
山东宁津附近海域大型底栖动物群落结构及季节变化*
2017-03-31 06:17王金宝李新正王洪法
海洋与湖沼 2017年6期
王金宝 李新正① 王洪法
(1. 中国科学院海洋研究所 青岛 266071; 2. 中国科学院大学 北京 100049; 3. 青岛海洋科学与技术国家实验室青岛 266237)
宁津街道位于山东半岛东端, 属山东省荣成市石岛区, 北、东、南三面环海。宁津海域自身独特的地形、地貌使得该处海流结构异常复杂, 受沿岸流和风场等因素影响, 该海域的海流存在着强烈的季节变化, 对大型底栖动物群落结构影响明显。宁津海域水深在近岸处较浅, 远岸处则较深, 呈明显的梯度增加现象, 等深线基本上与岸线平行, 整体水深比较浅,调查站位水深范围为6.75—38.38m。调查区域的桑沟湾为我国重要的海水养殖区, 海参、鲍鱼、海带、贝类等水产养殖业发达。外围海域为我国重要的黄海渔场, 许多甲壳动物、软体动物及棘皮动物等大型底栖生物具有重要的经济价值。底栖生物因其自身的生物特性, 一般栖息海底底内或底上, 迁移速度慢, 活动范围有限, 对逆境的逃避相对迟缓, 受环境影响更为深刻。因此, 大型底栖生物的种类组成、群落结构、次级生产力的变化能够更准确的反应海洋环境的长期、宏观变化(李新正, 2011)。大型底栖生物也是海洋生态系统的重要组成部分, 是能量流动和物质流动的重要环节。我国黄海近岸海域系统全面的大型底栖动物生态学研究始于二十世纪五十年代的全国海洋普查。多年来对南黄海大型底栖动物的研究较多(刘瑞玉等, 1963; 刘录三等, 2003; Liet al, 2005a, b;王金宝等, 2007; Zhanget al, 2012; 徐勇等, 2016; Xuet al, 2016a, b, 2017), 对北黄海大型底栖生物的主要研究近年来也有了较大进展(刘卫霞, 2009; 刘卫霞等,2009; 曲方圆等, 2009; 李新正等, 2016; 王金宝等,2016), 对山东半岛近岸海域大型底栖动物的研究主要有胶州湾、乳山湾、烟台近海等地(古丽娅诺娃等,1963; 李新正等, 2005; Liet al, 2007; 王金宝等, 2011;王洪法等, 2011; 王全超等, 2013)。目前, 关于黄海海域底栖生物尚缺少长期的、全海域范围的调查, 其群落变化规律及形成机制尚缺乏宏观分析(李新正,2011)。但是在山东半岛周边海域进行的大型底栖生物生态学调查研究较多, 有条件探讨该海域大型底栖生物群落变化机理, 预测未来变化趋势, 探究大型底栖生物生态服务功能, 提出资源保护建议, 为近海海域的开发利用提供基础数据。
石岛区域地质条件良好, 地势平坦, 相邻海域开阔, 水流湍急, 水体有较好的扩散条件, 多个大型海洋工程已经或准备选址于此地。针对附近海域进行了多次海洋生态调查(甘志彬等, 2012; 隋吉星等,2013)。相较于外海海域, 近岸及海湾等区域陆海相互作用强烈, 受人类活动干扰较大, 海洋捕捞、海水养殖及近岸海洋工程对大型底栖动物的群落结构产生较大影响。本次大型底栖动物调查对了解本海域的底栖动物群落结构和变化, 以及海洋工程建成运行前后对海洋环境的影响具重要意义。
1 材料与方法
1.1 调查海域和站位设置
