
云母晶面上准外延生长的胶原蛋白纳米线阵列的性质研究
2017-04-14 13:08杨德良曾凡喜孙铭顾文华李力
分析化学 2017年4期
杨德良+曾凡喜+孙铭+顾文华+李力


摘要 胶原蛋白纤维与同样具有周期性结构表面的羟磷灰石等矿物质晶体能够通过多种作用在介观尺度上相互识别,从而有效地进行规范骨骼、牙齿等器官生长的生物矿化。在本研究中,反相利用生物矿化原理,开发出一种与热壁外延生长(Hot wall epitaxy)技术效果相似的生物大分子纳米线阵列的简单制备技术,成功地将5~10 μg/mL的天然I型鼠尾胶原蛋白单体溶液在云母晶体的(001)晶面上培育成取向单一且长程有序的胶原蛋白纳米线阵列。原子力显微镜实验结果表明,纳米线阵列随胶原蛋白单体浓度增大而变得致密,但是单条纳米线的宽度与高度较为稳定,分别约为60.0和1.5 nm。有纳米线覆盖的云母晶面亲水性好,晶面接触角由25.8°降为9.5°。基于电子背面散射衍射和透射电子显微镜的分析表征结果,纳米线的取向应沿云母\[1〖TX-10\]方向, 更详实地验证了胶原蛋白纳米线的准外延生长机理。
关键词 胶原蛋白; 外延生长; 生物矿化; 纳米线; 电子衍射
1引言
外延生长(Epitaxial growth)常指一种矿物质晶体在另一种矿物质晶体表面生长,因而在界面上两种矿物质具有相同晶体结构取向的现象[1~5]。半个多世纪以来,利用外延生长原理在晶体基质表面控制和研究镀膜分子的膜层结构以及生长动力学已成为热门研究领域,并取得了诸多成果[1~13]。二十世纪中后期多种商品化无机发光二极管便是利用液相外延生长技术制备的[6]。近二十年来,结合有机化学中的尺寸与功能可调的分子合成技术, 以及各种微观晶体结构的基质选择,在超高真空背景下利用分子束沉积技术在基质表面控制分子膜层晶体的生长,为获得功能可预测的新型微纳光电材料与器件提供了多种可能性[7~9]。例如,通过热壁外延生长(Hot wall epitaxy)技术将对六联苯(pSexiphenyl)等高量子效率的蓝光小分子材料在云母、ITO等基质表面生成有序的纳米线阵列薄膜[10~13], 由于长程有序,表现出很强的光学各向异性。该薄膜还能产生光学增益, 获得随机激光,可用于制备有偏振特性的电致发光器件[12,13]。
外延生长在生物化学领域并不多见。但在自然界中,有规则重复结构的胶原蛋白纤维能够在介观尺度上与矿物晶体相互识别,从而给生物矿化提供支架,有效地调控矿物晶体生长的启动与阻断,在尺寸与取向方面规范骨骼、牙齿等器官的生长[14,15]。另一方面,反相利用矿物晶体与生物大分子的识别作用,在缓冲溶液中控制无机晶体表面的生物大分子自组装也同样具有可行性[16~19]。这种生物大分子在基质表面的外延生长对深入研究生物大分子功能,合成新型微纳材料,以及制备特异高效的生物探针都具有重要意义[20~22]。
本研究以天然I型鼠尾胶原蛋白单体和云母晶体为原料,开发了一种生物大分子在电荷周期性分布的晶体表面通过液相外延生长来合成纳米线阵列的“反相生物矿化”技术。此技术应用方便,无需定向流动的缓冲液培养[16]或复杂缓冲液的配制[17~19]。基于各种成像和分析手段的表征结果,对合成的胶原蛋白纳米线阵列的性质进行了初步探讨。
2实验部分
2.1仪器与试剂
Bruker Dimension Icon原子力显微镜和FESP悬臂探针(德国布鲁克公司), 探针为镀金和铬的硅材质,悬臂与针尖的力常数均为2.8 N/m,针尖高15 μm,锥角25°,曲率半径12 nm; Kruss DSA100液滴形状分析仪(德国Kruss公司); Auriga FIB聚焦离子束场发射扫描双束电镜(德国Zeiss公司); 空间分辨率50 nm。FEI Tecnai G2 20 LaB6高分辨透射电镜(美国FEI公司)。
5 mg/mL胶原蛋白单体溶液(Rat tail tendon collagen type I,北京索莱宝科技有限公司),溶于6 mmol/L乙酸; Na2HPO4、NaH2PO4、K2HPO4、KH2PO4(CP,国药集团化学试剂有限公司)。2M1 muscovite云母基片(北京中镜科仪技术有限公司)。单面与双面胶带(美国3M公司)。实验用水为自制去离子水,室温电阻率约18 MΩ·cm。
2.2實验方法
使用磷酸盐缓冲溶液配制特定浓度的胶原蛋白单体溶液。将云母片切割成约1 cm×1 cm的方片,用双面胶带粘在载玻片中央,用单面胶带撕开云母片表层, 使其露出新鲜晶面(001),滴上胶原蛋白单体溶液覆盖晶面约15 min后, 除去晶面上的蛋白溶液,并用缓冲溶液清洗晶面1~2次,以除去吸附不牢的蛋白质,接着用缓冲溶液覆盖云母晶面,进行约12 h的培育。完成培育后立即除去晶面上的缓冲溶液,用电热吹风烘干后对晶面上的膜层进行表征。
