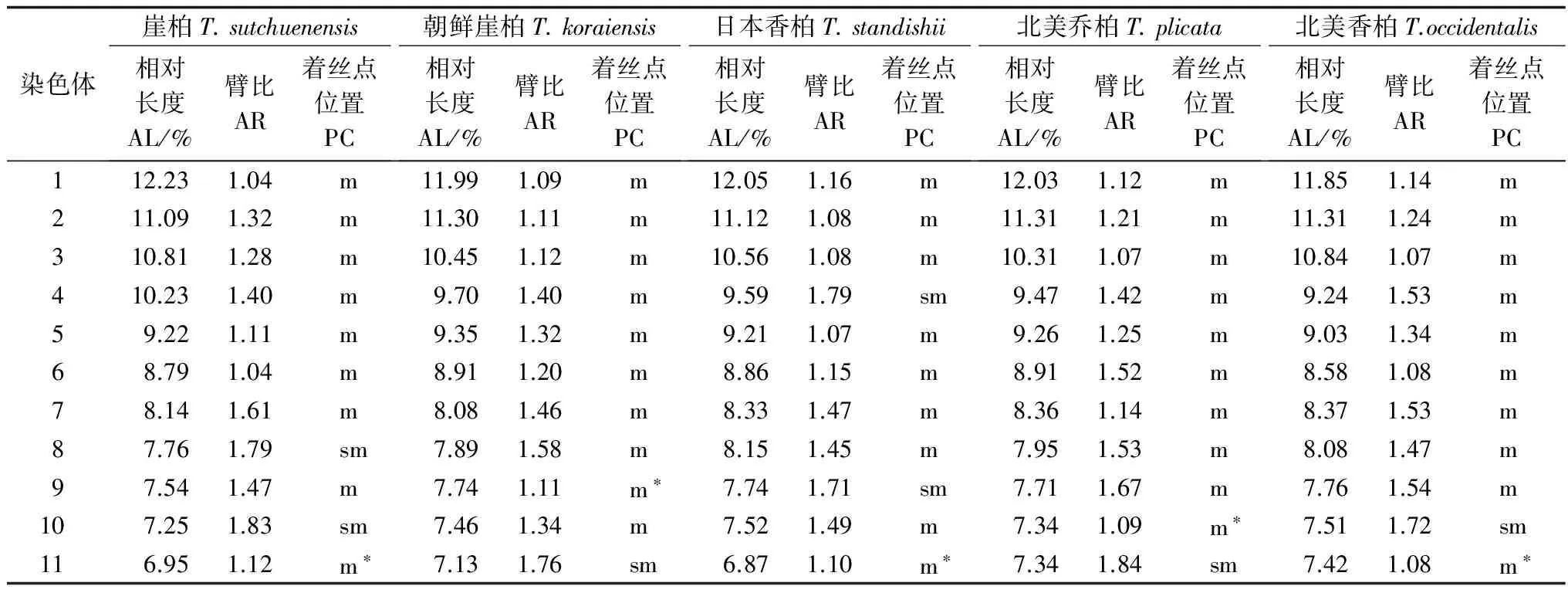
表2 5种崖柏属植物的染色体参数
注:*次缢痕, Note: secondary constriction.
