
Bt基因导入对海岛棉农艺性状和产量性状影响的研究
2017-05-11 08:58贾莉莉吴鹏昊汪志星陈全家王希东曲延英
干旱地区农业研究 2017年2期
贾莉莉,吴鹏昊,李 娟,汪志星,陈全家,王希东,曲延英
(新疆农业大学, 新疆 乌鲁木齐 830052)
Bt基因导入对海岛棉农艺性状和产量性状影响的研究
贾莉莉,吴鹏昊,李 娟,汪志星,陈全家,王希东,曲延英
(新疆农业大学, 新疆 乌鲁木齐 830052)
对9个经PCR检测确定为转Bt基因阳性的海岛棉品系的农艺性状和产量性状考察结果表明,外源基因Bt的导入对9个海岛棉品系的农艺性状和产量性状有不同程度的影响。① 受体为K222时,Bt基因导入使得株高、始节高、始节数、有效果枝数、铃数、有效铃数差异达到极显著水平(P<0.01)。当受体为XH25和XH30时,转基因品系农艺性状与对照差异无显著水平;② 9个转基因品系间株高、始节高、始节数和铃数差异达到极显著水平(P<0.01),有效果枝数和有效铃数差异达到显著水平(P<0.05);③ 9个转基因品系的产量性状与对照差异均无显著性;④ 株高、始节高、始节数、果枝数、有效果枝数、铃数和有效铃数与籽棉产量呈显著相关;⑤ 两年(2014和2015)和两点(北疆和南疆)间农艺性状及产量性状差异达到显著水平。
转基因;Bt基因;海岛棉;农艺性状;产量性状;显著性
我们对转基因植株的研究在于获得有价值的中间材料和培育农作物新品种以及商业化生产[1-3],这些不仅与转基因技术有关,与转基因后代农艺性状特性也密切相关,农艺性状变异方向和幅度都是不确定的,基因的复杂性,环境的无常变化以及人类科学技术的不成熟对转基因技术的研究增加了难度,也为新品系的筛选和种质资源的创新带来了困难[4-5]。Bt基因是培育转基因抗虫棉过程中最早导入的外源基因,其优良的抗性使得转基因棉大规模种植,尤其在陆地棉中,已经得到推广和大面积种植,但在海岛棉中的研究甚少,也并没有开始推广种植。目前,新疆是我国唯一的海岛棉产区,随着棉花生产的不断发展,棉铃虫已成为严重影响海岛棉生产的重要因素之一,加强生物技术在海岛棉育种中的研究与应用,选育出适应性强的抗虫转基因品种,能较好地减轻棉铃虫的危害,对丰富海岛棉抗虫特异资源以及提高育种技术水平等方面均具有重要意义[6-12]。
转基因棉在抗虫的同时,其农艺性状、产量性状、纤维品质等性状与其受体品种存在显著差异[13-15]。本研究在分子检测的基础上,对转Bt基因海岛棉的农艺性状和产量性状进行研究分析,比较转基因棉与非转基因棉在不同性状上的差异,在遗传背景不同的情况下,比较同一基因对不同受体的影响,主要农艺性状对籽棉产量的影响,农艺性状及产量性状间相关分析以及在年份、地点、品系之间的差异,揭示外源基因Bt对海岛棉农艺性状和产量性状差异表现所产生的效应,为后人对转Bt基因海岛棉表型性状研究和利用提供参考。
1 材料和方法
1.1 试验材料
试验材料于2014年和2015年在北疆石河子和南疆阿克苏16团种植。试验采用随机区组设计,行长3 m,2行区种植,重复3次,留苗密度为11 800 株·667m-2,同时试验用地肥力比较一致,田间管理相同,材料编号及品系名称见表1,No.1-No.9为转基因品系,No.10-No.12为非转基因品系。
表1 材料名称
1.2 试验方法
1.2.1 转Bt基因海岛棉的PCR检测 以小区试验中采集的棉花叶片为材料,采用CTAB法提取棉花基因组DNA,PCR扩增体系为25 μl,扩增程序为94℃,5 min;94℃,30 s;57℃,30 s;72℃,1 min;72℃,10 min,共35个循环。扩增产物经1%琼脂糖凝胶电泳,再用溴化乙锭染色10 min,凝胶成像仪拍照。
