
海拔高度对马铃薯地方品种形态结构及解剖结构的影响
2017-04-20 07:24:15王文祥达布希拉图周平沙本才耿世英刘佳海梅荣
中国马铃薯 2017年2期
王文祥,达布希拉图,周平,沙本才,耿世英,刘佳,海梅荣*
(1.云南农业大学农学与生物技术学院,云南昆明650201;2.云南农业大学资环学院,云南昆明650201)
栽培生理
海拔高度对马铃薯地方品种形态结构及解剖结构的影响
王文祥1,达布希拉图2,周平1,沙本才1,耿世英1,刘佳1,海梅荣1*
(1.云南农业大学农学与生物技术学院,云南昆明650201;2.云南农业大学资环学院,云南昆明650201)
为了研究海拔梯度对马铃薯地方品种形态结构和解剖结构的影响,测定和分析了在不同海拔种植的3个马铃薯地方品种的形态和解剖结构特点。结果表明,3个马铃薯地方品种的自身组织结构随着海拔的升高而存在着一定的规律性却又向着有利于生长方向发展而存在着特异性。3个品种的株高、分枝长和分枝数都是在海拔2 000 m处的数值最大,茎粗都是在海拔1 000 m处最大;3个品种的上下表皮厚度都是随着海拔的升高而增加(除‘盈江小洋芋’的下表皮厚度、‘老家洋芋’的上表皮厚度);‘盈江小洋芋’和‘老家洋芋’的叶片栅栏组织厚度均是在海拔3 000 m处最大,‘剑川红’在3个海拔的叶片栅栏组织厚度差异不大。3个品种在海拔3 000 m处的海绵组织厚度最大;3个品种的气孔开度都是在海拔1 000 m处最大,海拔2 000 m处最小;除‘老家洋芋’外,其他2个品种的气孔数都是随着海拔的升高而增加,而‘老家洋芋’在海拔2 000 m处的气孔数最多,在海拔3 000 m处的气孔数最少。
马铃薯地方品种;海拔梯度;形态结构;解剖结构
马铃薯(Solanum tuberosum L.),茄科茄属,又名洋芋、土豆、山药蛋、荷兰薯等,是1种块茎植物,也是全球第三大重要的粮食作物,仅次于小麦和玉米。马铃薯逐步成为人类重要的粮食、蔬菜、饲料和工业原料。
云南省位于E 97°31'~106°11',N 21°8'~29°15',地域辽阔,是高原山区省份[1],其生态环境、气候条件适宜马铃薯的生长,是中国西南地区的马铃薯主产区之一。马铃薯是云南省4大粮食作物之一,常年种植面积在22万hm2左右,单产在13 000 kg/hm2。由于马铃薯对农业经济促进和农民增收作用明显,被云南省列为重点发展的优势产业[1,2]。
云南省是中国的生物资源宝库,也是品种资源中心,在云南少数民族居住地区,还保留着丰富的马铃薯种质资源,现在内地品种正趋于单一化时,在云南省高寒山区和热带河谷有着丰富的马铃薯地方品种[3]。研究开发云南省地方品种对马铃薯产业的发展有着深厚的意义。随着人民生活水平的提高,对马铃薯的产量和品质提出了更高的要求。云南省有着高原特殊的立体气候,生态环境的多样性和气候的复杂性,造成不同区域品种的多样性[4]。
云南省地势复杂多变,高山河谷纵横交错,不同海拔地区马铃薯生长表现出的形态结构不同,不同海拔地区为马铃薯提供了不同的生长环境,把马铃薯放到不同的海拔地区,能表现出不同的形态结构[5]。海拔是不同环境因素的综合表现,各种环境因素在不同海拔梯度上表现出相应的变化,这为研究植物生理形态对环境的响应提供了良好的条件。在不同海拔梯度上,各种环境因素如温度、空气湿度、水分状况和光照强度等均发生了不同程度的变化。这些环境因素的改变会对植物叶片结构产生一定的影响[6]。植物在发育过程中,根、茎、叶在植物体内具有一定的形态结构,蕴含着一定的生理功能,在生理活动中影响着植物体的生长和发育。叶片作为植物进行光合作用和呼吸作用的主要器官,与周围环境联系紧密,对环境因子如水分、温度、光照等的变化敏感,随着环境变化,叶片在外部形态、厚度及内部解剖结构等方面表现出一定差异[7]。