根据不同的环境特点及受海岸工程活动影响的不同, 在宁津近海共布设15个站位(图1)。其地理范围在 36°42′59.05″—37°14′18.33″N、122°19′50.68″—122°50′31.07″E 之间。2009 年 11 月起共进行了 4 个调查航次, 分别在2009年11月(秋季)、2010年2月(冬季)、2010 年 5 月(春季)、2010 年 8 月(夏季)进行。根据工程要求和海区的自然状况, 秋季和春季对全部 15个站位进行了调查, 夏季和冬季则选取了重点站位SA4、SM4、SX10和SX11进行调查。
1.2 样品采集和处理方法
调查租用渔船进行。取样使用面积为0.1m2抓斗式采泥器(大洋-50型)定量采样, 各站成功取样2次合并为一个样品。所获泥样用底层为 0.5mm孔径的套筛冲洗获得大型底栖动物, 将获得的生物样品用95%的酒精固定后带回实验室。在实验室内对所获样品进行挑选、分大类、种类鉴定、个体计数和称重(精度为 0.001g的电子天平), 统计并计算种数、栖息丰度、生物量和次级生产力。样品采集和实验室内处理均按照《海洋调查规范》(GB/T12763.6-2007)进行。
1.3 数据处理
本研究的数据处理和分析主要通过 EXCEL、PRIMER 5.0、SURFER 12等软件完成。
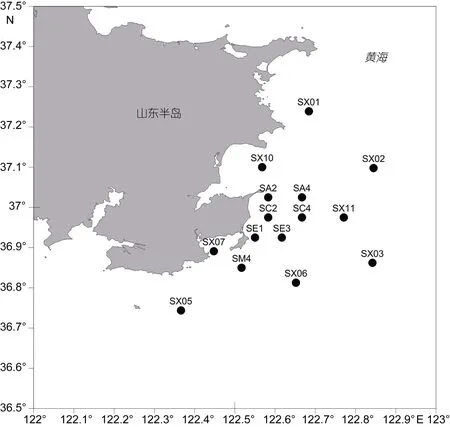
图1 宁津海域调查站位图Fig.1 The sampling stations in coastal waters off Ningjin
1.3.1 大型底栖动物的优势种类计算公式:

式中,N为采泥样品中所有种类的总个体数目,ni为第i种的个体数;fi为该种在各站位出现的频率; 当物种优势度Y>0.02时, 该种即为优势种(陈亚瞿等, 1995)。
1.3.2物种多样性指数运用物种多样性指数、物种均匀度指数和物种丰度指数对底栖动物数量动态特点进行分析。
Margalef物种丰富度指数(D)计算公式为:D= (S-1) / log2N
物种多样性指数采用 Shannon-Wiener指数(H′):
物种均匀度指数(J):J=H′ log2S
上述公式中,S为采泥样品中的种类总数,N为样品中个体总数,Pi为第i种底栖动物的个体数与样品中底栖动物总个体数的比值(ni/N)。
1.3.3群落结构分析
(1) 数据预处理
野外采集的样品经实验室定量鉴定分析后, 建立原始丰度矩阵, 为平衡优势种和稀有种在群落中的影响,数据经平方根转换后计算 Bray-Curtis相似性系数, 构建相似性矩阵。
(2) 群落划分
对转化后的相似性矩阵, 进行组间聚类分析(Group average cluster)和非度量多维标序(Non-metric Multi-Dimensional Scaling, MDS)分析群落结构。应用相似性百分比(SIMPER)计算同一群落中平均相似性,并分析主要贡献种。
(3) 群落稳定性分析
采用丰度/生物量比较曲线(ABC曲线)分析大型底栖动物群落受污染及扰动状况。在稳定的海洋环境中, 底栖生物群落结构近似平衡, 种内生物量的分布比丰度分布显优势, 生物量曲线明显在丰度曲线之上; 当底栖群落受到污染或扰动时, 大个体减少, 生物量曲线与丰度曲线相互靠近或者重叠在一起; 当群落受严重污染或剧烈扰动时, 对环境耐受强的小个体占优势, 生物量曲线明显在丰度曲线之下。