1.2.2 转Bt基因对海岛棉农艺性状、产量的影响 于9月中旬棉花吐絮期,每个小区选取生长一致的棉花5株,测定株高、始节高、始节数、果枝数、有效果枝数、铃数和有效铃数。室内考察单铃重和衣分等,棉花收获期每行收棉株中部正常吐絮铃30个。
试验数据采用Excel和SPSS21.0统计软件进行统计分析。
2 结果与分析
2.1 PCR检测结果分析
如图1所示,对转基因品系No.1-No.9进行DNA提取,PCR扩增,以非转基因品系No.10-No.12为阴性对照,以含有目的片段的质粒为阳性对照,进行1%琼脂糖凝胶电泳检测和拍照分析。结果可以看出,样品均有特异性条带,且与阳性对照质粒条带大小一致,均在400 bp左右,而阴性对照没有扩增出目的条带。
2.2 转基因与非转基因之间农艺性状的比较
2.2.1 K222转Bt品系农艺性状与对照的比较 由表2得出No.5的株高与对照相比显著降低了11 cm,差异达显著水平;No.5、No.1、No.4、No.6和No.2的始节高与对照差异达到显著水平,始节高有伸长有缩短;No.7的始节数比对照高2.4 cm,差异达显著水平;No.1和No.3的果枝数比对照高4.2 cm和5 cm,差异达显著水平;No.5、No.1、No.4和No.3的有效果枝数与对照相比增加,差异达显著水平,No.1、No.4和No.3的铃数与对照相比增加,差异达显著水平;7个转基因品系的有效铃数与对照差异无显著性。由以上结果得出品系间除了有效铃数与对照没有差异外,其余各性状都发生变化,有的升高,有的降低;No.5、No.1、No.4和No.3这4个品系的3~4个性状与对照差异达显著水平,占总数的50%。
K222转Bt的7个品系的农艺性状之间差异显著,如图2所示,No.5的株高显著低于No.1和No.4,No.1的株高显著高于No.6、No.7、No.2和No.3,No.4的株高显著高于No.6和No.7。No.6和No.7的始节高显著高于No.2、No.3、No.4、No.1。No.6和No.7的始节数显著高于No.2、No.3、No.4、No.1。No.2、No.4和No.5的果枝数显著高于No.6,No.3和No.1的果枝数显著高于No.6、No.7。No.1、No.4和No.5的有效果枝数显著高于No.6和No.7,No.3的有效果枝数显著高于No.1、No.6和No.7。No.4和No.5的铃数显著高于No.6和No.7,No.3和No.1的铃数显著高于No.2、No.6和No.7。No.5的有效铃数显著高于No.6,No.1、No.4和No.3的有效果铃数显著高于No.6和No.7。

注:M:分子marker;1-9:转基因品系;10:阳性对照;11-13:阴性对照;
Note: M:DNA marker; 1-9:Transgenic plants; 10:Positive plasmid; 11-13:Non-transgenic plants
图1 转Bt基因品系PCR琼脂糖凝胶电泳检测
Fig.1 Agarose gel electrophoresis of transgenicBtPCR product
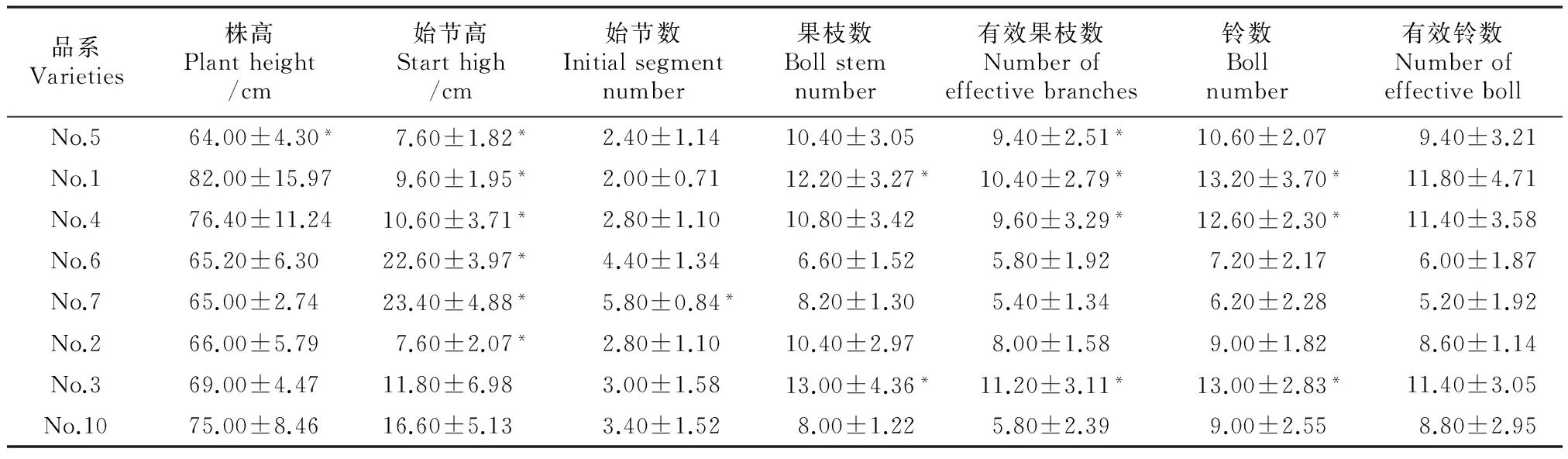
表2 转基因品系与对照农艺性状比较结果
注:*代表0.05显著水平;**代表0.01显著水平。
Note: * 0.05 represents significant level; **0.01 represents significant level; NG represents no significant level. The same below.