结果显示:崖柏和日本香柏有9对中部着丝粒染色体和2对近中部着丝粒染色体,朝鲜崖柏、北美乔柏和北美香柏有10对中部着丝粒染色体和1对近中部着丝粒染色体,近中部着丝粒的数目不是完全相同,但平均臂比与染色体长度比比较接近,均没有臂比大于2的染色体。因此,按照Stebbins[5]的核型分类标准,崖柏属5种植物的核型均属于1A类型,这是一种最为整齐的对称核型。对崖柏属5种植物的染色体相对长度指数按Kuo等[6]提出的分类方法进行分类发现,染色体相对长度组成中除M1、M2染色体外都有1对L染色体;且均只具有一对随体,但不同种次缢痕的位置有所不同,如崖柏位于第11对染色体上,而朝鲜崖柏位于第9对染色体上。
2.2 崖柏属植物的进化趋势分析
平均臂比(MAR)和染色体长度比(LR)可以表示不同种间核型的不对称性,二者愈大,其核型愈不对称。从MAR和LR的二维进化趋势图(图2A)发现,崖柏和日本香柏沿MAR和LR方向进化较快,较为进化;而朝鲜崖柏和北美香柏则较为原始。
平均臂比(MAR)和核型不对称系数(As.K.C)则从另一个角度反映了核型的不对称性,二者愈大,核型愈不对称。从二维进化图(图2B)可以看出:崖柏属5种植物均按MAR和As.K.C向右上角方向进化,崖柏MAR及As.K.C均最大,表明其在崖柏属中分化最晚;相比之下,朝鲜崖柏则较为原始。
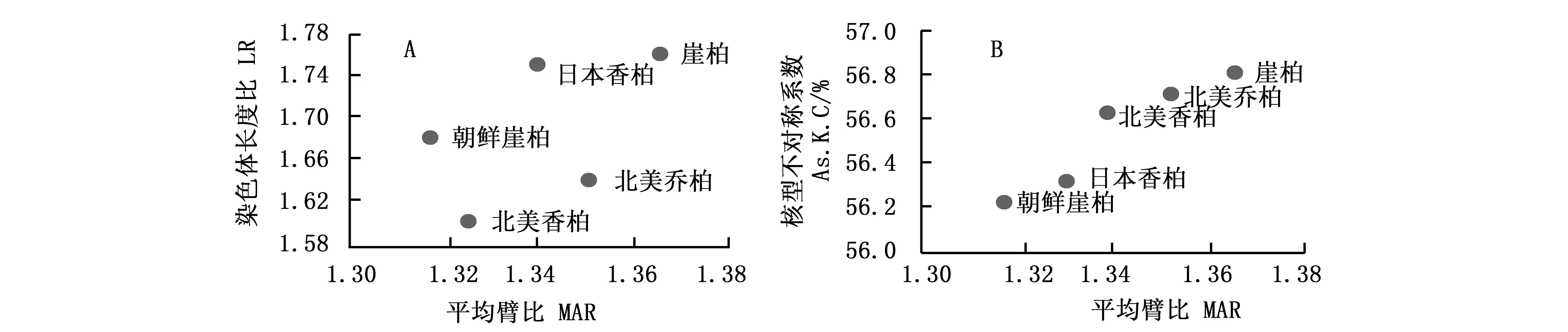
图2 崖柏属植物核型进化趋势图 (A:MAR vs LR;B:MAR vs As.K.C)Fig.2 Evolution trend map of karyotype in species of genus Thuja L.(A:Two-dimensional representation of evolutionary trend of MAR and LR; B:Two-dimensional representation of evolutionary trend of MAR and As.K.C)
3 讨论
李林初等[2]曾报道北美乔柏和北美香柏的核型公式分别为2n=2x=22=16m+6sm(2SAT)和2n=2x=22=20m(2SAT)+2sm;Kuroki等[3]曾报道日本香柏的核型公式为2n=2x=22=16m+6sm。其中,北美乔柏和日本香柏的核型公式与本文报道的结果不尽相同,这可能是由于取材、实验方法的处理不同或测量误差所致。
4 结论
本研究发现,崖柏和日本香柏的核型公式为2n=2x=22=18m(2SAT)+4sm,朝鲜崖柏、北美乔柏和北美香柏的核型公式为2n=2x=22=20m(2SAT)+2sm,5种植物的核型均属于1A类型。崖柏属处于柏科较进化的地位,而崖柏为崖柏属最后分化种。
[1] Eckenwalder J E. Conifers of the world: the complete reference[M]. Portland, United States:Timber Press,2009.
[2] 李林初,刘永强,王玉勤, 等. 侧柏亚科三种植物的核型及其细胞分类学研究[J]. 云南植物研究, 1996, 18(4): 439-444.
[3] Kuroki Y. Karyotype studies on important conifers[J]. Bull. Miyazaki Univ. Forests, 1969, 5: 1-103.
[4] 李懋学,陈瑞阳. 关于植物核型分析的标准化问题[J]. 植物科学学报, 1985, 3(4): 297-302.
[5] Stebbins G L. Chromosomal evolution in higher plants[M]. London: Edward Arnold, 1971.
[6] Kuo S R, Wang T T, Huang T C. Karyotype analysis of some formosan gymnosperms[J]. Taiwania, 1972, 17(1): 66-80.
[7] 郑 霞,高信芬,徐 波, 等. 七种木蓝属植物的染色体研究[J].植物科学学报,2011, 1(4): 417-422.
[8] Yang J, Wu J C, Gu Z J. Karyomorphology of three species inDipentodon(Dipentodontaceae),Perrottetia(Celastraceae), andTapiscia(Tapisciaceae) of the order Huerteales and their phylogenetic implications[J]. Journal of Systematics and Evolution, 2009, 47(4): 291-296.
