
半干旱区湿地—草原生态交错带边界判定及其群落多样性分析
——以宁夏四儿滩湿地为例
2017-04-10 00:32:04苏香玲王振锡
林业资源管理 2017年1期
苏香玲,王振锡
(1.新疆林业规划院,乌鲁木齐830049;2.新疆农业大学林学与园艺学院,乌鲁木齐830052;3.新疆教育厅干旱区林业生态与产业技术重点实验室,乌鲁木齐830052)
半干旱区湿地—草原生态交错带边界判定及其群落多样性分析
——以宁夏四儿滩湿地为例
苏香玲1,王振锡2,3
(1.新疆林业规划院,乌鲁木齐830049;2.新疆农业大学林学与园艺学院,乌鲁木齐830052;3.新疆教育厅干旱区林业生态与产业技术重点实验室,乌鲁木齐830052)
为探究半干旱区湿地—草原生态交错带的边界宽度及其群落多样性,于2014年7月对宁夏盐池县四儿滩湿地进行野外植被调查。利用移动分割窗技术对交错带进行定量判定,采用Shannon-Wiener指数、Simpson指数、Pielou均匀度指数、Margalef物种丰富度指数分析不同生境带以及不同样线间的植物群落多样性。结果表明:1)窗口宽度为6个样方时,能够清晰地划定此交错带的边界。东、西、南、北样线生态交错带的宽度分别为120,120,120,140m,距湿地核心区距离分别为180,220,240,200m。2)不同生境带Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef物种丰富度指数的排列顺序均为交错带>旱生带>湿生带,而 Simpson优势度指数的排列顺序为旱生带>交错带>湿生带。3)北样线的Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数均为最高值,而西样线4多样性指数均为最低值。研究结果可为干旱、半干旱地区湿地资源的保护、生物多样性的维持、生态系统的稳定提供理论支持和科学依据。
生态交错带;移动分割窗技术;群落多样性
生态交错带最早由Clements提出,他认为生态交错带是由气候决定的植物群丛交迭的应力区,主要包括了边缘、树线和群落交错带[1]。生态交错带是独特的环境单元[2],是生态系统中基本的景观单元[3],具有高物种多样性、丰富的特有种、大量的外来种、频繁的物质流动、敏感的时空动态性、结构的异质性和脆弱性。目前,国内外学者对不同类型的生态交错带进行了大量深入的研究。对农林生态系统生态交错带的研究发现:退耕撂荒地—云南松林交错带宽度约9m[4]。对森林-沼泽生态交错带的研究表明:森林—沼泽生态交错带植物多样性、群落结构特征、群落生产力均随着交错带环境梯度的变化而呈现有规律的分布格局,沿着沼泽至森林方向的交错区环境梯度,群落建群种发生更替现象[5]。贾宝全等对绿洲—荒漠交错带研究发现:由于生态交错带边缘效应的存在,交错带的植被盖度较高[6]。对北方农牧交错带研究表明:生态缺水是造成北方农牧交错带植被生态系统退化的主要原因[7]。
虽然国内对不同类型生态交错带报道较多,但对于湿地—草原交错带研究较少,特别是湿地—草原生态交错带的定量判定。本研究中湿地—草原生态系统位于宁夏盐池县四儿滩湿地,隶属于哈巴湖荒漠湿地国家级自然保护区,是保护区内湖泊面积较大的一块湿地。研究湿地—草原生态交错带边界判定及其群落多样性的变化过程,为干旱、半干旱地区宝贵湿地资源的保护、生物多样性的维持、生态系统的稳定提供理论支持和科学依据。
1 研究方法
1.1 研究区概况
四儿滩湿地位于盐池县南部,北纬37°43',东经107°24',面积大约为 1 420hm2,海拔为 1 320~1 370m。湿地水面面积主要受降水影响,具有季节性积水特征。湿地—草原交错带土壤含水量高,利于植被生长,植物种类丰富,包含了所有耐湿、耐盐碱的沼泽植物,例如:盐爪爪(Kalidium foliatum)、碱蓬(Suaeda glauca),等;还包含了耐干旱的草原植被,例如:刺沙蓬(Salsola ruthenica)、猪毛蒿(Artemisia scoparia)等;同时还有交错带的特有植物,例如海乳草(Glaux maritima)、草瑞香(Diarthron linifolium)等植物。
1.2 外业调查
外业调查于2014年7月采用样线与样方结合的方法进行野外植被调查。从湿地中央向东、西、南、北4个方向辐射4条样线(图1),每条样线的起点为湿地核心区固定的水泥桩,沿样线每隔20m随机布设1m×1m的样方(图2),并且依次对样方进行编号,直至旱生带植被群落结构不发生变化时停止布设样方。每条样线跨越湿生带、整个交错带及典型旱生带(图3)。样方内记录植物名称、植物种类、株数、高度、盖度、生物量等指标。
1.3 数据处理
使用Excel 2010计算植物物种重要值矩阵。利用游动分割窗技术对交错带进行定量判定。同时,采用Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数、Margalef物种丰富度指数计算植物群落多样性。