不同海拔地区有着多样的气候条件和环境资源,如光照辐射、气温、热量、降雨量和土壤条件。马铃薯有着不同的形态结构和生长习性,把不同地方马铃薯品种种植在不同的海拔表现出不同的差异,前人在此基础上研究较少[8]。以‘盈江小洋芋’、‘剑川红’和‘老家洋芋’3个地方品种为研究材料,在不同海拔进行试验,通过形态指标、解剖结构的分析和比较,为今后对不同海拔地区马铃薯种植情况及产量的提高提供理论依据[9]。
1 材料与方法
1.1 试验材料与设计
试验地一位于昆明市禄劝县皎西乡皎平渡,海拔1 000 m左右,当地年均温10.85℃、年降雨量972.2 mm;试验地二位于云南省禄劝县转龙镇普卡里大法期,海拔1 998 m,当地年均温14.2℃,年降雨量1 000 mm,光照强度大、日照时间长;试验地三位于昆明市禄劝县转龙镇轿子雪山炭山村,海拔2 988 m,当地阴雨绵绵,低压、高湿紫外线强,年均温10.0℃,年降雨量920 mm。3个试验地年干湿季节明显,5~10月为雨季,11月至次年的4月为旱季,属亚热带季风气候。供试马铃薯品种为:‘盈江小洋芋’(出自滇西南,海拔1 000 m左右),‘剑川红’(出自滇西北,海拔2 000 m左右),‘老家洋芋’(出自大理,海拔2 800 m左右),构成了来自低、中、高3个海拔高度的品种。
2013年12月22日,在3个海拔分别选取667 m2地块,再分别划分9个小区,每个品种种植3个小区,作为3次重复。所有小区统一正常施肥种植。
1.2 植株形态测定
在各个海拔的马铃薯生长的开花期,用卷尺在每个小区随机选取3株测定马铃薯株高、分枝长,电子数显游标卡尺测定茎直径,并统计分枝数。
1.3 解剖结构制作与测定
在开花期(2014年3月)从不同海拔将马铃薯取回,在各个海拔中,每个品种的3次重复小区中,随机移栽3盆已标记马铃薯,带回云南农业大学后山试验地进行管理。选取每株的中部叶片用冰盒保存,并带回实验室作切片材料。
1.3.1 马铃薯叶片横切片制作方案
取材:将马铃薯叶片(取无叶脉部分)切成3~5mm的小方块。
固定:将材料放入事先准备好的FAA固定液中保存12 h以上。
脱水:取出固定的材料放入酒精中脱水,酒精的浓度从50%升到95%,每个浓度中浸泡30 min再取出,放入另1个浓度中,最后放入纯酒精中浸泡2 h,取出放入新的纯酒精中,过夜后取出,放入纯酒精中浸泡2 h。
透明:取出酒精中的材料放入二甲苯与酒精的混合溶液中浸泡,二甲苯与酒精的浓度配比为1∶2,1∶1,2∶1,分别浸泡3 h,最后放到纯二甲苯中3次,分别浸泡6 h。
浸蜡:分别从材料中倒出1/3,1/2,2/3,全部的二甲苯,加入融化的石蜡,放到烘箱中,烘箱温度分别为35,45和60℃,每次放入烘箱中6 h。
包埋:将浸蜡材料倒入纸盒中,冷却放置。
切片与粘片:将包埋好的石蜡修块,粘片,用专业的切片机(HK-168)切成厚度为10 μm的连续切片,用毛笔将蜡片展平放置在载玻片上,放到40℃烘箱中3 d。
脱蜡与染色:将有材料的玻璃片放入纯二甲苯中浸泡30 min,取出放入二甲苯与酒精混合溶液(1∶1)30 min。取出后分别放入酒精浓度为100%,95%,90%,80%,70%,60%,50%,30%,15%,蒸馏水中各15 min。再放入铁矾(2%)2 h,苏木精染色(过夜),取出后分别放入蒸馏水,酒精浓度为15%,30%,50%,60%,70%,80%,90%,95%,100%,二甲苯∶酒精为1∶1,100%的二甲苯中各15 min。