1.3.4次级生产力次级生产力的计算根据 Brey(1990)的经验公式:
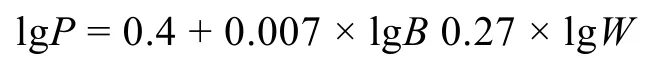
其中,P为每站大型底栖动物次级生产力[单位:g(AFDW)/(m2·a)],B为每站大型底栖动物年平均去灰干重生物量[单位: g(AFDW)/m2],W为每站大型底栖动物个体年平均去灰干重[单位: g(AFDW)/ind], 用精度为千分之一克的天平对各站物种称湿重, 根据经验公式, 湿重的9/50为去灰干重。由于W=B/A,A为每站大型底栖动物年平均栖息丰度(单位: ind/m2), 转换后:P=A0.27×B0.737/ 100.4(Liet al, 2005b)。
P/B值是生产力与去灰干重生物量的比值, 被认为是种群最大可生产量的指示值。该值指示了生物量的年度周转次数。
2 结果
2.1 种类组成及优势种
本调查研究在宁津附近海域四个季度月完成了38个站次调查, 经鉴定大型底栖动物隶属于11个门类, 共有159种。其中环节动物门的多毛类73种, 软体动物门25种, 节肢动物门39种, 棘皮动物门9种,其他类群共13种。多毛类种数最多。
由于季节变化和调查站位设置存在差异, 所以各季节的物种数有明显不同。秋季共鉴定出大型底栖动物 67种, 其中多毛类环节动物 40种, 占 59.70%;甲壳动物 14种, 占 20.90%; 软体动物 8种, 占11.94%; 棘皮动物 2种, 占 2.99%; 其他类群动物 3种, 占4.48%。冬季鉴定37种, 其中多毛类环节动物19种, 占51.35%; 甲壳动物7种, 占18.92%; 软体动物3种, 占8.11%; 棘皮动物3种, 占8.11%; 鱼类1种, 占2.70%; 其他类群动物4种, 占10.81%。春季鉴定 81种, 其中多毛类环节动物 35种, 占 43.21%;甲壳动物 25种, 占 30.86%; 软体动物 10种, 占12.35%; 棘皮动物 6种, 占 7.41%; 其他类群动物 5种, 占6.17%。夏季调查设大型底栖生物调查站4个,共采到大型底栖动物 52种, 其中多毛类环节动物种数最多为21种, 占40.38%; 其他依次为甲壳动物15种, 占28.85%; 软体动物9种, 占17.31%; 棘皮动物4种, 占7.69%; 其他类群动物3种, 占5.77%。秋季和春季调查站位相同, 冬季和夏季调查站位相同。春季种数大于秋季, 夏季种数大于冬季。
各航次间优势种组成出现一定差别, 但均以多毛类为主。拟特须虫(Paralacydonia paradoxa)在四个航次均为优势种。丝异蚓虫(Heteromastus filiforms)、索沙蚕(Lumbrinerissp.)、轮双眼钩虾(Ampelisca cyclops)出现在三个季度月。甲壳动物的钩虾类有两种作为优势种出现, 而棘皮动物仅日本倍棘蛇尾在春季作为优势种出现(表1)。
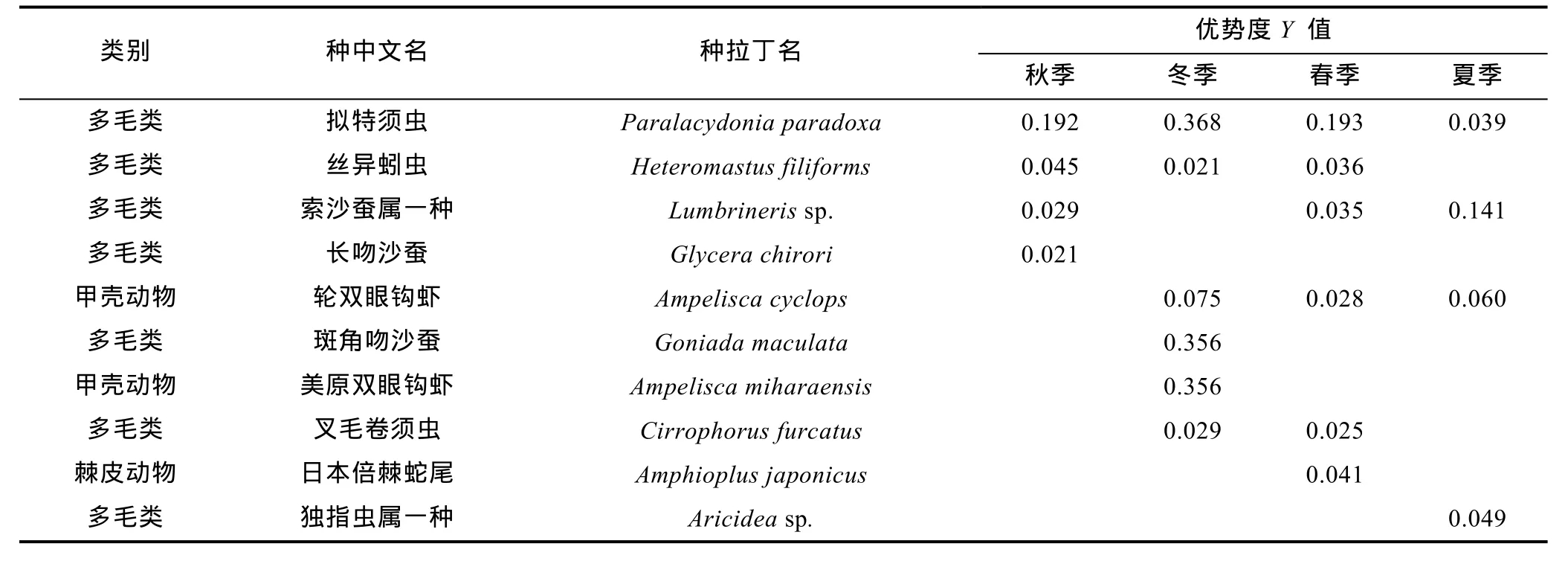
表1 各季度月优势种及优势度值Tab.1 The dominant species and their values of dominance
2.2 丰度和生物量