原子力显微镜设置为轻敲模式(Tapping mode),振幅约75 nm,驱动频率约192 kHz,接近探针悬臂在空气中的共振频率。液滴形状分析仪进行静态接触角测试前将样品置于真空干燥箱内脱气30 min。透射电镜/选区衍射(TEM/SAED)实验前将样品撕薄(<100 nm)后打孔成直径3 mm的样品;在TEM模式下检测到纳米线结构后,切换到NBD(Nanobeam electron Diffraction)模式对定点获取电子衍射花样,然后进行标定分析。电子背面散射衍射(EBSD)实验前,对样品表面进行镀金处理(厚度约5 nm)。EBSD设置:加速电压20 kV,工作距离13 mm。
3结果与讨论
3.1胶原蛋白单体在云母基质上的自组装
胶原蛋白是构成所有多细胞生物结缔组织的主要组成成分的蛋白质的总称。作为最重要的细胞外基质(ECM)蛋白,胶原蛋白单体能自组装成胶原纤维来构架并支撑所有无脊椎与脊椎动物的组织器官[14]。I型胶原蛋白最常见,其单体原胶原(Tropocollagen)为绳状的超螺旋分子,长约300 nm,直径约1.5 nm。尽管原胶原单股螺旋的周期为85.5 ,但超螺旋结构沿轴向的残基与电荷分布周期约为8.6 。在末端端肽(Telopeptide)的引导下,原胶原单体会互相错开平行排列自组装成细长的原纤维(Protofibril)。在生物矿化过程中,羟基磷灰石会首先在原纤维的特定位点结晶,并逐渐生长成带状小块,最后形成高度有序的交错结构。
本实验分别使用不同pH值的Na2HPO4/NaH2PO4和K2HPO4/KH2PO4缓冲溶液配制I型鼠尾胶原蛋白单体溶液,并用同种缓冲溶液来培育胶蛋白自组装。在pH 6.8~8.0的范围内,钠盐溶液中的胶原蛋白单体均能在云母晶体表面自组装成取向单一的蛋白纤维纳米线阵列。图1显示的是用不同浓度的胶原蛋白单体在云母晶体(001)晶面上自组装形成的纳米线阵列的AFM图。缓冲溶液均为Na2HPO4/NaH2PO4 (pH =7.5),其中图1A的胶原蛋白单体浓度为5 μg/mL,图1B的胶原蛋白单体浓度为10 μg/mL。 很显然,阵列中的纳米线随胶原蛋白单体浓度增大而变得更加致密,单条纳米线更长,且纳米线之间的交联逐渐显著。但是测量结果表明,由不同浓度的胶原蛋白单体溶液培育出的单条纳米线的宽度与高度较为稳定,宽度约为60 nm,高度约为1.5 nm。当使用钾盐缓冲溶液时,未在云母晶体表面上观测到任何自组装现象发生。
3.2胶原蛋白纳米纤维阵列的特性
分别对新撕开的和已有胶原蛋白纳米线膜层覆盖的云母(001)晶面样品进行了静态水滴接触角的基线圆法测试。为减小测量误差,对同一样品取3个点进行测量后取平均值。测量结果表明(见电子版文后支持信息: 图S1),云母晶面接触角为25.8°,而有蛋白纳米线覆盖(10 μg/mL)的晶面接触角仅为9.5°。上述结果表明,胶原蛋白纳米线膜层显著增强了云母晶面的亲水性,有利于使用该材料生产细胞培养器皿。
采用“反相生物矿化”技术制备出的胶原蛋白纳米线阵列与通过热壁外延生长技术生成的对六联苯(pSexiphenyl)纳米线阵列[10,11]极为相似:(1)纳米线受云母基质表面电荷分布影响,取向单一,且纳米线阵列结构长程有序;(2)纳米线密度由云母晶体表面吸附(沉积)的单体分子的数量决定。
3.3胶原蛋白纳米线阵列的取向机理
云母基片上的胶原蛋白纳米线阵列的形成可能受培养溶液的流动方向规导[16]或受溶液水分挥发过程中的致干效应的影响[17], 但是更倾向于接受“反相生物矿化”原理下的准外延生长现象[18,19]。根据外延生长理论,有两个因素决定外延晶体的结构:(1)界面上的结合力(包括化学键、静电力与范德华力);(2)界面上的晶格匹配[7]。在真空环境下,有机分子在无机晶体表面的外延生长受控于较弱的静电力与范德华力,在对基质加热时, 有机分子可在基质表面漂移;尽管有机分子与无机基质的晶格可能不匹配,但基质表面的电荷分布仍能对有机分子的取向起规范作用。因而,在热壁外延生长实验中,取向较为规范的有机分子单体在基质晶面上漂移时,碰撞结合形成晶种,并生长成更大的二维晶体,甚至形成有特定取向的纳米线阵列。由于纳米线的形成是单体分子在二维晶面上侧向结合所致,因此单体浓度在一定范围内,纳米线有恒定高度和宽度。尽管本实验中培养胶原蛋白的液相环境可能更为复杂,〖JP2但是,在界面上水分子与吸附在云母晶面上的单层胶原蛋白单体之间最重要的作用力,即熵力(Entropy force),也倾向于推动蛋白单体在云母晶面上漂移,从而有助于自组装生成納米线阵列。同时,缓冲溶液中的离子成分对纳米线阵列的形成至关重要,使用钾盐缓冲溶液的样品则观测不到胶原蛋白纳米线。这可能是特定的离子会影响基质与蛋白〖JP单体在界面的结合,或对胶原蛋白的成纤有较强的抑制所致[18]。