粘片:从二甲苯中取出玻片,立即用已融化的中性树胶封固,放入40℃烘箱中烘2~3 d。
待切片制作完成,在10 X和40 X显微镜下观察拍照,对上表皮、下表皮、栅栏组织、海绵组织厚度进行测定记录。
1.3.2 马铃薯叶片气孔数、气孔开度切片制作方案
将马铃薯叶片从冰盒中取出,用镊子撕取(长:3 mm;宽:2 mm)下表皮,放在准备好的载玻片上,盖上盖玻片,然后放到Motic Images Advanced 3.2数码成像显微镜(目镜10×10,物镜10×40)下观察并拍照,目镜(10×20)下观察气孔数并拍照记录,再调整目镜在(10×40)下观察气孔并拍照记录。将拍好的照片用Photoshop 7.0软件打开,用测量尺测量,取1 mm2的气孔数进行比较。
1.4 数据分析
运用Excel 2007和SPSS17.0(SPSS Inc.,USA)软件统计分析试验数据,利用Duncan's法进行平均数的多重比较[10]。
2 结果与分析
2.1 马铃薯各形态指标结果比较
2.1.1 马铃薯株高比较
由表1可知,‘盈江小洋芋’在海拔2 000 m处的株高极显著高于海拔1 000和3 000 m,其余不显著;‘剑川红’在海拔2 000 m处的株高极显著高于海拔1 000和3 000 m,‘剑川红’在海拔3 000 m处的株高显著高于海拔1 000 m,其余不显著;‘老家洋芋’在海拔2 000 m处的株高极显著高于海拔1 000和3 000 m,其余不显著。在海拔1 000 m处,‘剑川红’与‘老家洋芋’的株高均极显著高于‘盈江小洋芋’,其余不显著;在海拔2 000 m处,‘剑川红’与‘老家洋芋’的株高均极显著高于‘盈江小洋芋’,其余不显著;在海拔3 000 m处,‘剑川红’的株高显著高于‘老家洋芋’,‘剑川红’和‘老家洋芋’的株高均极显著高于‘盈江小洋芋’。
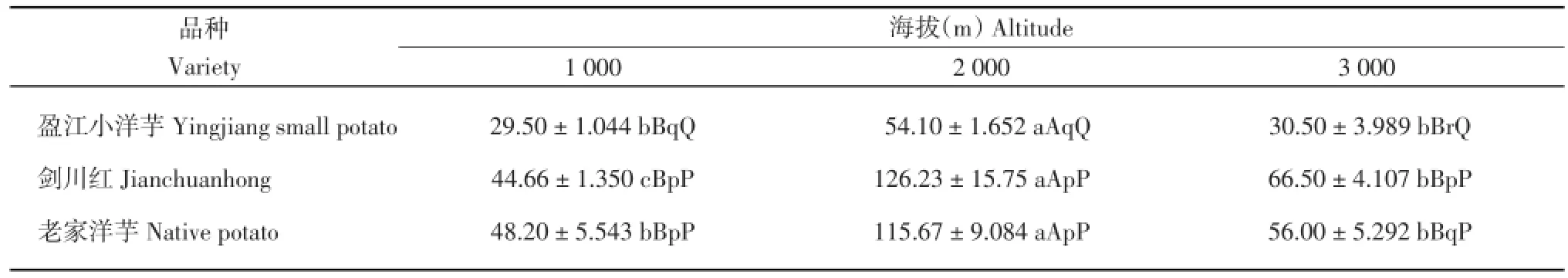
表1 3个海拔地区的马铃薯株高(cm)Table 1 Plant height of potatoes at three altitudes
2.1.2 马铃薯分枝数比较
由表2可知,‘盈江小洋芋’在海拔2 000 m处的分枝数显著高于海拔1 000 m,其余不显著;‘剑川红’在海拔2 000 m处的分枝数显著高于海拔3 000 m,‘剑川红’在海拔2000m处的分枝数极显著高于海拔1000m,‘剑川红’在海拔3000m处分枝数显著高于海拔1000m;‘老家洋芋’在海拔2000m处的分枝数极显著高于海拔1 000 m,‘老家洋芋’在海拔2 000 m处的分枝数显著高于海拔3 000 m,其余不显著。在海拔1 000 m处,3个品种的分枝数差异不显著;在海拔2 000 m处,‘剑川红’的分枝数显著高于‘盈江小洋芋’,其余不显著;在海拔3 000 m处,‘剑川红’的分枝数显著高于‘盈江小洋芋’和‘老家洋芋’,其余不显著。
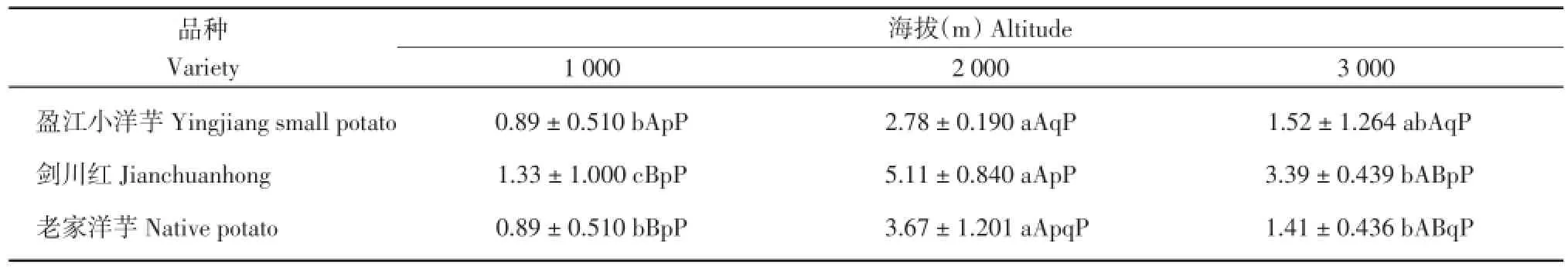
表2 3个海拔地区的马铃薯分枝数(No.)Table 2 Branches of potatoes at three altitudes
2.1.3 马铃薯分枝长比较