[9] 李林初,钱 吉. 两种中国特有松属植物核型的研究兼论松属的系统位置[J]. 云南植物研究, 1993, 15(1): 47-56.
[10] 高和琼,庄南生,王 英, 等. 橡胶树两个品种的核型分析[J]. 植物科学学报, 2009, 27(5): 537-540.
[11] 邢世岩,高进红,姜岳忠, 等. 银杏特异种质核型进化趋势[J]. 林业科学, 2007, 43(1): 21-27.
[12] Levitzky G. The karyotype in systematics[J]. Bull Appl Bot Genet Plant Breed, 1931, 27: 220-240.
[13] Stebbins G L. Chromosomal variation and evolution[J]. Science, 1966, 152(3728): 1463-1469.
[14] 李林初,傅煜西. 柏木属的核型及细胞地理学研究[J]. 植物分类学报, 1996, 34(2): 117-123.
[15] 李林初,岑益群. 扁柏属的核型及柏木亚科的细胞分类学研究[J]. 云南植物研究, 1996, 18(1): 72-76.
[16] 马颖敏,邢世岩,王玉山, 等. 中国侧柏地理种源核型分析与进化趋势[J]. 分子植物育种, 2009, 7(6): 1186-1192.
[17] Mciver E E, Basinger J F .The morphology and relationships ofThujapolaris sp. nov.(Cupressaceae) from the early Tertiary, Ellesmere Island, Arctic Canada[J]. Canadian Journal of Botany, 1989, 67(6): 1903-1915.
[18] Farjon A. A bibliography of Cupressaceae and Sciadopitys[M]. Royal Botanic Gardens: Kew, England, 2005.
[19] Li J H, Xiang Q P. Phylogeny and biogeography ofThujaL.(Cupressaceae), an Eastern Asian and North American disjunct genus[J]. Journal of Integrative Plant Biology, 2005, 47(6): 651-659.
[20] Peng D, Wang X Q. Reticulate evolution inThujainferred from multiple gene sequences: implications for the study of biogeographical disjunction between eastern Asia and North America[J]. Molecular phylogenetics and evolution, 2008, 47(3): 1190-1202.
(责任编辑:詹春梅)
Karyomorphology of FiveThujaSpecies
NIYan-yan,YANGWen-juan,LIUJian-feng,JIANGZe-ping
(Research Institute of Forestry, Chinese Academy of Forestry, Key Laboratory of Tree Breeding and Cultivation, State Forestry Administration, Beijing 100091, China)
[Objective]To study the issues related to evolution and classification of the genusThujaL. [Method]Using the methods of toot tip squash and staining, the karyological characteristics of fiveThujaspecies were analyzed. [Result]The karyotype formulas ofT.sutchuenensisandT.standishiiare 2n=2x=22=18m(2SAT)+4sm, while the karyotype formula ofT.koraiensis,T.plicataandT.occidentalisare 2n=2x=22=20m(2SAT)+2sm, all belonging to 1A type. [Conclusion]It seems thatThujaL. is in a progressive evolution status in the family Cupressaceae. The evolutional trend analysis suggest thatT.sutchuenensisis the most advanced species, whileT.koraiensisis perhaps the most primitive species inThujaL.
karyotype;Thujasutchuenensis;Thujakoraiensis;Thujastandishii;Thujaplicata;Thujaoccidentalis
2015-07-14 基金项目: 国家林业局“948”项目(2013-4-47);中央公益性科研院所专项基金(RIF2010-15) 作者简介: 倪妍妍(1991—),女,硕士研究生,主要从事树木生理生态研究. * 通讯作者.E-mail:liujf@caf.ac.cn
10.13275/j.cnki.lykxyj.2017.02.001
S791
A
1001-1498(2017)02-0189-05
猜你喜欢
生物技术进展(2022年1期)2022-02-11 03:45:46
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
科学之谜(2018年9期)2018-12-17 09:05:08
艺术生活-福州大学厦门工艺美术学院学报(2016年6期)2016-11-01 21:55:41
生物学教学(2016年12期)2016-04-10 23:04:28
哈尔滨医药(2015年2期)2015-12-01 03:57:21
投资与理财(2015年15期)2015-09-10 06:27:07
大众投资指南(2015年6期)2015-05-30 10:48:04
中国当代医药(2015年9期)2015-03-01 02:02:12
遗传(2015年4期)2015-02-04 06:29:22