图1 样线布设示意图Fig.1 Distribution of sample lines

图2 样方布设示意图Fig.2 Sample layout

图3 理想状态下湿生带、交错带、旱生带划分示意图Fig.3 Distribution of wet region,ecotone,arid region in ideal status
1.3.1 重要值
重要值计算公式如下[8]:

式中:Ra为相对多度,RC为相对盖度,Rh为相对高度,Rb为相对生物量,Rf为相对频率,I为重要值。
1.3.2 多样性指数
结合研究的内容,植被群落多样性指数采用Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数、Margalef物种丰富度指数进行分析,其公式如下[9]:
Shannon-Wiener多样性指数:

Simpson优势度指数:

Pielou均匀度指数:

Margalef物种丰富度指数:

其中:Pi=Ni/N
式中:Ni为样方中第i种植物的重要值;N为样方植物重要值总和;H为Shannon-Wiener多样性指数;S为植物物种数。
1.3.3 边界确定方法
移动分割窗技术是利用多个(偶数)样方组成的窗体沿样带逐个样方滑动,计算2个1/2窗体间的相异系数,将所得的距离(相异)系数沿样带序列作图,根据陡峭峰值区间判定交错带的一种技术[10]。本文采用平方欧氏距离(D)计算窗体间的相异系数[11]:

式中:Dn为窗体为n时平方欧氏距离,X-ia和X-ib分别代表A半窗体和B半窗体在参数为i时的值,m为窗口变量数。
2 结果与分析
2.1 适应窗口宽度的选择
移动分割窗技术通过窗体平滑的方法避免了单纯计算样带相邻样方间相异系数产生的跌宕起伏的干扰噪声和难以判定交错带格局,能更客观地判定交错带的位置和宽度[12]。图4是以四儿滩湿地南样线上的植物重要值为指标,采用不同分割窗大小计算的欧氏距离在样带系列上的分布图。由图4(a)可知,当窗口宽度为4个样方时,平方欧氏距离出现了多个峰值,多波峰噪音干扰了交错带判定,交错带判定不准确。然而,当窗口宽度为6或者大于6个样方时,平方欧式距离变化趋于一致,说明窗口宽度为6个样方时,能够清晰地划定湿地—草原交错带的边界。由野外调查可知,湿生带(样方编号1—编号8)主要出现了盐爪爪、细枝盐爪爪(Kalidium gracile)等湿生植物,交错带(样方编号9—编号15)出现了芨芨草(Achnatherum splendens)、芦苇(Phragmites australis)、赖草(Leymus secalinus)、白刺(Nitraria sibirica)、臭蒿(Artemisia hedinii)、西伯利亚蓼(Polygonum sibiricum)等植物,这些植物多生长在地形较低、水分条件较好的低洼地、沼泽地,是典型的交错带植物。旱生带(样方编号16—编号21)主要出现了刺沙蓬、猪毛蒿、蒙古虫实(Corispermum mongolicum)、苦豆子(Sophora alopecuroides)等耐干旱的植物。这也说明游动分割窗技术能够准确判定交错带的位置。