由表3可知,‘盈江小洋芋’在海拔2 000和3 000 m处的分枝长均极显著高于海拔1 000 m,‘盈江小洋芋’在海拔2 000 m处的分枝长显著高于海拔3 000 m;‘剑川红’在海拔2 000 m处的分枝长极显著高于海拔1 000 m,‘剑川红’在海拔2 000 m处的分枝长显著高于海拔3 000 m,‘剑川红’在海拔3 000 m处的分枝长极显著高于在海拔1 000 m;‘老家洋芋’在海拔2 000和3 000 m处的分枝长均极显著高于海拔1000m,其余不显著。在海拔1000m处,3个品种的分枝长差异不显著;在海拔2 000 m处,‘剑川红’的分枝长显著高于‘老家洋芋’,‘剑川红’和‘老家洋芋’的分枝长均极显著高于‘盈江小洋芋’;在海拔3 000 m处,‘剑川红’和‘老家洋芋’的分枝长极显著高于‘盈江小洋芋’,其余不显著。
2.1.4 马铃薯茎粗比较
由表4可知,‘盈江小洋芋’在海拔1 000 m处的茎粗极显著高于海拔2 000和3 000 m,其余不显著;‘剑川红’在海拔1 000 m处的茎粗极显著高于海拔2 000和3 000 m,其余不显著;‘老家洋芋’在海拔1 000 m处的茎粗极显著高于海拔2 000和3 000 m,其余不显著。在海拔1 000 m处,‘剑川红’的茎粗显著高于‘老家洋芋’,‘剑川红’和‘老家洋芋’的茎粗极显著高于‘盈江小洋芋’;在海拔2 000 m处,‘剑川红’的茎粗显著高于‘老家洋芋’,‘剑川红’和‘老家洋芋’的茎粗均极显著高于‘盈江小洋芋’;在海拔3 000 m处,‘老家洋芋’的茎粗显著高于‘盈江小洋芋’,其余不显著。
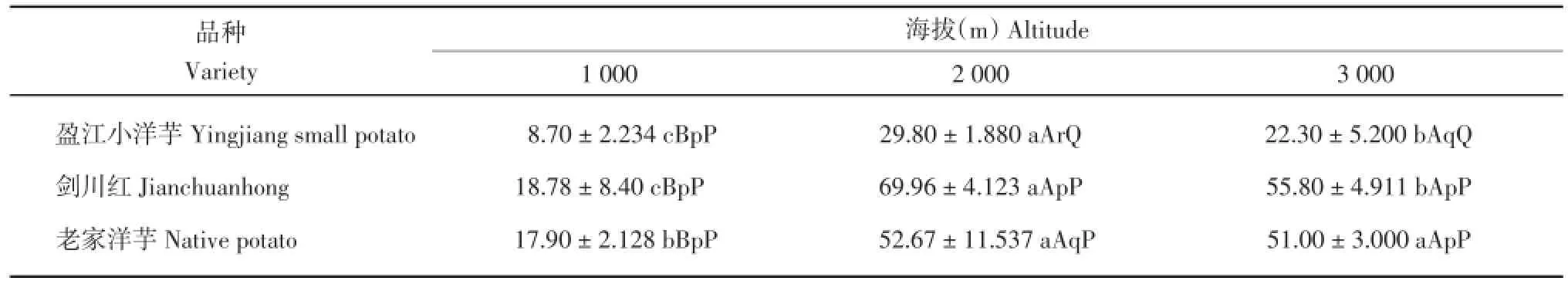
表3 3个海拔地区的马铃薯分枝长(cm)Table 3 Branch length of potatoes at three altitudes
2.2 马铃薯各叶片解剖结构指标结果比较
2.2.1 马铃薯叶片上表皮厚度比较
由表5可知,‘盈江小洋芋’在海拔3 000 m处的叶片上表皮厚度显著高于海拔1 000 m,其余不显著;‘剑川红’在海拔3 000 m处的叶片上表皮厚度极显著高于海拔1 000 m,其余不显著;‘老家洋芋’在海拔2 000 m的叶片上表皮厚度显著高于海拔1 000和3 000 m,其余不显著。在海拔1 000 m处,3个马铃薯品种的叶片上表皮厚度差异不显著;在海拔2 000 m处,‘老家洋芋’的叶片上表皮厚度极显著高于‘剑川红’,其余不显著;在海拔3 000 m处,3个马铃薯品种的叶片上表皮厚度差异不显著。
2.2.2 马铃薯叶片栅栏组织厚度比较
由表6可知,‘盈江小洋芋’在海拔3 000 m处的叶片栅栏组织厚度极显著高于海拔1 000和2 000 m,其余不显著;‘剑川红’在3个海拔处的差异不显著;‘老家洋芋’在海拔3 000 m处的栅栏组织厚度显著高于海拔2 000 m,其余不显著。在海拔1 000 m处,‘老家洋芋’的叶片栅栏组织厚度极显著高于‘盈江小洋芋’和‘剑川红’,其余不显著;在海拔2 000 m处,‘剑川红’叶片栅栏组织厚度显著高于‘盈江小洋芋’,其余不显著;在海拔3 000 m处,‘老家洋芋’叶片栅栏组织厚度极显著高于‘剑川红’,其余不显著。2.2.3马铃薯叶片海绵组织厚度比较
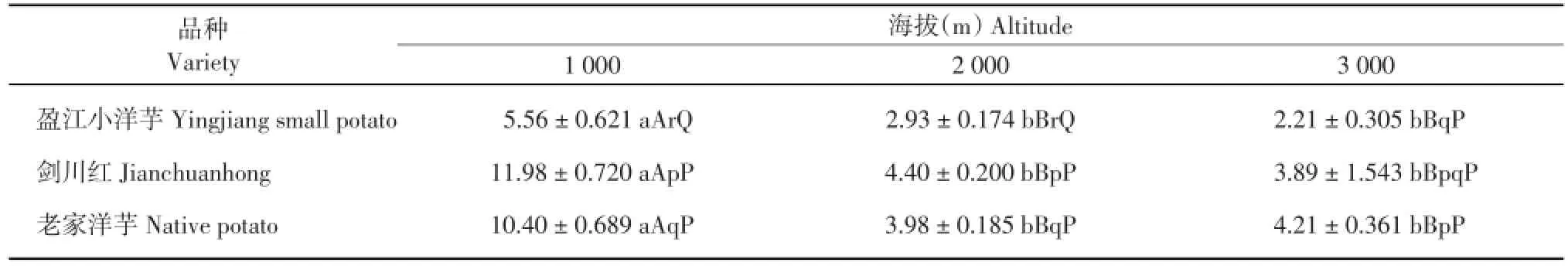
表4 3个海拔地区的马铃薯茎粗(mm)Table 4 Stem diameter of potatoes at three altitudes
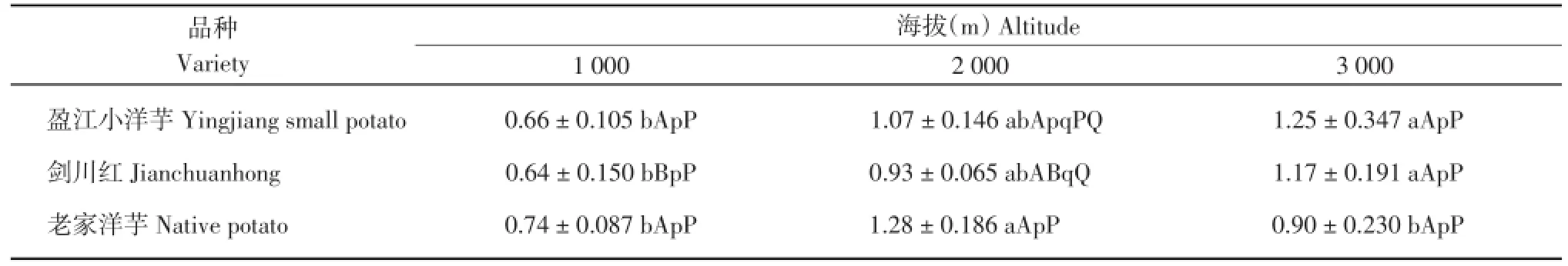
表5 3个海拔地区的马铃薯上表皮厚度(μm)Table 5 Upper epidermis thickness of potatoes at three altitudes