2.2.1丰度四个季度月航次调查大型底栖生物丰度平均值为 233.90ind./m2。秋季航次平均丰度为146.00ind./m2, 其中多毛类平均丰度为121.00ind./m2,占82.88%; 其次为甲壳动物 14.33ind./m2, 占 9.82%;其他类群最少仅为 1ind./m2, 占 0.68%, 丰度最高的站是SX11站, 达500ind./m2; 最低的为SA2站, 仅采集到几个多毛类碎片。冬季航次平均丰度为316.25ind./m2, 其中多毛类动物平均丰度最高为215.00ind./m2, 占67.98%; 其次为甲壳动物70.00ind./m2, 占 22.13%; 鱼类最少仅为 5ind./m2, 占0.40%。从站位来看, 丰度最高的站是 SX11 站, 达570ind./m2, 最低的为SM4站, 为60ind./m2。春季平均丰度为178.33ind./m2, 其中多毛类动物的平均丰度占了绝对优势, 为116.67ind./m2, 占65.42%; 其次为甲壳动物35.33ind./m2, 占19.81%; 棘皮动物为17.00ind./m2, 占 9.53%; 软体动物为 6.00ind./m2, 占3.36%; 其他类群动物的平均丰度最少仅为3.33ind./m2, 占 1.87%。分站位看, 丰度最高的站是SX10站, 达 395ind./m2; 较低的站位为 SE1站仅5ind./m2, 而 SE3站仅采到水螅类的部分群体。夏季航次平均丰度为295.00ind./m2, 其中多毛类动物的平均丰度占绝对优势, 为158.75ind./m2, 占53.81%; 其次为甲壳动物 57.50ind./m2, 占 19.49%; 软体动物46.25ind./m2, 占 15.68%; 棘皮动物 21.25ind./m2, 占7.20%; 其他类群动物的平均丰度最少 11.25ind./m2,占3.81%。丰度最高的是SX10站, 达570ind./m2; 最低的是 SX11站, 为 145ind./m2。不同季节平均丰度出现一定变化, 春季>秋季; 冬季和夏季相差不大,冬季>夏季。各季节均以多毛类占优势。
从各季度月丰度空间分布(图 2)可以发现, 调查海域丰度分布的基本趋势为外海大于沿岸, 北部海域大于西南海域, 桑沟湾区域丰度相对较高。分季节看, 秋季在宁津近岸海域明显丰度较低, 外围海域较大; 冬季, 各站位差别较大, 近岸的SM4站丰度最低;春季也表现为近岸低, 外围高, 但桑沟湾内站位丰度相对较高; 夏季调查站位较少, 只有桑沟湾内 SX10站较高, 其他各站均较低。
2.2.2生物量四个季度月航次调查大型底栖生物平均生物量为 9.144g/m2。秋季航次不同站位生物量差距较大, 平均生物量为 15.302g/m2, 其中多毛类动物占了绝对优势, 为13.633g/m2, 占89.09%, 最少的类群为棘皮动物, 平均生物量为 0.215g/m2, 仅占1.41%。生物量最高的站为SX01站, 达160.025g/m2,最低的为 SA2站, 仅为 0.010g/m2。冬季航次平均生物量为 4.623g/m2, 其中棘皮动物占优势, 为1.695g/m2, 占 36.67%; 软体动物最少, 平均生物量为0.168g/m2, 占3.62%。分站位看, 生物量最高的站为 SM4站, 7.570g/m2, 最低的为 SA2站, 为1.470g/m2。