目前, 尚未有详实和精准的晶体结构表征实验来确证胶原蛋白纳米线阵列的外延生长机理, 本研究首先对覆盖纳米纤维的云母基片进行了TEM的选区衍射(SAED)实验。通过对衍射光斑的标定与坐标分析,纳米线的取向大致与云母\[001\]晶轴成60°夹角(见电子版文后支持信息:图S2),这与文献\[18\]报道Laue衍射实验结果一致。此外,在云母晶面热壁外延生长的对六联苯纳米线阵列也和云母\[001\]晶轴大致成60°夹角(通过\[1 2〖TX- 1〖TX-\]psp//\[ 3〖TX- 4 0\]mica推导出PSP纳米线//\[1〖TX- 1 0\]mica)[10,11],表明这两种截然不同的生长环境下纳米线生长机理的相似性。
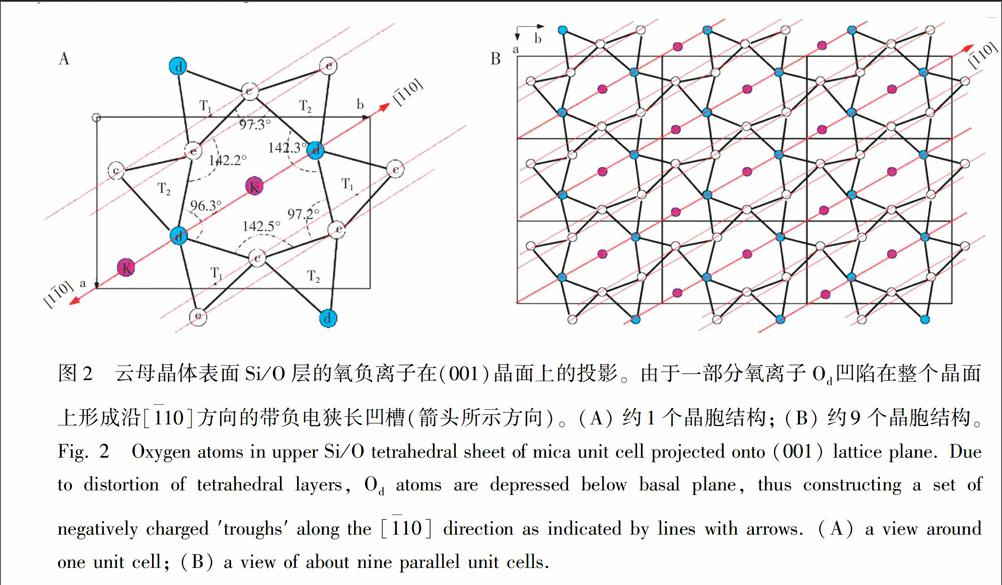
本实验中所用的基质云母(Mica, 2M1 muscovite)组成大致为{KAl2(AlSi3O10)(OH)2},属于单斜晶系(a=5.2906 , b=9.0080 , c=20.0470 , β=95.757°), 晶胞是互成120°的双层结构[23]。云母片可以层层撕开而露出与前一层原子排列成120°的崭新晶面。如图2A所示,在每个单层里,中心的Al与O以八面体结构组合并被表面的Si/O层包夹。(001)晶面的Si/O以四面体结构连接形成六边形的O2
Symbolm@@ 空洞,容纳层间的K+。在溶液中,晶体表面的K+可被其它离子或带电基团取代。由于Si/O四面体结构的变形导致一部分氧离子凹陷下去0.22 ,使得氧离子空洞呈不规则六边形,并在整个晶面上形成如图2B所示沿\[1〖TX-10\]方向,与\[001\]晶轴成60°的带负电狭长凹槽,这也是(001)晶面上电荷分布的二次对称轴。此对称轴使得唯有沿\[1〖TX-10\]方向或垂直于\[1〖TX-10\]方向的电荷分布是唯一的。根据此对称性以及观测到胶原蛋白纳米线阵列取向的单一性,可以推断纳米线的取向或者平行于该狭长氧负离子凹槽,或者垂直于该方向。
对镀金处理的样品表面(镀金厚度约5 nm)进行的EBSD实验也许可以进一步确定纳米线的取向。图3为某特定样品选区SEM图像及该区背散射电子衍射花纹图。基于对该区多点衍射花纹进行标定和坐标分析,纳米线的取向却大致与云母\[001\]晶轴成0°。这与纳米线取向沿云母\[1〖TX-10\]方向并不矛盾:可能是制样过程中对云母晶面造成破坏,使下层扭转120°的晶面露出所致(图3C)。如果垂直于\[1〖TX-10\]方向,纳米纤维则不可能与云母\[001\]晶轴平行。基于理论推导和实验结果,以及在云母晶面上通过热壁外延生长的对六联苯纳米线的取向(PSP纳米线//\[1〖TX-10\]mica)[10,11], 之前研究报道胶原蛋白在云母晶面沿\[110\]方向生长的结果需进一步验证[18]。
4結 论
利用“反相生物矿化”原理,开发出一种可与热壁外延生长技术效果媲美,但实验操作更简便的生物大分子纳米线阵列的制备方法, 无需超净室或真空环境,所有操作均在室温下进行,不需对基质晶体进行加热。 此方法生成的蛋白纳米纤维阵列结构可控,为生产细胞培养器皿、制备高特异性的生物探针,以及合成新型有机微纳光电材料与器件提供了一种可行技术。
References
1Ward M D. ACS Nano, 2016, 10(7): 6424-6428
2YU WeiHua, SHENG JuanJuan. Chinese J. Anal. Chem., 1986, 14(3): 239-239
俞薇华, 盛娟娟. 分析化学, 1986, 14(3): 239-239
3SUN Peng, WANG Chao, YUAN Guang, LI JunJie, GU ChangZhi. Chinese J. Lumin., 2016, 37(7): 793-797
孙 鹏, 王 超, 元 光, 李俊杰, 顾长志. 发光学报, 2016, 37(7): 793-797
4ZHANG QuanLin, SU LongXing, WU TianZhun, WANG YuChao, ZHU Yuan, CHEN MingMing, GUI XuChun, TANG ZiKang. Chinese J. Lumin., 2015, 36(10): 1171-1177