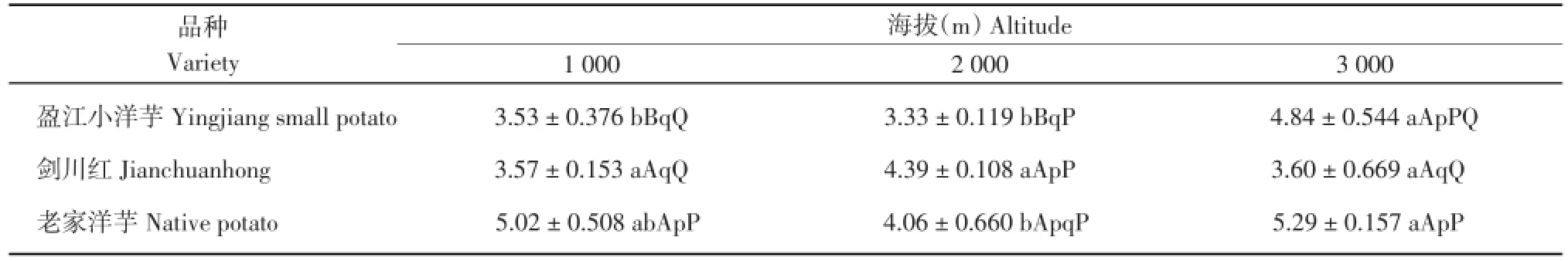
表6 3个海拔地区的马铃薯栅栏组织厚度(μm)Table 6 Palisade tissue thickness of potatoes at three altitudes
由表7可知,‘盈江小洋芋’在海拔3 000 m处的叶片海绵组织厚度极显著高于海拔1 000和2000m,其余不显著;‘剑川红’在海拔3000m处的叶片海绵组织厚度显著高于海拔1 000和2 000 m,其余不显著;‘老家洋芋’在海拔3 000 m处的叶片海绵组织厚度极显著高于海拔1 000和2 000 m,‘老家洋芋’在海拔1 000 m处的叶片海绵组织厚度极显著高于海拔2 000 m。在海拔1 000 m处,‘老家洋芋’的叶片海绵组织厚度极显著高于‘盈江小洋芋’和‘剑川红’,其余不显著;在海拔2 000 m处,‘盈江小洋芋’和‘老家洋芋’的叶片海绵组织厚度均极显著高于剑川红’,其余不显著;在海拔3 000 m处,‘盈江小洋芋’和‘老家洋芋’的叶片海绵组织厚度均极显著高于‘剑川红’,其余不显著。
2.2.4 马铃薯叶片下表皮厚度比较
由表8可知,‘盈江小洋芋’在海拔2 000 m处的叶片下表皮厚度显著高于海拔1 000 m,其余不显著;‘剑川红’在3个海拔处的叶片下表皮厚度差异不显著;‘老家洋芋’在海拔3 000 m处的叶片下表皮厚度极显著高于海拔1 000 m,其余不显著。在海拔1 000 m处,3个品种的叶片下表皮厚度差异不显著;在海拔2 000 m处,3个马铃薯品种的叶片下表皮厚度差异不显著;在海拔3 000 m处,3个马铃薯品种的叶片下表皮厚度差异不显著。
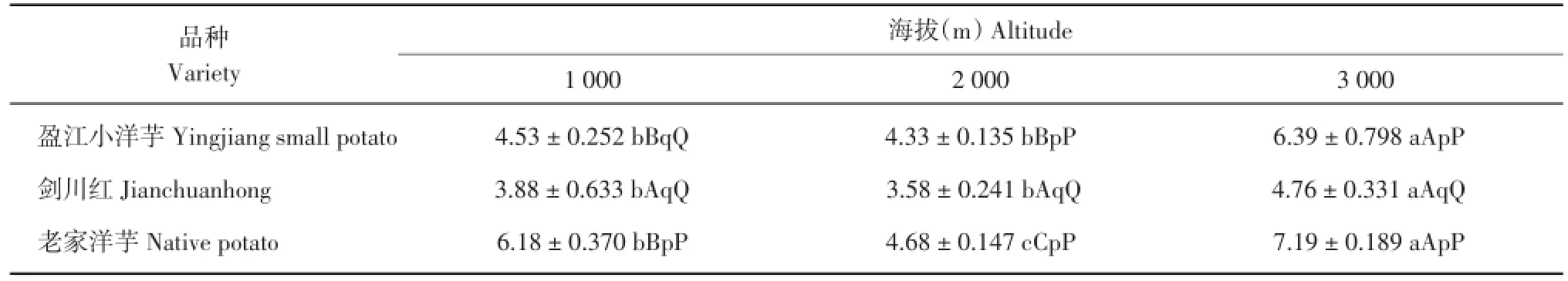
表7 3个海拔地区的马铃薯海绵组织厚度(μm)Table 7 Sponge tissue thickness of potatoes at three altitudes
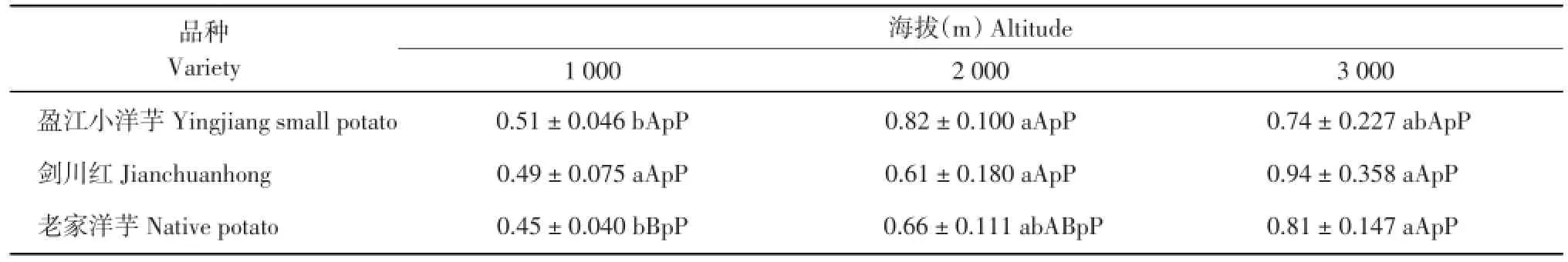
表8 3个海拔地区的马铃薯下表皮厚度(μm)Table 8 Lower epidermis thickness of potatoes at three altitudes
2.2.5 马铃薯叶片气孔开度比较
由表9可知,‘盈江小洋芋’在海拔1 000 m处的叶片气孔开度显著高于海拔2 000和3 000 m,其余不显著;‘剑川红’在海拔1 000 m处的叶片气孔开度显著高于海拔2 000和3 000 m,其余不显著;‘老家洋芋’在海拔1 000 m处的叶片气孔开度极显著高于海拔2 000和3 000 m,‘老家洋芋’在海拔3 000 m处的叶片气孔开度显著高于海拔2 000 m。在海拔1 000 m处,3个马铃薯品种的叶片气孔开度差异不显著;在海拔2 000 m处,‘剑川红’的叶片气孔开度显著高于‘老家洋芋’和‘盈江小洋芋’,其余不显著;在海拔3 000 m处,3个马铃薯品种的叶片气孔开度差异不显著。
2.2.6 马铃薯叶片气孔数比较
由表10可知,‘盈江小洋芋’在海拔3000m处的叶片气孔数极显著高于海拔1 000和2 000 m,其余不显著;‘剑川红’在3个海拔的叶片气孔数差异不显著;‘老家洋芋’在海拔2000m处的叶片气孔数极显著高于海拔3 000 m,其余不显著。在海拔1 000 m处,‘老家洋芋’的叶片气孔数极显著高于‘盈江小洋芋’和‘剑川红’,其余不显著。在海拔2 000 m处,‘老家洋芋’的叶片气孔数极显著高于‘盈江小洋芋’,其余不显著;在海拔3 000 m处,3个品种的叶片气孔数差异不显著。