春季航次平均生物量为 3.397g/m2, 其平均生物量和所占比例依次为多毛类动物 1.688g/m2,占 49.70%; 甲壳动物 0.652g/m2, 占 19.20%; 软体动物 0.518g/m2, 占 15.25%; 棘皮动物 0.418g/m2, 占12.32%, 生物量最高的站为SX10站, 达11.995g/m2,最低的为 SE1站, 仅为 0.125g/m2。夏季航次平均生物量为 13.254g/m2。棘皮动物最高为 6.706g/m2, 占50.60%; 多毛类次之为 5.196g/m2, 占 39.21%; 其次为甲壳类为 0.698g/m2, 占 5.26%; 再次为软体动物4.178g/m2, 占4.18%; 其他类群0.100g/m2, 占0.76%。生物量最高的站为 SX11站, 达 24.685g/m2, 最低的是 SA2站, 为 8.155g/m2。不同季节间平均生物量出现较大差异, 秋季最高, 春季最低。表现为秋季>春季,夏季>冬季。
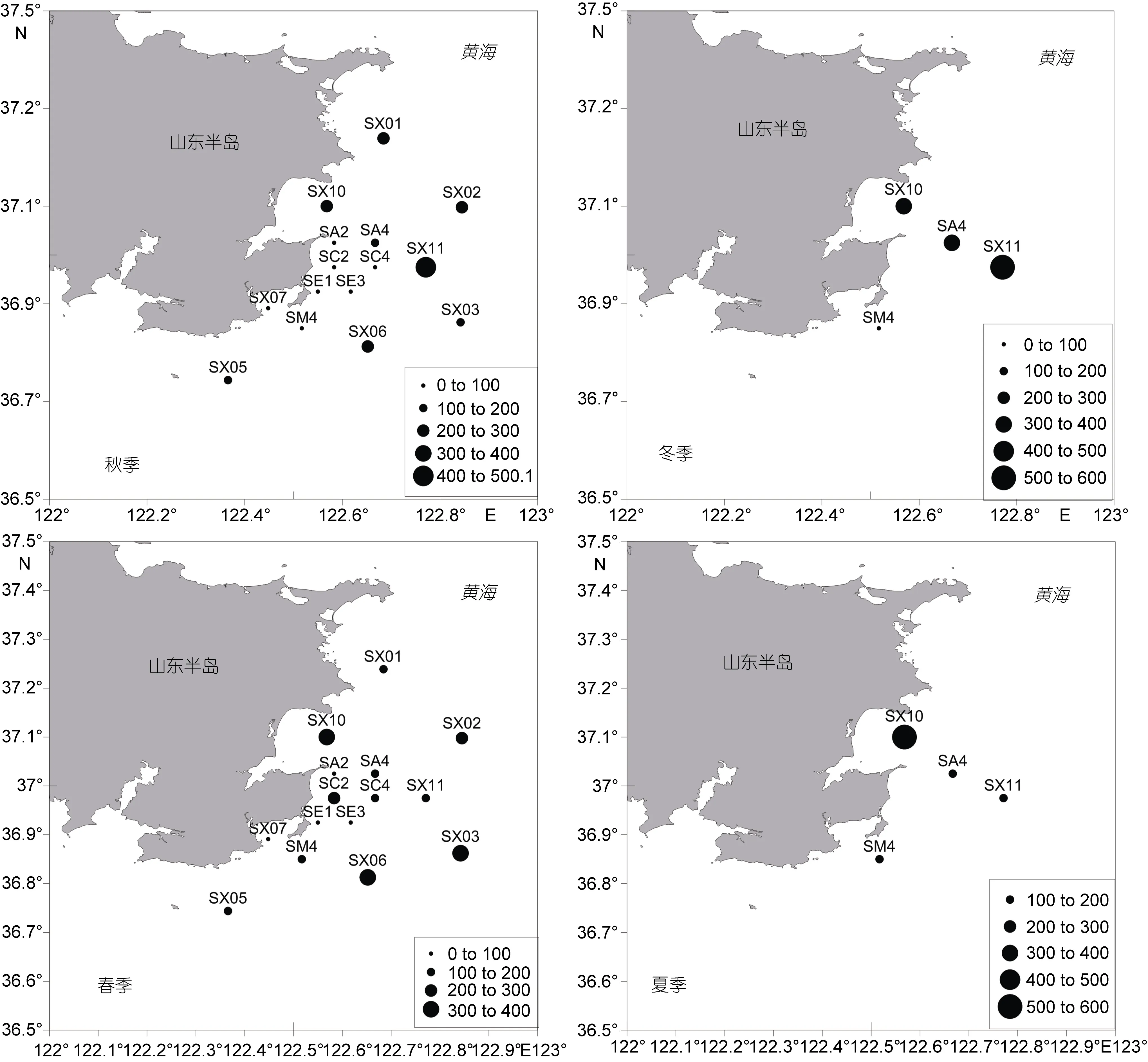
图2 宁津近岸海域大型底栖动物丰度不同季节的空间分布Fig.2 Spatial distribution of macrobenthic abundance in different seasons in coastal waters off Ningjin
图 3为各季度月生物量空间分布图。可以发现,生物量、丰度空间分布有相似性, 但各季节又表现出一定差异。调查海域生物量分布的基本趋势为外海大于沿岸, 各站位不同季节生物量差别较大, 桑沟湾区内SX10站生物量较平均。分季节看, 秋季在宁津近岸海域生物量较低, 外围和北部海域相对较大; 冬季,近岸的 SM4和远岸的 SX11站生物量相对丰度来说较高; 春季也表现为近岸低, 外围高, 桑沟湾内生物量高; 夏季调查站位较少, 外围海域站位较高。
2.3 次级生产力和P/B值
2.3.1次级生产力根据次级生产力公式计算各站位的次级生产力结果如表2所示。本调查海区次级生产力较低, 平均值仅为 1.903g(AFDW)/(m2·a), 其中, 夏季 3.038g(AFDW)/(m2·a)> 秋季 2.746g(AFDW)/(m2·a)>冬季1.472g(AFDW)/(m2·a)>春季1.073g(AFDW)/(m2·a)。在次级生产力的空间分布方面, 平均值最高的是 SX01站, 为 10.428g(AFDW)/(m2·a), 位于调查区的北部; 最低的是 SE1站, 为 0.031g(AFDW)/(m2·a), 靠近宁津沿岸。次级生产力最高值出现在秋季的 SX01站, 为 20.201g(AFDW)/(m2·a), 该站生产力高是因为采集到个体较大的多毛类澳洲鳞沙蚕(Aphrodita australis); 除去未采集到标本的站位外,最低值是秋季的SE1站, 为0.025g(AFDW)/(m2·a)。

图3 宁津近岸海域不同季节大型底栖动物生物量的空间分布Fig.3 Spatial distribution of macrobenthic biomass in different seasons in coastal waters off Ningjin