图4 南样线不同窗口宽度的平方欧氏距离Fig.4 Squared Euclidean distance(SED)under different window width of south line transect
图5是四儿滩湿地东、西、北样线基于游动分割窗技术进行的生态交错带的判定,其结果也能较好地反应交错带的边界,且图中峰值较高,宽度较小,说明交错带过渡明显。
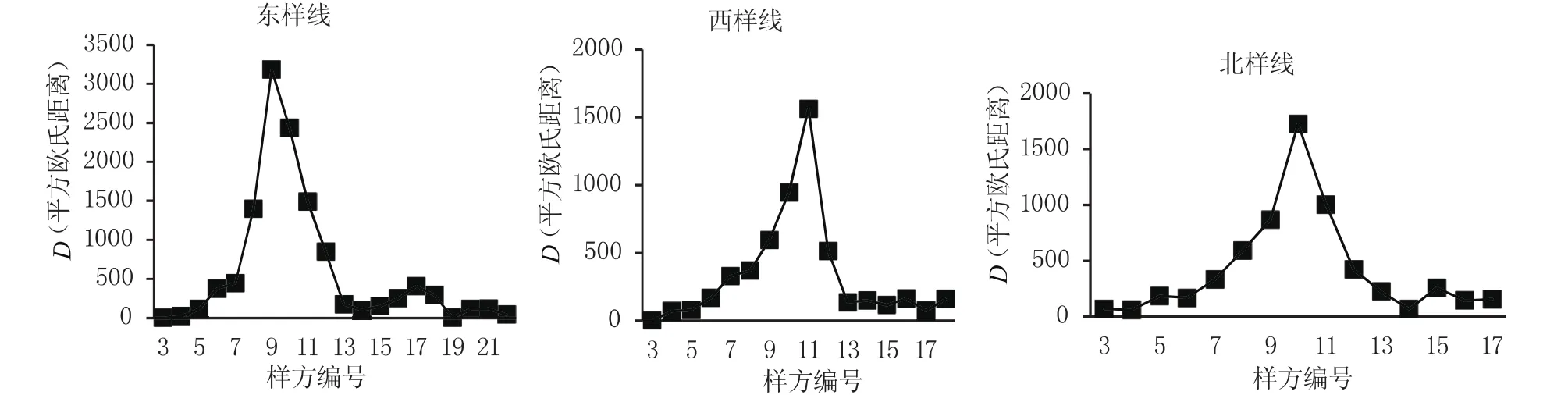
图5 东、西、北样线窗口宽度为6个样方的平方欧氏距离Fig.5 Squared Euclidean distance(SED)under six window width of east,west,north line transect
2.2 交错带边界和宽度的确定
用移动分割窗技术分析交错带的宽度时,陡峭的峰值出现区就交错带所在的位置,峰宽表示交错带的宽度,峰宽的端点即为交错带与相邻生态系统的边界[13]。本研究中,东样线的峰值出现在第9个样方,峰宽为样方编号7—编号13之间的距离,样方间间距为20m,所以东样线生态交错带的宽度为120m,生态交错带距湿地核心距离为180m(表1)。西样线生态交错带的宽度为120m,生态交错带距湿地核心距离为220m。南样线生态交错带的宽度为120m,生态交错带距湿地核心距离为240m。北样线生态交错带的宽度为140m,生态交错带距湿地核心距离为200m。不同样线的生态交错带宽度不同,生态交错带距湿地核心区的距离也不同,说明湿地的水域对植被的影响并不是等距离均一的向四周扩散的,而是表现出了方位上的差异性。
2.3 不同生态环境带间植物群落多样性分析
对东样线、西样线、南样线、北样线湿生带、交错带、旱生带进行植被群落多样性分析,结果见表2。
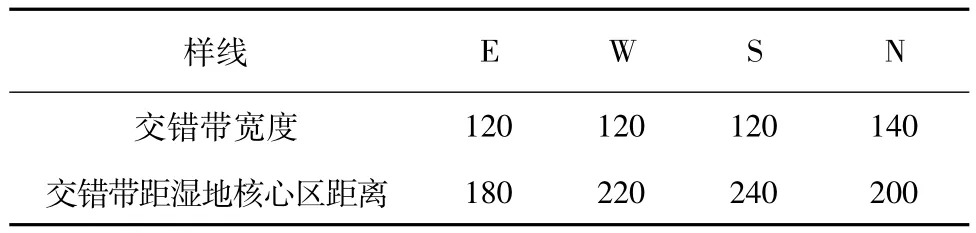
表1 不同样线生态交错带边界和宽度Tab.1 Ecotone boundary and width of different line transects m
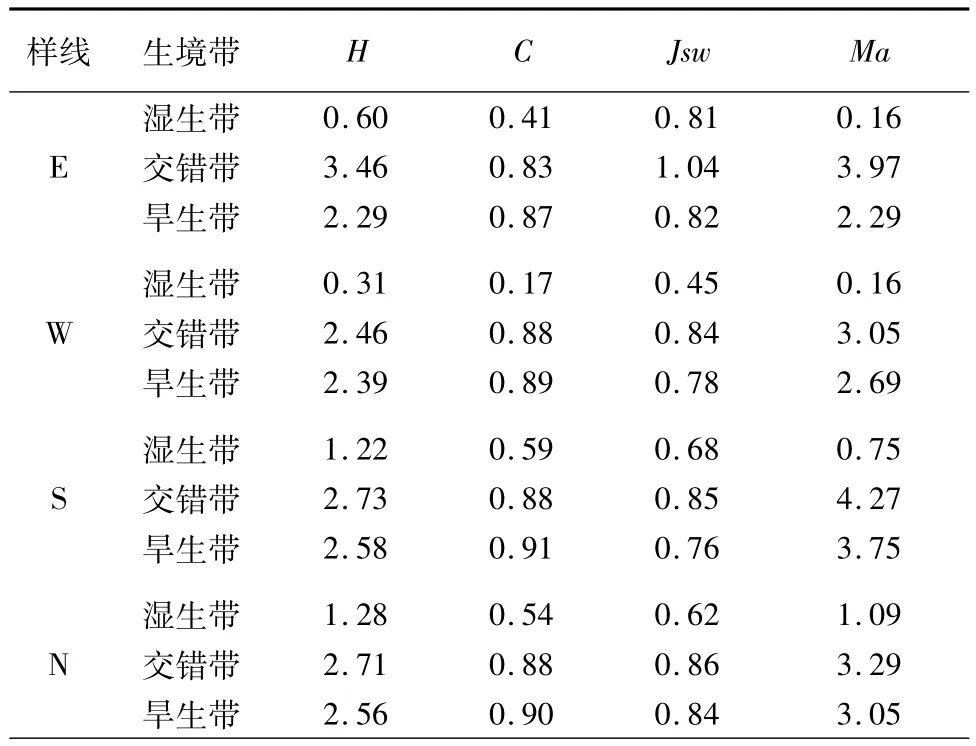
表2 四儿滩湿地不同生境带植物群落多样性Tab.2 Plant community diversity of different habitat regions in Siertan wetland