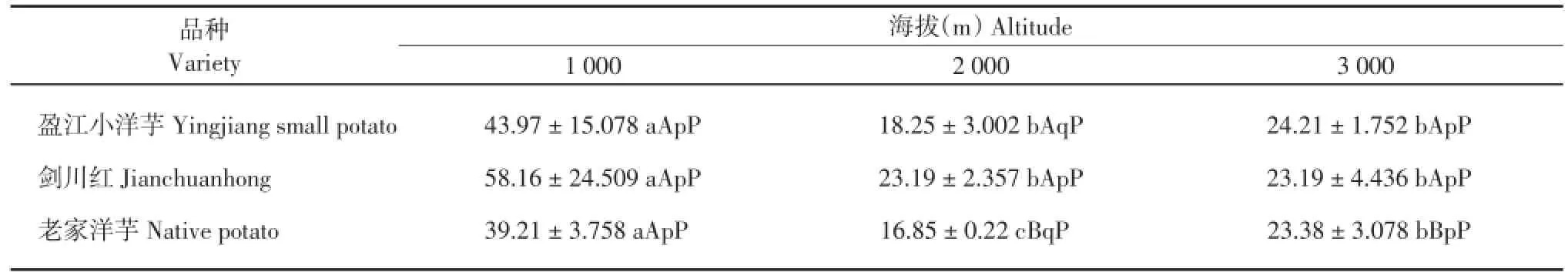
表9 3个海拔地区的马铃薯气孔开度(μm2)Table 9 Stomatal aperture of potatoes at three altitudes

表10 3个海拔地区的马铃薯气孔数(No./mm2)Table 10 Stomata number of potatoes at three altitudes
3 讨论
不同海拔,带来多样的环境条件,光照、温度不同对马铃薯的生长环境造成一定的影响。试验通过3个地方品种马铃薯的形态结构、解剖结构的测定分析研究海拔梯度对马铃薯形态结构、解剖结构的影响[11]。
试验的马铃薯品种分别来自3个不同的地方,长期生活在不同的海拔,适应了当地的气候条件,并成为地方特有品种。试验地海拔不同光照强度不同,温度不同,海拔1 000 m的地方光照充足,但气温较低;海拔2 000 m地方光照充足,气温较高;海拔3 000 m地方光照强烈,气温低,常年阴雨。将3个品种均放到3个不同海拔,马铃薯表现出不同的差异。
3.1 海拔梯度对马铃薯形态结构的影响
强光照使得马铃薯叶片的光合作用增强,产生更多的光合产物。低温会抑制呼吸作用,降低光合作用产物的消耗。在强光照、蒸腾作用下,水分能促进植物的运输作用。本试验研究表明,‘盈江小洋芋’的株高、分枝数和分枝长在海拔2 000 m处的数值最大,在海拔3 000 m处的数值较大,在海拔1 000 m处的数值最小,茎粗在海拔1 000 m处的数值最大,且随着海拔高度增加茎粗减小。‘剑川红’的株高、分枝数和分枝长在海拔2 000 m处的数值最大,在海拔3 000 m处的数值较大,在海拔1 000 m处的数值最小,茎粗在海拔1 000 m处的数值最大,且随着海拔高度增加茎粗减小。‘老家洋芋’的株高、分枝数、分枝长在海拔2 000 m处的数值最大,在海拔3 000 m处的数值较大,在海拔1 000 m处的数值最小,茎粗在海拔1 000 m处的数值最大,在海拔2 000和3 000 m处的差异不大。在3个海拔高度上,‘盈江小洋芋’的各项形态指标都是最低的,这是因为3处种植地的温度都达不到‘盈江小洋芋’的最佳适宜生长点。而‘剑川红’、‘老家洋芋’都适宜中高海拔的气温和光照,所以各项形态指标要较‘盈江小洋芋’好。在海拔2 000 m的试验地,光照强度、降雨量和温度都是最适合马铃薯生长的,所以3个品种在海拔2 000 m处的株高、分枝数以及分枝长都是最大的。在海拔1 000 m左右的试验地,年降雨量、温度和光照强度没有试验地二(海拔2 000 m)的高,虽然降雨量以及温度条件与试验地三(海拔3 000 m)处差异不大,但光照强度却没有试验地强,导致在试验地一的3个马铃薯品种的各项形态指标(除茎粗外)最差。
3.2 海拔梯度对马铃薯解剖结构的影响
植物叶片是植物进行光合作用的主要器官,是植物对环境变化比较敏感且可塑性较大的器官,环境的变化会导致植物叶片的长、宽和厚、薄的变化[5]。上表皮受光照强度不同有明显变化,强光有利于上表皮变厚。栅栏组织是叶片叶绿素含量较多的组织,在叶片中有栅栏组织和海绵组织分化的植物中,栅栏组织的厚度、栅栏组织细胞中叶绿体的分布常常决定于外界环境条件,特别是光照条件[12]。下表皮叶绿素含量少,但是含有大量自由水。下表皮分布着大量的气孔,控制植物气体和水分的交换。植物下表皮的厚度影响着植物气体和水分的交换[13]。本试验结果为‘盈江小洋芋’和‘剑川红’的上表皮厚度都是随着海拔的升高而增加,只有‘老家洋芋’是在海拔2 000 m处的上表皮厚度最大,但也是海拔3 000 m处的上表皮厚度高于海拔1 000 m。‘剑川红’和‘老家洋芋’的下表皮厚度都是随着海拔的升高而增加,只有‘盈江小洋芋’是在海拔2 000 m处的下表皮厚度最大,但也是海拔3 000 m处的下表皮厚度高于海拔1 000 m处,说明强光照会使表皮厚度增加。