2.2.2 XH25-ZKC农艺性状与XH25,XH30-ZKC农艺性状与XH30的分析比较 经方差分析得出No.8、No.9的农艺性状结果与对照No.11、No.12差异无显著性;都保持着原品系的性状,可以说明Bt基因导入受体No.11和No.12,没有改变它们的农艺性状。
2.3 同一基因不同受体之间农艺性状的比较
由表3得出,各品系之间性状差异达到显著水

表3 同一基因不同受体农艺性状差异结果

图2 7个品系农艺性状差异
Fig.2 The difference of seven transgenic plants in agronomic characters
平所占比例为株高占66.67%、始节高占26.67%、始节数占20%、有效果枝数占6.67%、铃数占26.67%,有效铃数占6.67%,可以看出株高的差异幅度最大,始节高和铃数次之;No.7与No.8和No.9有差异的性状所占比例为66.67%,其次No.6为50%,其余各品系与No.8和No.9有差异的性状所占比例为8.33%和16.67%,说明No.7和No.6的农艺性状与No.8和No.9差异较大;No.8的农艺性状与No.9差异没有显著性。
由图3可以看出, No.5、No.6、No.7、No.2、No.3的株高与No.8相比极显著降低,No.5、No.6、No.7、No.2的株高与No.9相比极显著降低,No.3的株高与No.9相比显著降低,No.8的株高与No.9差异无显著水平,总体可以看出,外源基因Bt导入No.10、No.11和No.12不同受体中,得到No.11和No.12的株高普遍比No.10的高(71.43%)。No.8和No.9的始节高与No.6和No.7相比极显著降低,No.8的始节数、有效果枝数和有效铃数与No.7相比极显著降低,No.8的铃数与No.1和No.7相比极显著降低和显著升高。No.9始节数与No.6和No.7相比极显著降低,No.9的铃数与No.6和No.7相比显著升高。

图3 同一基因不同受体农艺性状差异
Fig.3 The difference of the same gene and different receptors in agronomic characters
2.4 产量性状的分析比较
2.4.1 转基因品系与非转基因品系之间产量性状比较 经方差分析和多重比较,得出No.10转Bt的7个品系的产量性状与对照相比差异无显著性。7个品系籽棉产量的范围在3 099.60~6 711.84 kg·hm-2之间,最高的为No.1,最低的为No.6,籽棉产量较高的品系为No.3、No.5和No.4,分别为6 279.12 kg、5 025.24 kg和5 827.68 kg。No.8的籽棉产量与No.11之间差异无显著水平,No.9籽棉产量与No.12之间差异无显著水平。
7个品系的单铃重的变化在2.84~3.49之间,整体变化较小,No.7的单铃重最大,达到了3.49,最小的是No.4,为2.84,42.86%的品系的单铃重在3.00以上,它们是No.1,No.7和No.3,它们的单铃重分别为3.16,3.49和3.06;7个品系的衣分含量在31%~42%之间,No.5和No.4的衣分最高,达到了42%,较高的品系还有No.6,衣分达到了37%,No.7的衣分最低;子指的范围在10.44~12.26之间,12.00以上的是No.2和No.3,分别为12.24和12.26,11.00以下的是No.6,为10.44。No.8的单铃重、衣分和子指与No.11之间差异无显著水平,No.9的单铃重、衣分和子指与No.12之间差异无显著水平。
2.4.2 同一基因不同受体之间产量性状差异 通过对同一基因不同受体的9个品系产量性状的方差分析和多重比较,得出仅有子指性状在品系间差异达到显著水平,其余性状差异均无显著性。由图4可以看出,受体为K222的转基因品系No.6的子指与受体为XH25的转基因品系No.8和受体为XH30的转基因品系No.9相比显著降低。
2.5 转基因品系农艺性状及产量相关分析
转基因品系各性状相关分析结果如表4,(1) 株高与果枝数、有效果枝数、铃数、有效铃数、单铃重、子指和籽棉产量呈正相关,与籽棉产量呈显著正相关,与始节高、始节数、衣分呈不显著负相关,可见增加株高会引起籽棉产量显著增加;(2) 始节高与果枝数、有效果枝数、铃数、有效铃数、衣分、子指、籽棉产量呈负相关,与果枝数、有效果枝数、有效铃数呈极显著负相关,与铃数、籽棉产量呈显著负相关,与始节数、单铃重呈正相关,与始节数呈极显著正相关,可见始节高的增加会引起始节数和单铃重增加,其余性状都会随始节高增加而降低;(3) 除了单铃重与始节数呈正相关以外,其余性状都与始节数呈负相关,其中果枝数、有效果枝数、铃数、有效铃数与始节数呈极显著负相关,与籽棉产量呈显著负相关;(4) 