2.3.2P/B值各站位不同季节的P/B值如表2所示。调查海域P/B值的平均值为1.801。分季节看, 冬季(2.203)>春季(1.833)>秋季(1.806)>夏季(1.461), 夏季P/B值相对较小, 冬季最大。各站位的年度P/B平均值中最高的是SE1站, 为2.571; 最低的是SX01站,为 1.467。P/B值最高的站位是夏季的 SX11站, 为4.581, 最低值的站位是秋季的SX01, 为0.701。
2.4 物种多样性指数
不同季节各站位物种多样性指数数值如表3所示。
丰富度指数(D)的年平均值为2.436, 各航次间变化较大。秋季各站位的丰富度指数相对较小, 均值为1.783(SA2未计算), 其他最小为SE1站(0.311), 最大为 SX01站(2.979); 冬季仅采集了四个站位, 均值为2.346, 最小 SM4站(0.977), 最大为 SX10站(3.104);春季均值为2.619, SE1和SE3未计算, 其他站位最小为SC2站(1.292), 最大为SX02站(3.727); 夏季均值为2.996, 仅采集了四个站位, 最小为SM4站(2.561),最大为SX10站(3.940)。不考虑采样站位数量的差别,夏季>春季>冬季>秋季。
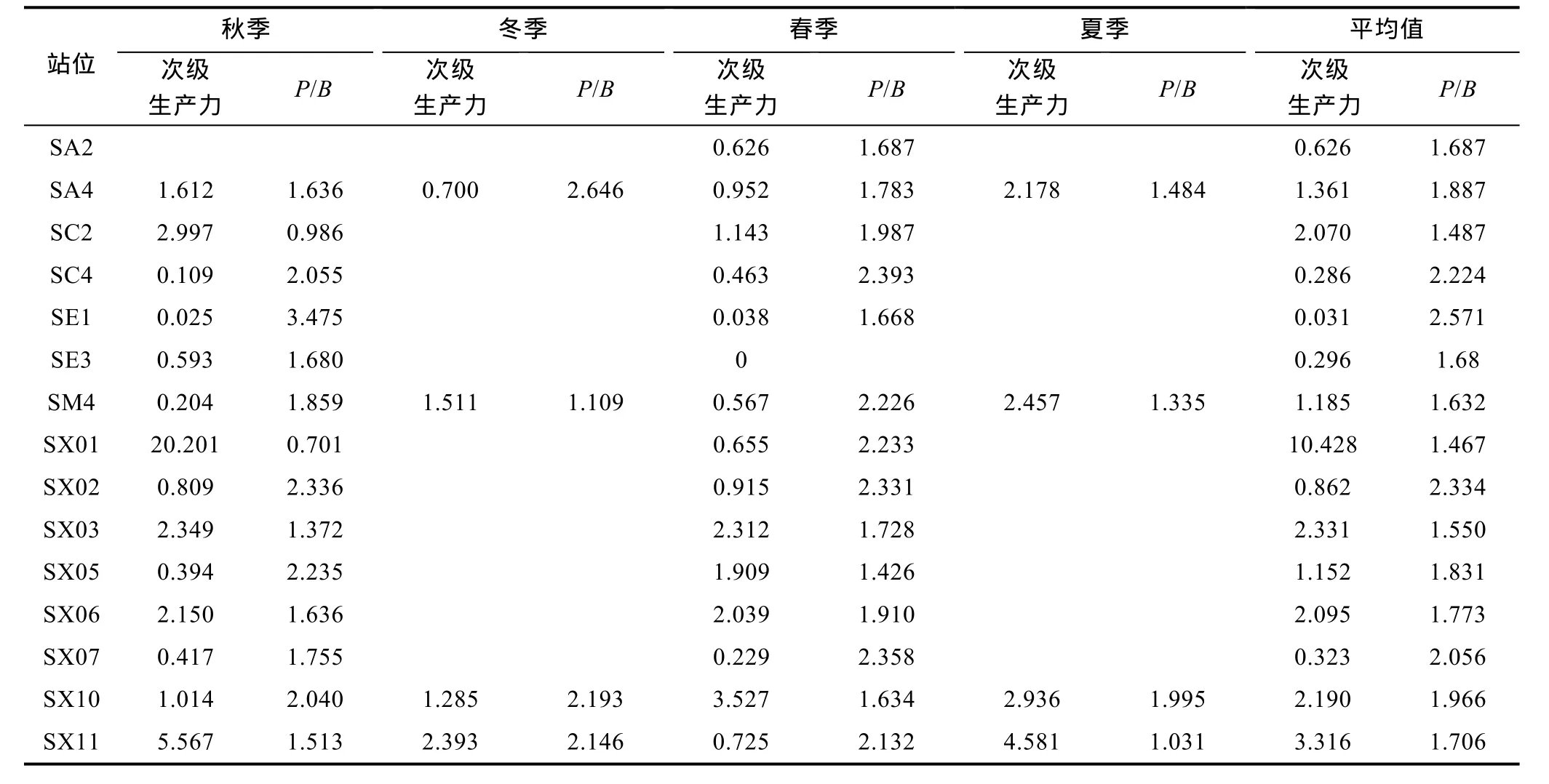
表2 不同季节各站位的次级生产力g(AFDW)/(m2·a)和P/B值Tab.2 The secondary productions of macrobenthos and the P/B values in different seasons in each station
均匀度指数(J)的年平均值为 0.800, 各航次间变化不大。秋季各站位的均匀度指数相对较小, 均值为0.811, SA2未计算, 其他最小为SX06站(0.641), 最大为 SC4站(0.961); 冬季均值为 0.706, 最小 SX11站(0.608), 最大为 SA4站(0.814); 春季均值为 0.832,SE1站和 SE3站未计算, 其他站位最小为 SX11站(0.724), 最大为SA2站(0.950); 夏季均值为0.850, 仅采集了四个站位, 最小为 SX10站(0.738), 最大为SX11站(0.892)。不考虑采样站位数量的差别, 夏季>春季>秋季>冬季。
物种多样性指数(H')的年平均值为 2.924, 不同季节出现一定变化。秋季各站位的多样性指数相对较小, 均值为 2.469, SA2未计算, 其他最小为 SE1站(0.722), 最大为 SX01站(3.582); 冬季仅采集了四个站位, 均值为 2.617, 最小 SM4站(1.585), 最大为SA4站(3.257); 春季均值为3.166, SE1站和SE3站未计算, 其他站位最小为 SC2站(2.254), 最大为 SX02站(3.797); 夏季仅采集了四个站位, 均值为3.444, 多样性较高, 各站位差别较小, 最小为 SM4站(3.337),最大为SX11站(3.486)。不考虑采样站位数量的差别,夏季>春季>冬季>秋季。
2.5 群落结构