Shannon-Wiener指数综合了物种群落的均匀度和丰富度,是表述物种多样性最好的指数。由表2可知,4条样线Shannon-Wiener指数的排列顺序为:交错带>旱生带>湿生带。说明交错带物种多样性最丰富,群落组成复杂,群落稳定性高。而湿生带多样性指数最低,说明湿生带群落组成简单,物种多样性较低。这也从侧面印证了生态交错带的边缘效应,即生态交错带显示出较高的生物多样性。
生态优势度是群落水平上的综合数值,与每个种的重要值分布有关,其数值大小可以反映群落内各种群结构关系的复杂性[14]。由表2可知,4条样线的Simpson优势度指数的排列顺序为旱生带>交错带>湿生带,这说明旱生带优势种明显,而湿生带优势种不明显。
群落均匀度是表征群落种群空间分布均匀程度的重要指标。群落均匀度与群落优势度是两个相反的概念,该指标常作为群落物种多样性指数的辅助指标,在反映群落特征结构方面有重要意义。不同生境带均匀度指数最大的是交错带、其次是旱生带、再次是湿生带,说明交错带群落分布最均匀,湿生带群落分布最集中。
Margalef物种丰富度指数能直接反映物种的丰富程度,是植物群落内部或生境中物种个体数量多少的指标,其值越大,说明物种个体的丰富度越高,反之则说明物种个体的丰富度较低[15]。东、西、南、北样线交错带的丰富度分别为3.97,3.05,4.27,3.29,各样线交错带丰富度指数均为最高,说明交错带物种最为丰富,而4条样线中湿生带丰富度指数均小于交错带、旱生带,说明湿生带物种单一,多为耐湿、耐盐碱的小灌木。
2.4 不同样线间植物群落多样性分析
四儿滩湿地不同样线H指数、C指数、Jsw指数、Ma指数见表3。
从表3中可以看出不同样线方向湿地—草原交错带群落组成结构差异显著。H指数最大的是北样线,其次是南样线、再次是东样线、最后是西样线,H指数值分别为2.93,2.84,2.75,2.31。各样线C指数的排列顺序为北样线>南样线>东样线>西样线,其变化趋势与H指数一致。各样线均匀度指数Jsw的排列顺序为北样线>东样线>南样线>西样线,数值分别为0.82,0.80,0.78,0.71。丰富度指数最高的是南样线(4.84),其次是北样线(4.44),再次是东样线(3.83),最后是西样线(3.27)。北样线的H指数、C指数、Jsw指数均为最高值,说明北样线群落最为丰富,而西样线4个多样性指数均为最低值,说明西样线群落组成简单,物种多样性较低。