果枝数与有效果枝数、铃数、有效铃数、单铃重、子指、籽棉产量呈正相关,与有效果枝数、铃数、有效铃数、籽棉产量呈极显著正相关,与子指呈显著正相关,与衣分呈负相关,可见果枝数的增加会引起有效果枝数、铃数、有效铃数、子指和籽棉产量的增加;(5) 有效果枝数与铃数、有效铃数、衣分、子指、籽棉产量呈正相关,与铃数、有效铃数、籽棉产量呈极显著正相关,与单铃重呈负相关;(6) 铃数与有效铃数、衣分、子指、籽棉产量呈正相关,与有效铃数、籽棉产量呈极显著正相关,与单铃重呈负相关;(7) 有效铃数与衣分、子指、籽棉产量呈正相关,与籽棉产量呈极显著正相关,说明增加有效铃数会提高籽棉产量;(8) 单铃重与衣分、籽棉产量呈负相关,与子指呈正相关,衣分与子指呈负相关,与籽棉产量呈正相关,子指与籽棉产量呈正相关。
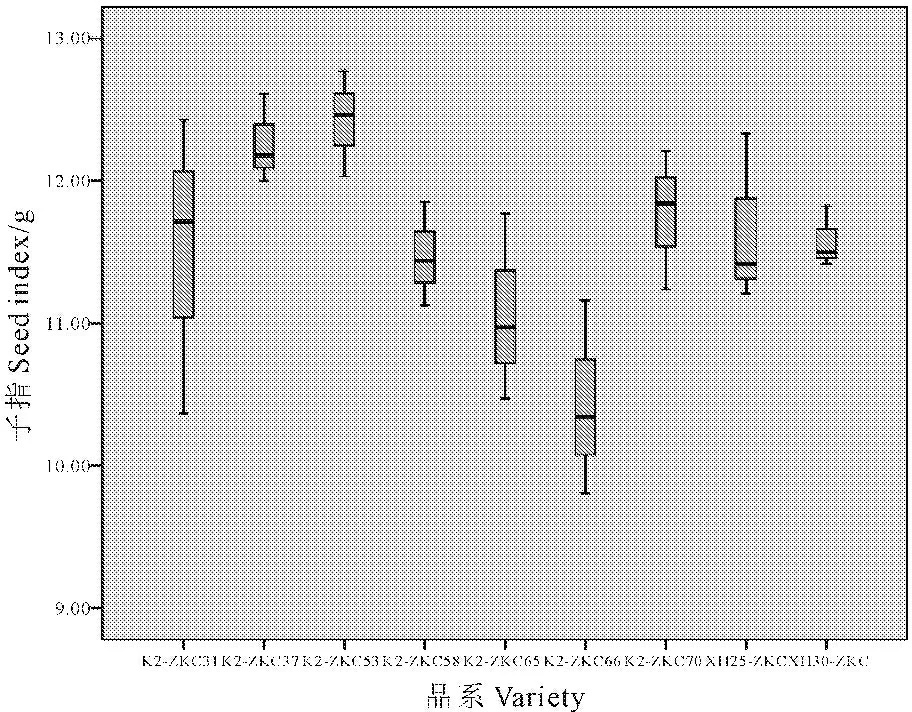
图4 同一基因不同受体间产量性状差异
Fig.4 The difference of the same gene and different receptors in yield characters
2.6 不同年份、地点和品系之间农艺性状及产量性状差异比较
通过对两年(2014年和2015年),两点(北疆和南疆)的农艺性状及产量性状差异比较得出(表5),转Bt基因海岛棉的农艺性状和产量性状在3个主效应(年份、地点和时间)以及3个两两互作效应(地点×品系、年份×品系和年份×地点)之间都有显著差异。(1) 年份间(2014年和2015年)除去株高、单铃重和子指没有显著差异,其他性状都有显著差异,地点间(北疆和南疆)有效果枝数、铃数、有效铃数、衣分和籽棉产量有显著差异,其它性状没有显著差异,品系间转基因与非转基因品系之间仅有单铃重和籽棉产量有显著差异,其它性状都无显著差异,农艺性状和产量性状在3个主效应之间的显著差异都是不同的,在3个主效应里,籽棉产量一直都是有显著差异的,说明年份、地点和品系能够影响籽棉产量的变化;(2) 地点×品系的互作效应只有株高和始节高这两个性状有显著差异,其它性状都无显著差异,年份×品系的互作效应在株高、始节高和始节数这三个性状上有显著差异,其它性状都无显著差异,年份×地点的互作效应在始节高、铃数、有效铃数和衣分这四个性状有显著差异,其它性状都无显著差异,在两两互作效应上,始节高一直都是有显著差异的;(3) 在年份×地点×品系的互作效应中各个性状差异都不显著。

表4 农艺性状及产量性状相关分析

表5 年份、地点和品系之间农艺性状和产量性状差异
续表5

源Origin因变量DependentvariableⅢ型平方和SquaressumofⅢDf均方MeansquareFSig.地点Areas品系Varieties地点×品系Area×Varieties年份×品系Year×Varieties株高Plantheight/cm144.4531144.4531.3730.242始节高Startheight/cm24.384124.3840.8290.364始节数Initialsegmentnumber0.58410.5840.5070.477果枝数Bollstemnumber23.835123.8352.6590.104有效果枝数Numberofeffectivebranches57.8157.88.2230.005**铃数Bollnumber154.0131154.01311.6580.001**有效铃数Numberofeffectiveboll316.6751316.67530.1520.000**单铃重Singlebollweight0.16810.1680.3970.530衣分Ginningoutturn5520.90915520.909838.3090.000**子指Seedindex/g0.08810.0880.1530.696籽棉产量Yield/(kg·hm-2)57604306.3815760430626.6980.000**株高Plantheight/cm93.528193.5280.8890.347始节高Startheight/cm13.475113.4750.4580.499始节数Initialsegmentnumber0.33410.3340.290.591果枝数Bollstemnumber14.168114.1681.580.210有效果枝数Numberofeffectivebranches15.606115.6062.220.138铃数Bollnumber26.068126.0681.9730.161有效铃数Numberofeffectiveboll5.08415.0840.4840.487单铃重Singlebollweight1.66911.6693.9530.049*衣分Ginningoutturn0.01610.0160.0020.960子指Seedindex/g1.30111.3012.2640.135籽棉产量Yield/(kg·hm-2)8426473.866184264743.9050.050*株高Plantheight/cm569.7781569.7785.4160.021*始节高Startheight/cm191.6841191.6846.5140.011*始节数Initialsegmentnumber0.110.10.0870.768果枝数Bollstemnumber13.068113.0681.4580.229有效果枝数Numberofeffectivebranches16.2116.22.3050.130铃数Bollnumber17.112117.1121.2950.256有效铃数Numberofeffectiveboll6.70916.7090.6390.425单铃重Singlebollweight1.63411.6343.8710.051衣分Ginningoutturn0.10910.1090.0170.898子指Seedindex/g0.26710.2670.4640.497籽棉产量Yield/(kg·hm-2)7978221.507179782223.6980.057株高Plantheight/cm2548.15312548.15324.220.000**始节高Startheight/cm211.7921211.7927.1980.008**始节数Initialsegmentnumber8.55918.5597.4320.007**果枝数Bollstemnumber9.56819.5681.0670.303有效果枝数Numberofeffectivebranches26.45126.453.7630.054铃数Bollnumber1.01311.0130.0770.782有效铃数Numberofeffectiveboll8.55918.5590.8150.368单铃重Singlebollweight0.34310.3430.8120.369衣分Ginningoutturn0.06910.0690.010.919子指Seedindex/g0.16710.1670.290.591籽棉产量Yield/(kg·hm-2)49744.804149744.80.0230.880
续表5

源Origin因变量DependentvariableⅢ型平方和SquaressumofⅢDf均方MeansquareFSig.年份×地点Year×Area年份×地点×品系Year×Area×Varieties误差Error总计Sum株高Plantheight/cm324.6841324.6843.0860.080始节高Startheight/cm192.7171192.7176.550.011*始节数Initialsegmentnumber0.25310.2530.220.640果枝数Bollstemnumber15.901115.9011.7740.184有效果枝数Numberofeffectivebranches15.022115.0222.1370.145铃数Bollnumber58.368158.3684.4180.037*有效铃数Numberofeffectiveboll45.25145.254.3090.039*单铃重Singlebollweight0.02110.0210.050.823衣分Ginningoutturn5545.86915545.869842.0990.000**子指Seedindex/g0.04810.0480.0830.773籽棉产量Yield/(kg·hm-2)7289957.712172899583.3790.068株高Plantheight/cm349.3091349.3093.320.070始节高Startheight/cm50.934150.9341.7310.190始节数Initialsegmentnumber0.00310.0030.0030.958果枝数Bollstemnumber0.00110.0010.0000.990有效果枝数Numberofeffectivebranches0.08910.0890.0130.911铃数Bollnumber11.001111.0010.8330.362有效铃数Numberofeffectiveboll1.7511.750.1670.683单铃重Singlebollweight0.41610.4160.9860.322衣分Ginningoutturn0.07610.0760.0120.915子指Seedindex/g0.02610.0260.0460.831籽棉产量Yield/(kg·hm-2)4543183.268145431