图4—图7为秋季和春季大型底栖动物群落结构聚类分析和MDS分析结果。因冬季和夏季采样站位较少, 仅四个站位, 所以未做群落结构分析。秋、春两个航次群落在基本稳定的基础上出现一定的季节变化。根据聚类结果, 秋季和春季均可分为5个群落。秋季航次大型底栖动物在 30%的 Bray-Curtis相似性水平上, 分为5个群落。群落Ⅰ包括8个站位(SA4、SE3、SX01、SX02、SX03、SX05、SX06、SX11), 是外海群落。SIMPER分析表明群落Ⅰ的平均相似性为37.39%, 主要贡献种为拟特须虫(39.28%)、丝异蚓虫(12.27%)、索沙蚕(11.20%), 三个种累计贡献率为62.75%; 群落Ⅱ包括3个站位(SC4、SM4、SX07), 是石岛湾和中间过渡群落。SIMPER分析表明群落Ⅱ的平均相似性为 36.95%, 主要贡献种为拟特须虫(53.17%)、长吻沙蚕(46.83%); SX10站是桑沟湾群落;SE1和 SC2是近岸群落, 但两者组成差别较大, SE1主要是甲壳动物钩虾类, SC2站甲壳动物和棘皮动物较多, 可能与两站位底质不同有关。
春季航次共调查15站位。SE1仅采集到一种群体生物标本, SE3未采集到标本。SE1站的群体生物与其他各站均不同, 其余各站的相似性较高。春季各
调查站位大型底栖动物在30%的Bray-Curtis相似性水平上, 可分为 5个群落。群落Ⅰ包括 7个站位(SA4、SM4、SX02、SX03、SX05、SX06、SX11), 是外海群落。SIMPER分析表明群落Ⅰ的平均相似性为 37.39%, 与秋季群落Ⅰ相似性相同。主要贡献种为拟特须虫(37.35%)、丝异蚓虫(8.43%)和日本倍棘蛇尾(8.72%), 三个种累计贡献率为54.50%。群落Ⅱ包括SC4和SX07站, 属于石岛湾和过渡区域群落。SIMPER分析表明群落Ⅱ的平均相似性为41.71%。主要贡献种为丝异蚓虫(26.24%)、叉矛卷须虫(21.56%)和不倒翁虫(Sternaspis scutata)(21.56%),三个种累计贡献率为69.51%。群落Ⅲ包括SX01和SX10, 属于桑沟湾和北部群落, SIMPER分析表明群落Ⅲ的平均相似性为32.40%。主要贡献种为索沙蚕(30.60%)、寡鳃齿吻沙蚕(Nephtys oligobranchia)(23.13%)和长吻沙蚕(11.57%), 三个种累计贡献率为65.30%。群落Ⅳ包括SA2和SC2, 属于宁津近岸群落。SIMPER分析表明群落Ⅳ的平均相似性为38.38%。主要贡献种为长吻沙蚕(31.35%)、拟特须虫(25.43%)和日本倍棘蛇尾(25.43%), 三个种累计贡献率为82.02%。
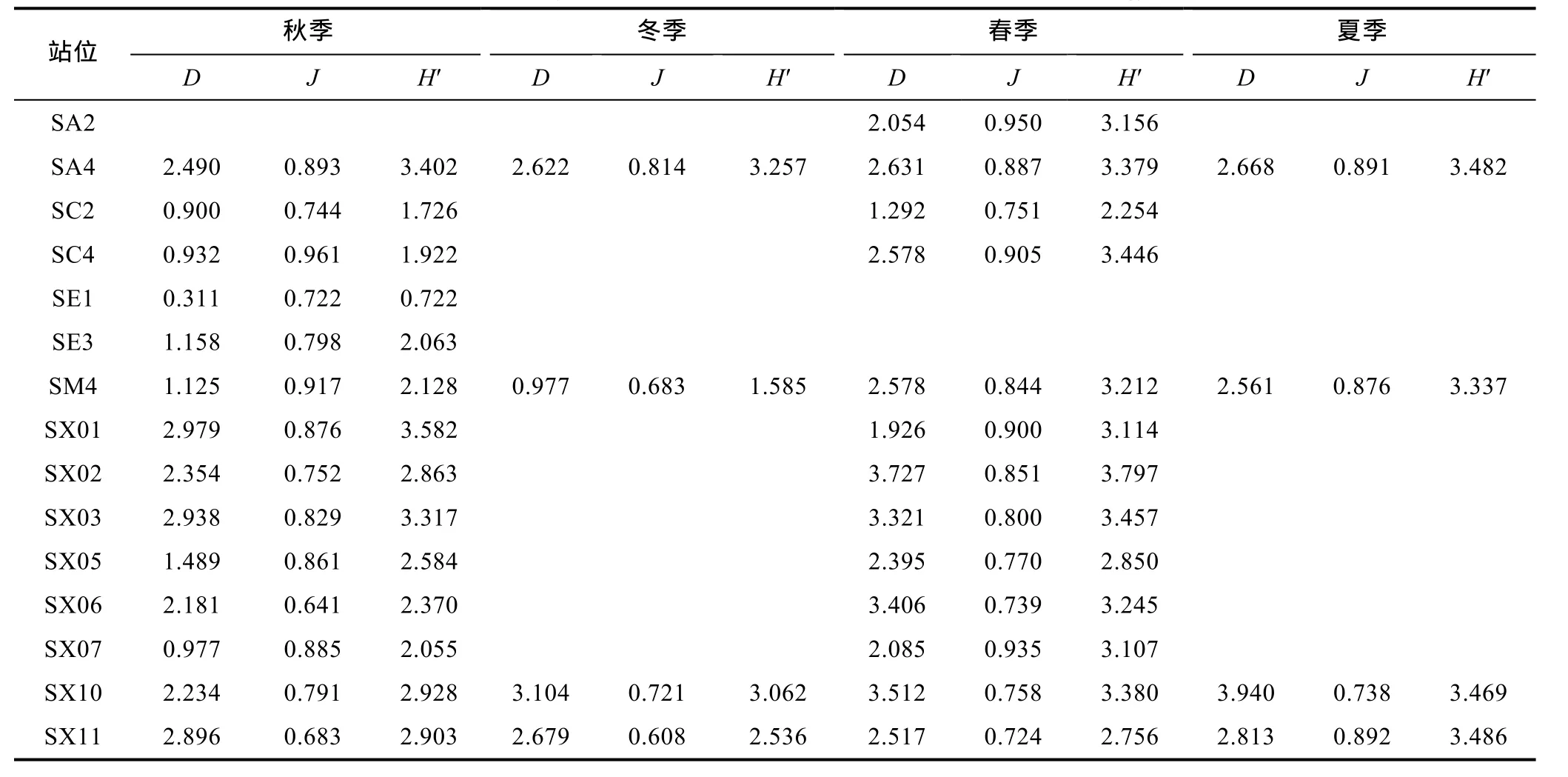
表3 宁津近海大型底栖动物多样性指数(丰富度指数D, 均匀度指数J, 物种多样性指数H′)Tab.3 Diversity indices of macrobenthos in coastal waters off Ningjin
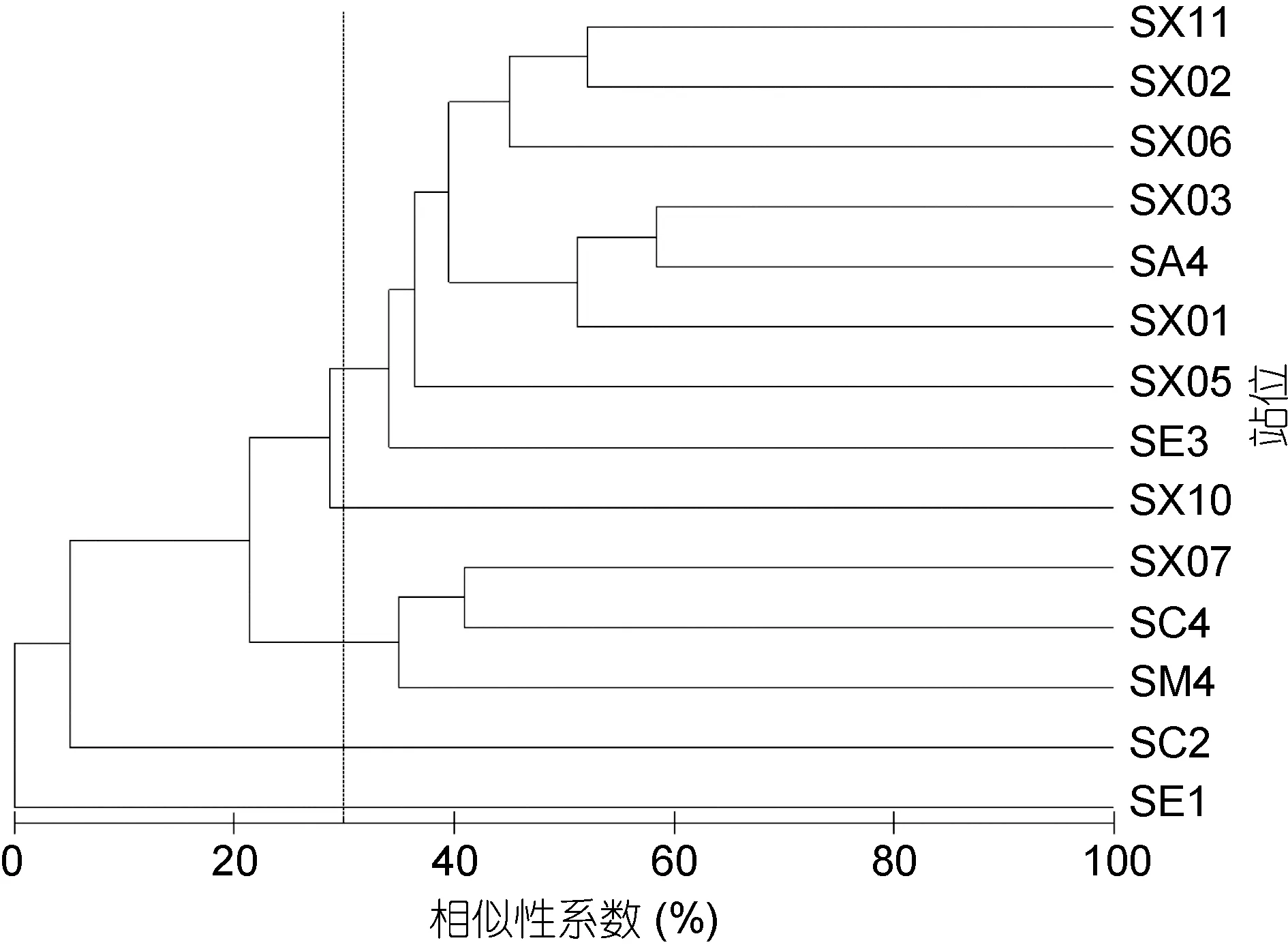
图4 秋季大型底栖动物群落结构聚类分析Fig.4 The graph of cluster analysis in the autumn
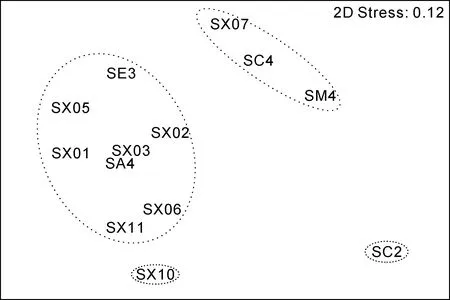
图5 秋季大型底栖动物群落结构MDS分析Fig.5 The graph of MDS analysis in the autumn