表3 四儿滩湿地各样线群落多样性Tab.3 Plant community diversity of different line transects in Siertan wetland
3 结论与讨论
1)采用移动分割窗技术分析四儿滩湿地—草原生态交错带的宽度,发现窗口宽度为6个样方时,能够清晰地划定此交错带的边界。东、西、南、北样线生态交错带的宽度分别为120,120,120,140m,距湿地核心区距离分别为180,220,240,200m。各样线生态交错带的宽度存在差异,其距湿地核心区距离也不同,说明湿地的水域对植被的影响并不是等距离均一地向四周扩散的,而是表现出了方位上的差异性。这种差异性是因为湿地周围的地势存在方位上的差别,这种地势高低的差异导致了四儿滩湿地的水域呈现东北—西南长,西北—东南短的形状。
2)通过对4条样线湿生带、交错带、旱生带植物群落多样性分析,发现Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef物种丰富度指数均为交错带>旱生带>湿生带;而Simpson优势度指数的排列顺序为旱生带>交错带>湿生带。这是因为盐池县自2002年实行全县禁牧以来,湿地受人为活动影响程度降低,植被恢复良好,植被演替进入了相对稳定的阶段,尤其是交错带,这就导致了交错带多样性指数、均匀度指数、丰富度指数均为最高。同时,旱生带位于湿地外围,在不同程度上受到人为活动的干扰,优势群落逐渐形成,所以此生境带优势度较高。
3)通过分析不同样线间植物群落多样性,发现不同样线方向湿地—草原交错带群落组成结构差异较大。北样线的 Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数均为最高值,而西样线的Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数、Margalef物种丰富度指数均为最低值。北样线物种丰富度最高,群落结构复杂且稳定性高,这是因为北样线离居民点较远,受人为干扰小,物质、能量运输速度快,群落盖度、生物量明显提高。西样线群落结构单一是因为离居民点较进,人为干扰(例如放牧、践踏等)强烈,植被被牲畜啃食、植被盖度、生物量大大减少,群落结构指标下降。
4)由于采用的是样方与样线的调查方法,四儿滩湿地样线的群落组成和多样性与样方布设有关。同时,湿地植被生长与降雨等环境因子存在显著关系,当环境条件恶劣时,植被的生长、分布等受到影响。这些影响了结果的准确性,在以后的工作中需要加以改进。
[1]朱芬萌,安树青,关保华,等.生态交错带及其研究进展[J].生态学报,2007,27(7):3032-3042.
[2]Jack B,Kris F,Robert J W.The edge effect and ecotonal species: birds communities across a natural edge in southeastern Australia[J].Ecology,2002,83(11):3048-3059.
[3]夏红霞,朱启红,宫渊波.生态交错带物种多样性研究综述[J].福建林业科技,2013,40(1):221-226.
[4]熊丹,欧晓昆,黄文君,等.基于土壤养分的农林生态系统生态交错带宽度测定[J].生态科学,2014,33(3):594-602.
[5]牟长城,韩士杰,罗菊春,等.长白山森林——沼泽生态交错带群落和环境梯度分析[J].应用生态学报,2001,12(1):1-7.
[6]贾宝全,慈龙骏,蔡体久,等.绿洲——荒漠交错带环境特征初步研究[J].应用生态学报,2002,13(9):1104-1108.
[7]夏哲超,潘志华,张璐阳,等.基于水分的北方农牧交错带植被生态系统退化机理研究——以武川县为例[J].资源科学,2010,32(2):317-322.
[8]郑翠玲,曹子龙,赵廷宁,等.浑善达克沙地南缘农牧交错带弃耕地植被的演替规律[J].中国水土保持科学,2005,3(1): 72-76.
[9]房用,王淑军,刘月良,等.现代黄河三角洲的植被群落演替阶段[J].东北林业大学学报,2008,36(9):89-93.
[10]王婷,欧晓昆,张志明,等.退耕还林工程影响下农林生态系统生态交错带的宽度测定[J].云南大学学报:自然科学版,2012,34(5):604-612.
[11]王志述.宁夏四儿滩湿地——干草原交错带边界判定及植被数量特征分析[D].北京:北京林业大学:2014:20-24.
[12]石培礼,刘兴良.游动分割窗技术在生态交错带定量判定中的应用:以四川巴郎山岷江冷杉林线为例[J].植物生态学报,2002,26(2):189-194.
[13]洪伟,吴承桢,林成来,等.福建龙栖山森林群落林窗边缘效应研究[J].林业科学,2000,36(2):33-38.
[14]李振基,刘初钿,杨志伟,等.武夷山自然保护区郁闭稳定甜槠林与人为干扰甜槠林物种多样性比较[J].植物生态学报,2000,24(1):64-68.
[15]何彤慧,程志,张玉峰,等.银川平原沟渠植物多样性特征及影响因素[J].湿地科学,2013,11(3):352-358.
Research on Boundary Definition and Plant Community Diversity of wetland-grassland Ecotone—A case study:Siertan Wetland in Ningxia