832.1060.149株高Plantheight/cm24408.578232105.209始节高Startheight/cm6826.47823229.424始节数Initialsegmentnumber267.1672321.152果枝数Bollstemnumber2079.9442328.965有效果枝数Numberofeffectivebranches1630.7222327.029铃数Bollnumber3064.82223213.21有效铃数Numberofeffectiveboll2436.57823210.502单铃重Singlebollweight57.4021360.422衣分Ginningoutturn895.6641366.586子指Seedindex/g78.171360.575籽棉产量Yield/(kg·hm-2)2.934E81362157656株高Plantheight/cm1397069.25240始节高Startheight/cm46256.75240始节数Initialsegmentnumber2400.75240果枝数Bollstemnumber28772.5240有效果枝数Numberofeffectivebranches19222.5240铃数Bollnumber28080240有效铃数Numberofeffectiveboll22327.25240单铃重Singlebollweight1531.572144衣分Ginningoutturn31009.424144子指Seedindex/g19450.31144籽棉产量Yield/(kg·hm-2)3.757E9144
3 讨论与结论
3.1 转Bt基因海岛棉农艺性状和产量性状的变化
外源基因的导入对海岛棉农艺性状和产量性状有着不同程度的影响,本试验得出,14.29%转基因品系的株高、57.14%转基因品系的始节高与对照相比显著降低,28.57%转基因品系的始节高、14.29%转基因品系的始节数、28.57%转基因品系的果枝数、57.14%转基因品系的有效果枝数和42.86%转基因品系的铃数与对照相比显著增加,产量性状与对照差异无显著性。刘芳等[16]对转基因材料农艺性状分析得出,同一转基因品种(品系)的高代材料性状间存在广泛的分离和遗传变异,这与本试验结果相吻合。刘剑光等[17]研究Bt基因对棉花形态性状的影响,得出GK-12的株高明显低于泗棉3号,刘慧君等[18]研究GO基因导入对棉花农艺性状的影响,得出转基因棉株高极显著低于受体和显著低于对照,这与本试验得到的转基因品系株高显著降低一致,原因可能是外源基因在受体中表达消耗光合产物的积累和矿物质营养,使营养器官生长减慢,株型逐渐矮化。钱进等[19]对转MvNHX1基因棉花抗旱性研究和品系变异分析得出转基因品系在有效果枝数、单铃重和衣分上明显高于对照,这与本试验结果一致。在试验中也出现转基因品系农艺性状与对照相比无显著变化,比如No.8 和No.9,Bt基因的导入没有改变它们的农艺性状,与原品系保持一致,可能与连续种植几代后的基因沉默现象[20]、体细胞无系性变异有关[21-22],有些品系的农艺性状与对照相比有显著和极显著变化,这些都还可能与Bt基因插入的位点是随机的,插入的某些位点对某些性状基因表达产生了正面和负面的影响有关[23]。转基因农艺性状发生的变异越多,出现的有利性状几率就越大,可用于改良现有的品种和根据育种目标选择优良品种[24-25]。
3.2 Bt基因导入对不同受体农艺性状和产量性状的影响
同一基因不同受体间性状差异多数达到显著水平,当遗传背景不同时,相同基因的插入会对受体的表型产生不同的影响,比如Bt基因导入No.10、No.11和No.12受体中,在各品系之间株高变异的数目和范围是最多和最广泛的,始节高和铃数次之,有效果枝数和有效铃数最少;不同受体间转基因品系差异显著性有相同也有不同,比如No.5的株高与No.8和No.9差异都达到极显著水平,No.1的铃数与No.8差异都达到极显著水平,与No.9差异无显著性,No.8的性状与No.9差异没有显著性;不同受体间转基因植株的性状差异数目所占比例也不相同,比如No.7与No.8和No.9性状差异比例为66.67%,No.6与No.8和No.9性状差异比例为为50%,其余品系与No.8和No.9性状差异比例仅为8.33%和16.67%,说明No.7和No.6的农艺性状与No.8和No.9差异较大。对于这些性状差异数目出现多的,性状变异广泛的材料,我们难以直接利用,但可以加以研究和通过常规育种手段改良[26-29],筛选出性状优良的品系,为育种提供基础。
3.3 转基因品系农艺性状对产量的影响