张权林, 苏龙兴, 吴天准, 王玉超, 祝 渊, 陈明明, 桂许春, 汤子康. 发光学报, 2015, 36(10): 1171-1177
5Lim S H, Ko Y H, Rodriguez C, Gong S H, Cho Y H. Light: Sci. Appl., 2016, 5(2): e16030
6Nelson H. J. Cryst. Growth, 1974, 27: 1-5
7Tersoff J, Tromp R M. Phys. Rev. Lett., 1993, 70(18): 2782-2785
8Shekhah O, Wang H, Paradinas M, Ocal C, Schüpbach B, Terfort A, Zacher D, Fischer R A, Woll C. Nat. Mater., 2009, 8(6): 481-484
9Jesse S, Borisevich A Y, Fowlkes J D, Lupini A R, Rack P D, Unocic R R, Sumpter B G, Kalinin S V, Belianinov A, Ovchinnikova O S. ACS Nano, 2016, 10(6): 5600-5618
10Plank H, Resel R, Purger S, Keckes J, Thierry A, Lotz B, Andreev A, Sariciftci N S, Sitter H. Phys. Rev. B, 2001, 64(23): 235423
11Plank H, Resel R, Andreev A, Sariciftci N S, Sitter H. J. Cryst. Growth, 2002, 237239(3): 2076-2081
12Zenc C, Cerullo G, Lanzani G, Graupner W, Meghdadi F. Phys. Rev. B, 1999, 59(22): 14336-14341
13Andreev A, Quochi F, Cordella F, Mura A, Bongiovanni G, Sitter H, Hlawacek G, Teichert C, Sariciftci N S. J. Appl. Phys., 2006, 99(3): 034305
14Puleo D A, Bizios R. Biological Interactions on Materials Surfaces: Understanding and Controlling Protein, Cell, and Tissue Responses, Springer Science, Springer, 2009, pp. 200-218
15Ihli J, Clark J N, Ct A S, Kim Y Y, Schenk A S, Kulak A N, Comyn T P, Chammas O, Harder R J, Duffy D M, Robinson I K, Meldrum F C. Nat. Commun., 2016, 7: 13141
16Jiang F, Horber H, Howard J, Muller D J. J. Struct. Biol., 2004, 148: 268-278
17Bozec L, Horton M. Biophys. J., 2005, 88(6): 4223-4231
18Sun M, Stetco A, Merschrod E F. Langmuir, 2008, 24(10): 5418-5421
19Leow W W, Hwang W. Langmuir, 2011, 27(17): 10907-10913
20Xiao G, Zhou J F, Huang X, Liao X P, Shi B. RSC Adv., 2014, 4(8): 4010-4019
21Ghosal K, Thomas S, Kalarikkal N, Gnanamani A. J. Polym. Res., 2014, 21(5): 410
22Li P P, Chen X, Yang W S. Langmuir, 2013, 29(27): 8629-8635