SU Xiangling1,WANG Zhenxi2,3
(1.Xinjiang Forestry Planning Institute,Urumqi 830049,China;2.College of Forestry and Horticulture,Xinjiang Agricultural University,Urumqi 830052,China;3.Key Laboratory of Forestry Ecology and Industry Technology in Arid Region,Education Department of Xinjiang,Urumqi 830052,China)
In order to study boundary definition and plant diversity of wetland-grassland ecotone in Yanchi country,Ningxia,vegetation investigation was carried out in July,2014.The moving split-window technique was applied to the quantitative determination of ecotone and diversity indexes(species diversity index,community dominant concentration index,community evenness index,species richness index)were used to analyze plant community diversity.The results showed:(1)When window width was 6 quadrats,itcan clearly delineate the ecotone boundary;The ecotone boundary of east,west,south,north line transect is 120m,120m,120m,140m respectively and distance from core area of wetland is 180m,220m,240m,200m respectively;(2)species diversity index,community evenness index,species richness index of the four transect lines all followed the order of ectone>arid region>wet region,while community dominant concentration index was in such an order as arid region>ectone>wet region;(3)The species diversity index,community dominant concentration index,community evenness index of north line transect were higher,but four diversity indexes of west line transect were lower.The results can provide theoretical support and scientific basis for the protection of valuable wetland resources,the maintenance of biological diversity and the stability of ecological system in arid and semi-arid regions.
ecotone,moving split-window technique,plant community diversity
S718.55;Q948
A
1002-6622(2017)01-0063-07
10.13466/j.cnki.lyzygl.2017.01.012
2016-11-09;
2016-11-23
新疆维吾尔自治区高校科研计划科学研究重点项目(XJEDU2013I16);中国博士后科学基金项目(2015M572668XB)
苏香玲(1976-),女,新疆塔城人,高工,学士,研究方向:森林经理。Email:645641681@qq.com
王振锡(1977-),男,河南郸城人,副教授,博士,研究方向:林业3S技术应用,森林生态与经营。
猜你喜欢
区域治理(2023年6期)2023-03-02 14:34:54
中国农学通报(2022年32期)2022-11-28 05:01:16
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
——以北长山岛为例
海洋开发与管理(2020年9期)2020-10-13 13:31:22
现代农业科技(2020年10期)2020-06-04 09:13:31
林业调查规划(2020年3期)2020-06-03 07:02:36
珠江水运(2017年17期)2017-10-10 08:16:03
中央民族大学学报(自然科学版)(2015年4期)2015-06-11 03:41:42