棉花作为一种重要的经济作物,其使用价值及商品价值都是其他作物不可替代,无论导入何种基因,都是为其高产、稳产、优质、高效等服务,都是改良其不足,发挥更大价值。没有高产的作物,即使品质再好都是不可选育的,所以研究农艺性状对产量的相关性对新品种选育有重要的指导意义。本研究中株高、果枝数、有效果枝数、铃数、有效铃数与籽棉产量呈显著正相关,与刘昌文得到的株高、铃数与产量呈正相关一致[30];始节高与始节数与籽棉产量呈显著负相关,根据农艺性状与产量的相关性,可人为调控棉花的生长,减少田间管理的盲目性。
3.4 两年和两点间农艺性状和产量性状的变化
年份、地点、品系、地点×品系的互作效应、年份×品系的互作效应和年份×地点的互作效应之间农艺性状及产量性状有不同程度的差异,不同年份和不同地点之间气候差异较大,比如温度,温度变化会影响棉花的生理活性,一些性状表达所需能量相对不足,生理代谢受到影响,表型发生变化[31-32]。本研究可以为今后海岛棉田间表型性状研究提供参考依据。
[1] 何龙飞,韦善清,卢升安,等.转SCK基因水稻后代农艺性状变异的研究[J].广西农业生物科学,2002,21(2):73-77.
[2] Tabashnik B E, Van Rensburg. Field-evolved insect resistance to bt crops: definition, theory, and data[J]. Journal of Economic Entomology, 2009,102(6):2011-2025.
[3] Lu YH, Wu KM, Jiang YY, et al. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in chin[J]. Science, 2010,328(5982):1151-1154.
[4] 张 欣,付亚萍,周君莉,等.水稻规模化转基因技术体系构建与应用[J].中国农业科学,2014,47(21):4141-4154.
[5] Flores S, Saxena D, Stotzky G. Transgenic Bt plants decompose less in soil than non-Bt plants[J]. Soil Biology & Biochemistry,2005,37(6):1073-1082.
[6] 喻树迅.我国棉花生产现状与发展趋势[J].中国工程科学,2013,15(4):9-13.
[7] 张 锐,王 远,孟志刚,等.国产转基因抗虫棉研究回顾与展望[J].中国农业科技导报,2007,9(4):32-42.
[8] 秦永华,乔志新,刘进元.转基因技术在棉花育种上的应用[J].棉花学报,2007,19(6):482-488.
[9] 郑巨云,李雪源,王俊铎,等.新疆转基因棉花育种展望[J].中国棉花,2010,37(11):2-5.
[10] 王孝纲,别 墅,张教海,等.我国转基因棉花育种概况与展望[J].湖北农业科学,2003,(3):32-35.
[11] Tabashnik BE, Carriere Y, Dennehy TJ, et al. Insect resistance to transgenic Bt crops: Lessons from the laboratory and field[J]. Journal of Economic Entomology, 2003,96(4):1031-1038.
[12] Romeis J, Meissle M, Bigler F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control[J]. Nature Biotechnology, 2006,24(1):63-71.
[13] 丰 嵘,张宝红,郭香墨.外源Bt基因对棉花产量性状及抗虫性的影响[J].棉花学报,1996,8(1):10-13.
[14] 张香桂,周宝良,陈 松,等.抗虫棉与常规棉主要农艺及经济性状的比较研究[J].中国棉花,2004,31(5):14-16.
[15] 刘冬梅,武芝霞,刘传亮,等.花粉管通道法获得棉花转基因株系主要农艺性状变异分析[J].棉花学报,2007,19(6):450-454.
[16] 刘 方,王坤波,宋国立,等.棉花转基因材料的获得及主要农艺性状变异分析[J].棉花学报,2009,21(1):23-27.
[17] 刘剑光,肖松华,狄佳春,等.Bt基因导入对棉花农艺性状的影响[J].中国棉花,2003,30(3):15-17.
[18] 刘慧君,简桂良,邹亚飞.GO基因导入对棉花农艺性状及抗病性的影响[J].分子植物育种,2003,1(5/6);669-672.
[19] 钱 进,杨云尧,任燕萍,等.转MvNHX1基因棉花抗旱性研究和株系变异分析[J].分子植物育种,2014,12(1):80-86.