23Guven N Z. Zeitschrift für KristallographieGrystalline Mater., 1971, 134: 196-212
AbstractCollagen fibrils and hydroxyapatite might recognize each other at the mesoscale by multiple cooperative interactions due to their intrinsically repetitive structured surfaces, and thus effectively directing the biomineralization, a biological process involving regulating the growth of bones, teeth and other organs. In this work, we developed a simple technique to prepare nanowire arrays of biological macromolecules by reversely using the biomineralization mechanism, with results similar to the hot wall epitaxy, a molecular beam deposition technique under vacuum. With this technique, we successfully cultured 5-10 μg/mL rat tail type I collagen monomer solutions into collagen nanowire arrays on the mica (001) lattice plane along one unique direction across the whole cleavage surface. The atomic force microscope experiments indicated that the nanowires in the arrays became more crowded with higher monomer concentration, but their width and height remained unchanged, about 60.0 nm and 1.5 nm, respectively. The collagen nanowire coating enhanced the hydrophilicity of the mica surface, reducing the contact angle from 25.8° to 9.5°. Based on the characterization results of electron back scattering diffraction and transmission electron microscope, the collagen nanowires were most likely to be oriented along the mica \[1〖TX-10\] direction, which validated the quasiepitaxial growth mechanism in more details.
KeywordsCollagen; Epitaxial growth; Biomineralization; Nanowire; Electron diffraction
(Received 3 November 2016; accepted 1 December 2016)
The work was supported by the National Natural Science Foundation of China (No. 51303082)
猜你喜欢
消费者报道(2019年6期)2019-08-17
分析化学(2018年12期)2018-01-22
Coco薇(2017年12期)2018-01-03
饮食与健康·下旬刊(2017年3期)2017-03-30
哈尔滨理工大学学报(2016年4期)2016-11-10
中国高新技术企业(2015年24期)2015-06-25
饮食科学(2009年12期)2009-12-11
家庭医学·下半月(2009年5期)2009-07-16
饮食科学(2009年2期)2009-02-26