‘盈江小洋芋’和‘老家洋芋’的叶片栅栏组织厚度均是在海拔3 000 m处最大,二者在海拔1 000 m处的栅栏组织厚度又较海拔2 000 m处高;‘剑川红’在3个海拔的叶片栅栏组织厚度差异不大。3个马铃薯品种在海拔3 000 m处的海绵组织厚度最大,且均是在海拔1 000 m处的海绵组织厚度大于海拔2 000 m。因为光照是影响栅栏组织和海绵组织厚度的重要因素,而光照强度随海拔升高而升高,由此可以基本看出海拔越高栅栏组织厚度和海绵组织厚度随着海拔的升高而增大,但这3个马铃薯地方品种出自于3个不同的地方,引入试验地后许多错综复杂的生长因子使这些被引种而来的马铃薯在生长过程中被迫表现出了各自的特异性来满足自身生长的需求,从而出现了‘剑川红’在3个海拔的栅栏组织厚度差异不大,出现‘盈江小洋芋’和‘老家洋芋’的栅栏组织和3个马铃薯品种的海绵组织厚度在海拔1 000 m处大于海拔2 000 m处的现象。
叶是植物进行光合作用和蒸腾作用的主要器官,具有较大的表面积,与周围环境的接触十分紧密,植物对环境的反应较多地体现在叶的形态结构上[14]。叶片的气孔数多少和气孔开度的大小能反应出马铃薯的光合作用和呼吸作用的强弱[15]。试验研究结果表明,马铃薯的3个品种都是在海拔1 000 m处的气孔开度最大,除‘老家洋芋’外其他2个品种在海拔2 000和3 000 m处的差异不大。除‘老家洋芋’外,其他2个品种的气孔数都是随着海拔升高而升高的,‘老家洋芋’在海拔2 000 m处的气孔数是最多的,海拔1 000 m处的气孔数较海拔3 000 m处的气孔数多。‘老家洋芋’的气孔开度和气孔数表现出与其他2个品种不同的特异性,可能是为了满足自身更好生长的结果。一定范围内气孔数越多,气孔开度越大越好,气孔数目多蒸腾速度快,有利于水分和矿质元素的运输。从光合作用角度讲,气孔密度大,单位时间单位面积上吸收的CO2的量多,同化CO2的量也随之增加,光合作用速度加快,形成的有机物就多。
总而言之,随着海拔升高,光照强度变大,马铃薯的形态结构、叶片厚度向适宜自身生长的方向发展,在3个不同的海拔地区,3个地方品种的株高、分枝数、分枝长都是在海拔2 000 m的地方最为良好,茎粗则是在海拔1 000 m的地方生长最为良好,这也表明了光照强度不同,马铃薯自身的生长激素的分配不同。3个马铃薯地方品种的自身组织结构随着海拔的升高而存在着一定的规律性却又向着有利于生长方向发展而存在着特异性,例如‘老家洋芋’的气孔开度和气孔数表现出与其他2个品种不同的特异性等都是品种自身形成的适应环境的结果。
[1]吴长松,刘红梅,龙玲,等.对黔西北高海拔山区优质杂交玉米制种基地产地检疫思考[J].中国种业,2013(10):12-14.
[2]桑月秋,杨琼芬,刘彦和,等.云南省马铃薯种植区域分布和周年生产[J].西南农业学报,2014,27(3):1003-1008.
[3]郑殿升,游承俐,高爱农,等.云南及周边地区少数民族对农业生物资源的保护与利用[J].植物遗传资源学报,2012,13(5): 699-703.