[20] 连丽君,吕素莲,李汝忠,等.转BetA/als基因棉花材料的农艺性状考察[J].棉花学报,2008,20(6):447-451.
[21] 崔海瑞,王忠华,舒庆尧,等.转Bt基因水稻克螟稻杂交转育后代农艺性状的研究[J].中国水稻科学,2001,15(2):22-27.
[22] 唐祚舜,李良材,田文忠.基因枪法转基因水稻后代农艺性状的表现[J].中国农业科学,2001,34(6):581-586.
[23] 莫春红.转Bt基因水稻Bt蛋白表达量检测及螟虫抗性和农艺性状评价[D].武汉:华中农业大学,2014.
[24] 徐泽俊,聂以春,张献龙,等.转双价抗虫基因棉花的主要农艺性状的遗传变异[J].植物遗传资源学报,2011,12(1):125-130.
[25] 吕淑平,郭小平,赵元明.转基因抗虫棉Bt基因导入对受体材料农艺性状的影响[J].中国农学通报,2004,20(3):36-37.
[26] 王忠华,张立成.Bt抗虫转基因水稻的育种应用[J]. 核农学报,2007,21(2):148-151,200.
[27] 陈文岳.转Bt基因水稻“克螟稻”的遗传改良研究[D].杭州:浙江大学,2005.
[28] 王彩霞,吴殿星,沈圣泉,等.转基因及常规水稻幼苗对潮霉素敏感性的研究[J]. 核农学报,2005,19(3):168-171.
[29] 沈圣泉,舒庆尧,包劲松,等.应用近等基因系研究Bt基因对水稻性状表现的影响[J].浙江大学学报(农业与生命科学版),2005,31(3):283-287.
[30] 刘昌文,张 燕,宋义前.新疆早熟棉花主要农艺性状相关性及多项式趋势分析[J].天津农业科学,2008,14(3):11-16.
[31] 周桂生,周福才,谢义明,等.温度胁迫对转Bt基因抗虫棉毒蛋白的表达和棉铃虫死亡率的影响[J].棉花学报,2009,21(4):302-306.
[32] 沈 平,林克剑,张永军,等.转Bt基因棉不同品种杀虫蛋白季节性表达及其对棉铃虫的控制作用[J].棉花学报,2010,22(5):393-397.
Effects of introducingBtgene on agronomic characters and yield characters ofGossypiumbarbadenseL.
JIA Li-li, WU Peng-hao, LI Juan, WANG Zhi-xing, CHEN Quan-jia,WANG Xi-dong, QU Yan-ying
(XinjiangAgriculturalUniversity,Urumqi,Xingjiang830052,China)
The agronomic and yield characters of nine strains ofGossypiumbarbadenseL., in whichBtgene was detected by PCR, were studied. The results showed that agronomic characters and yield characters were impacted to different extent.(1) When K222 was the recipient, the difference in plant height, start high, initial segment number, number of effective branches, boll number and number of effective bolls got to excellent significant level (P<0.01). When XH25 and XH30 were recipients, there was no difference between transgenic and non-transgenic; (2) The difference in plant height, start high, initial segment number and boll number between the nine strains got to excellent significant level (P<0.01). Difference in number of effective branches and number of effective boll arrived at significant level (P<0.05); (3) There was no significant difference between transgenic and non-transgenic in yield characters; (4) Plant height, start high, initial segment number, boll stem number, number of effective branches, boll number and number of effective boll were significantly related to yield; (5) The difference between two years (2014 and 2015) and two areas(the northern of Xinjiang and the southern of Xinjiang) got to significant level in agronomic characters and yield characters.
transgenic;Btgene;GossypiumbarbadenseL.; agronomic characters; yield characters; significant level
1000-7601(2017)02-0103-11
10.7606/j.issn.1000-7601.2017.02.17
2016-01-01基金项目:转基因长绒棉新品种培育(2014ZX08005-005-009)
贾莉莉(1991—),新疆人,研究生,研究方向棉花分子育种。 E-mail:595851794@qq.com。
王希东,硕士生导师,E-mail:372257510@qq.com。 曲延英,博士生导师,主要从事作物遗传育种。 E-mail:xjyyq5322@126.com。
S331
A
猜你喜欢
棉纺织技术(2021年10期)2021-10-15
今日农业(2020年14期)2020-12-14
现代农村科技(2020年9期)2020-09-08
中国棉花加工(2020年1期)2020-07-01
棉花科学(2020年2期)2020-06-01
中国棉花加工(2020年5期)2020-01-15
中国纤检(2019年5期)2019-12-09
云南农业科技(2019年2期)2019-04-04
中国棉花(2018年7期)2018-08-29
安徽农学通报(2016年7期)2016-05-03