[4]和顺荣,木德伟,王建忠,等.马铃薯晚疫病避病栽培技术及在迪庆州应用[J].西南农业学报,2010,23(4):1360-1365.
[5]周绍松,李永梅,周敏.作物种间和品种间水分利用效率差异研究进展[J].云南农业大学学报,2007,22(5):736-740.
[6]唐探,姜永雷,冯程程,等.滇西北不同海拔梯度下急尖长苞冷杉叶片的生态解剖结构特性研究[J].江西农业大学学报,2015, 37(2):218-224.
[7]张春影,王艳,李美善,等.长白山不同海拔牛皮杜鹃叶片解剖结构的比较分析[J].延边大学农学学报,2014,36(1):39-43.
[8]沈宝云,刘星,王蒂,等.甘肃省中部沿黄灌区连作对马铃薯植株生理生态特性的影响[J].中国生态农业学报,2013,21(6): 689-699.
[9]苗百岭,侯琼,梁存柱.基于GIS的阴山旱作区马铃薯种植农业气候区划[J].应用生态学报,2015,26(1):278-282.
[10]宁海龙.田间试验数据的计算机分析[M].北京:科学出版社, 2012:25-29.
[11]桂克印,李炎林,唐前瑞,等.不同光处理下绿萝叶片及叶片解剖结构[J].吉首大学学报:自然科学版,2007(5):95-98,102.
[12]陈豫梅,陈厚彬,陈国菊,等.橡胶叶片心态结构与抗旱性的关系的研究[J].热带农业科学,2001,21(4):14-16.
[13]郑顺林,杨世民,李世林,等.氮肥水平对马铃薯光合及叶绿素荧光特性的影响[J].西南大学学报:自然科学版,2013,35(1): 1-9.
[14]柯曼琴.茎尖脱毒马铃薯植株的形态解剖学研究[J].曲阜师范大学学报,1996,22(4):86-88.
[15]陈晨,张永成.马铃薯不同品种间气孔密度及叶绿素含量的差异性研究[J].中国农学通报,2013,29(27):83-87.
辰翔矿业有限公司
专业生产马铃薯育种膨胀蛭石
河北灵寿县辰翔矿业有限公司位于河北省石家庄市灵寿县,是一家专业生产蛭石片、膨胀蛭石、珍珠岩的企业,已有30多年的发展历史。辰翔公司根据马铃薯育种特点,研发了育种专用膨胀蛭石。本公司生产的马铃薯专用膨胀蛭石性价比高,已在国内十几家马铃薯育种公司应用,并得到一致好评。本公司蛭石产品型号齐全,也可根据客户需求订制生产。
如果您对我们的产品感兴趣,欢迎致电联系,索要资料、样品。
联系人:薛刚15613123526、15833992815
地址:河北省石家庄市灵寿县燕川工业区
电话:0311-82616100(传真)
Effects of Elevations on Morphological and Anatomical Structure of Local Potato Varieties
WANG Wenxiang1,DABU Xilatu2,ZHOU Ping1,SHA Bencai1,GENG Shiying1,LIU Jia1,HAI Meirong1*
(1.College of Agronomy and Biotechnology,Yunnan Agricultural University,Kunming,Yunnan 650201,China; 2.College of Resources and Environment,Yunnan Agricultural University,Kunming,Yunnan 650201,China)
ract:To study the influence of altitudinal gradient on potato morphological and anatomical structure of local varieties,morphological and anatomical structural features of three local varieties were measured and analyzed in a field experiment.There were some patterns in the organizational structure of three local potato varieties with the increase of altitude while there existed in specificity conductive to the development of growth direction.The potato at 2 000 m possessed numerical maximum in plant height,branch length and branch number of these three varieties and stem diameter at 1 000 m.Thickness of upper and lower epidermis of these three potato varieties were increasing with the rise of altitude(except for the lower epidermis thickness of'Yingjiang small potato',and upper epidermis thickness of'Native potato').The maximal leaf palisade tissue thickness of'Yingjiang small potato'and'Native potato'were all at an altitude of 3 000 m,the leaf palisade tissue thickness of'Jianchuanhong'in the three altitudes had no difference.The potato at 3 000 m possessed maximal sponge tissue thickness in these three varieties.The potato at 1 000 m possessed maximal stomatal conductance and the potato at 2 000 m possessed minimal value in these three varieties.Except for'Native potato',the stomata number of the other two varieties were increasing with the rise of altitude,however,the stomata number of'Native potato'possessed maximal value at 2 000 m and minimal value at 3 000 m.
rds:potato landrace;altitudinal gradient;morphological structure;anatomical structure
S532
A
1672-3635(2017)02-0077-09
2016-03-24
国家自然科学基金青年基金(31201147);云南省现代农业马铃薯产业技术体系建设栽培研究室岗位专家专项经费(云财教[2014]160号)。
王文祥(1991-),男,硕士研究生,主要从事植物生产生理研究。
海梅荣,副教授,主要从事农作物生产生理研究,E-mail:2250029499@qq.com。
猜你喜欢
云南画报(2021年6期)2021-07-28 07:10:48
云南画报(2021年5期)2021-07-22 08:45:18
学苑创造·A版(2020年5期)2020-07-04 12:35:46
学生天地(2020年19期)2020-06-01 02:11:30
民族音乐(2019年3期)2019-08-14 01:05:18
Asian Herpetological Research(2019年1期)2019-03-27 12:47:44
现代园艺(2018年2期)2018-03-15 08:00:05
特别健康(2017年9期)2017-11-06 00:32:43
中国糖料(2016年1期)2016-12-01 06:49:04
中国音乐教育(2014年3期)2014-05-16 03:38:40