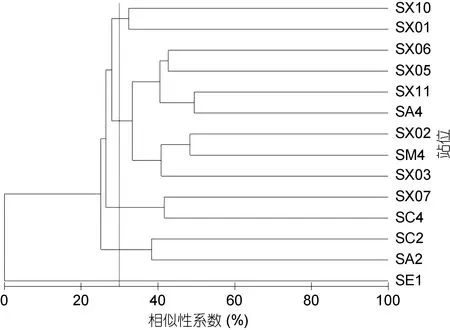
图6 春季大型底栖动物群落结构聚类分析Fig.6 The graph of cluster analysis in the spring
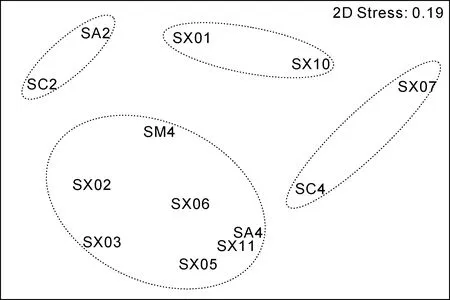
图7 春季大型底栖动物群落结构MDS分析结果Fig.7 The graph of MDS analysis in the spring
2.6 ABC曲线
图8和图9分别为秋季和春季宁津海域大型底栖动物群落的ABC曲线。从ABC曲线分析各季节底栖动物的群落受扰动情况, 两个航次生物量曲线均在丰度曲线之上, 说明海域受扰动情况较轻。2009年11月至2010年5月航次的W值分别为0.409和0.536,均为正值, 也说明了环境受外界影响小, 群落结构较稳定。
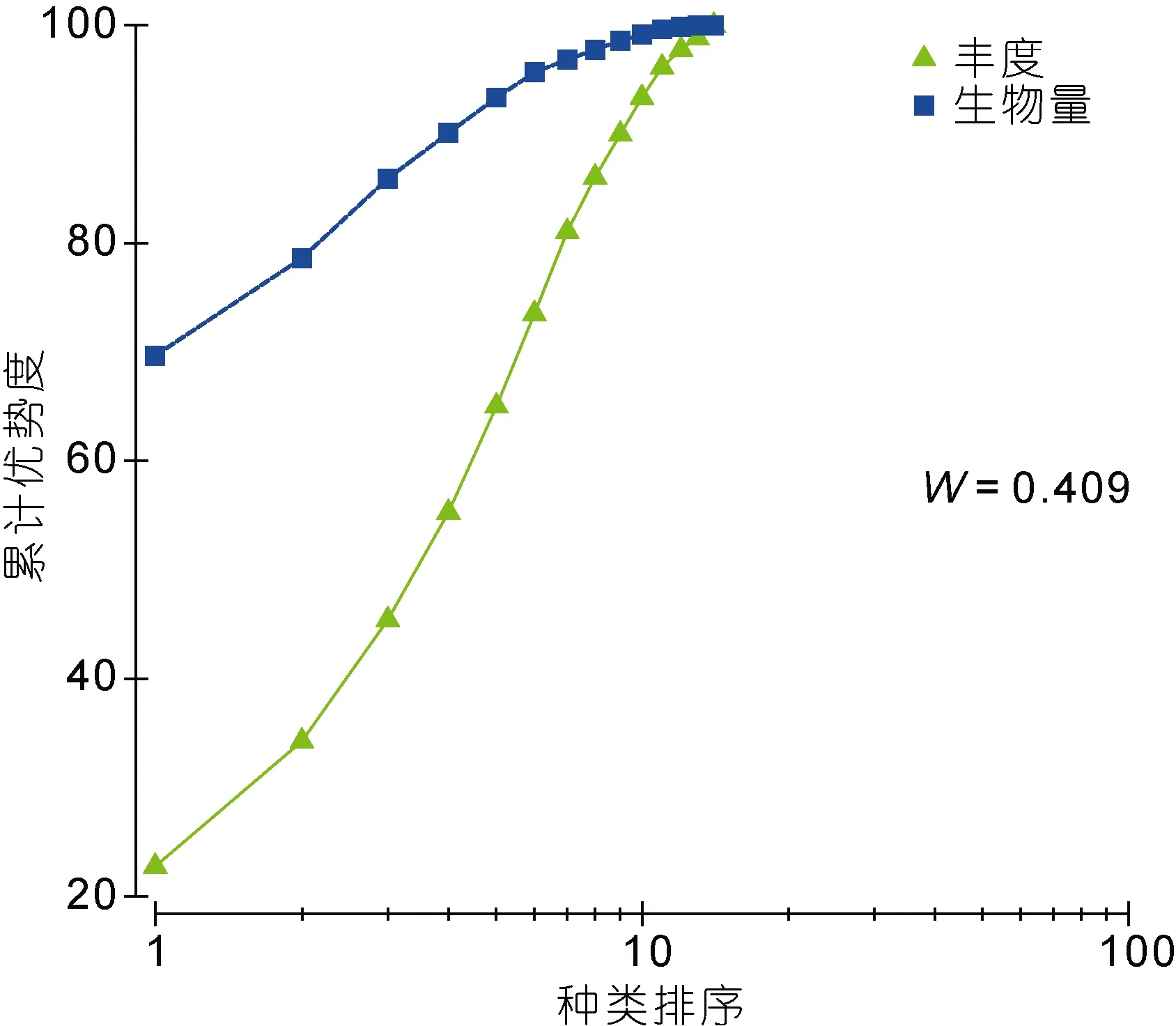
图8 秋季大型底栖动物ABC曲线Fig.8 ABC plot of macrobenthic community in the autumn
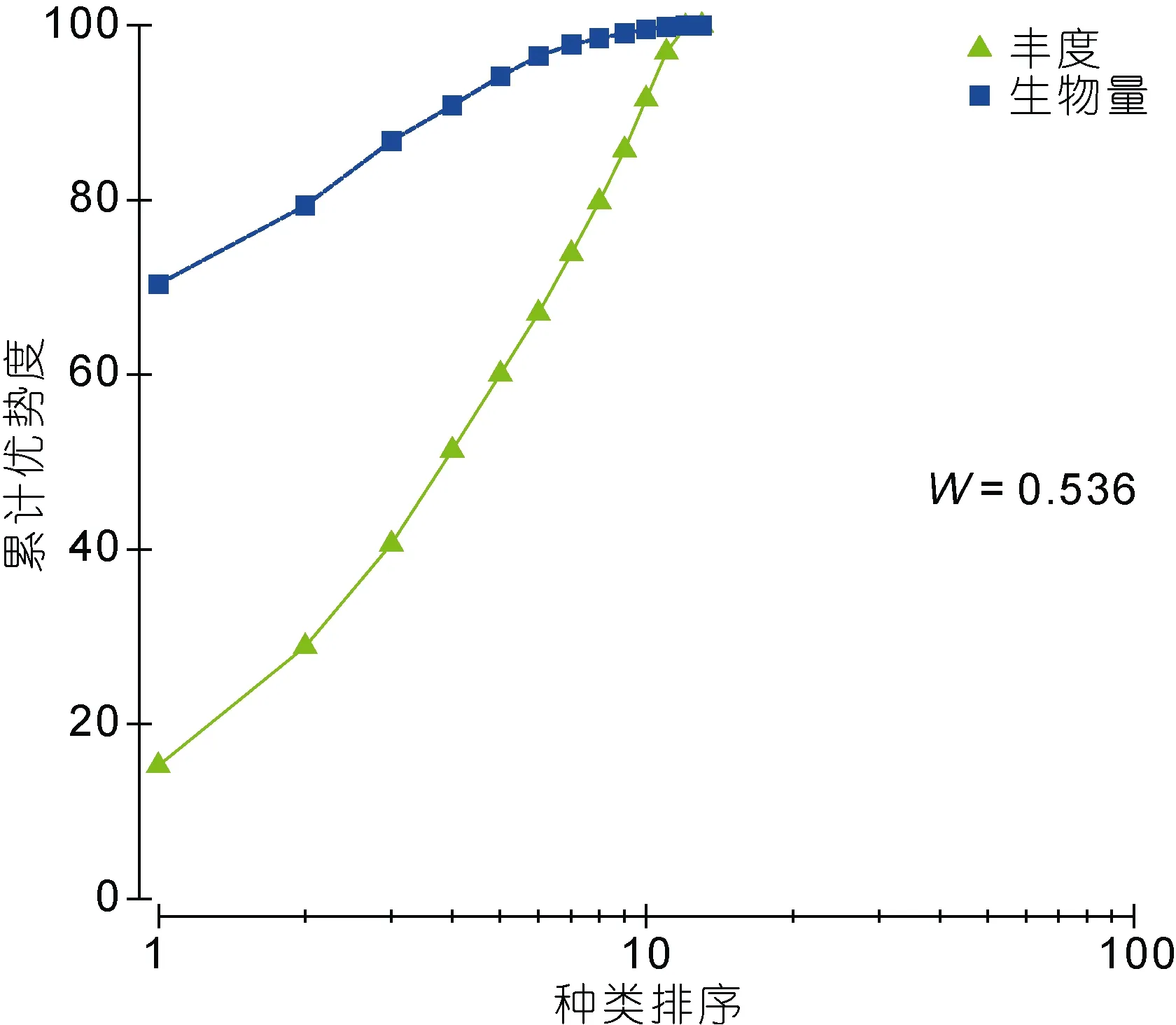
图9 春季大型底栖动物ABC曲线Fig.9 ABC plot of macrobenthic community in the spring
3 讨论
3.1 种类组成
3.1.1调查海区大型底栖动物年度变化物种数量和种类组成是群落结构的基础。通过物种数量的变化可以了解群落结构的变化, 物种多样性越高的区域, 群落结构越稳定。通过不同年度物种数的变化,可以对该区域群落变化趋势有一定的了解。宁津海域在2007年度及2008—2009年度分别进行了四个季度月调查。2007年该区域共采集到大型底栖动物 243种, 其中多毛类动物 96种; 软体动物 43种; 甲壳动物77种; 棘皮动物13种; 其他类群动物14种(甘志彬等, 2012); 2008—2009年度共鉴定大型底栖生物164种, 其中多毛类90种, 甲壳动物35种, 软体动物17种, 棘皮动物 8种,其他类群 14种(隋吉星等,2013)。本调查共发现159种底栖生物, 各调查均以多毛类种数最多, 调查区域大型底栖动物种数有减少的趋势。
调查海域的优势种在保持稳定性的基础上也出现一定变化。2007年主要优势种有拟特须虫、日本倍棘蛇尾、不倒翁虫和叉毛卷须虫等(甘志彬等, 2012);2008—2009年主要优势种有拟特须虫、叉毛卷须虫、索沙蚕、日本倍棘蛇尾等(隋吉星等, 2013); 本次调查中拟特须虫、叉毛卷须虫、日本倍棘蛇尾等依然是主要的优势种, 但不倒翁虫则没有出现在优势种中, 且日本倍棘蛇尾仅在春季作为优势种出现, 优势度有降低的趋势。
相对于种数, 物种多样性指数考虑了样品数量大小, 更能全面反映群落的结构特征。2007年该海域的物种多样性指数 3.075, 均匀度指数秋季(0.86)>冬季(0.85)>夏季(0.82)>春季(0.81), 各季节相差不大(甘志彬等, 2012)。本调查物种多样性指数的年平均值为2.924, 明显小于2007年的调查。季节变化则是夏季>春季>冬季>秋季, 也呈现出一定的不同。
通过以上比较可以发现, 本次调查物种数量、物种多样性指数均有所减小, 优势种也出现了一定变化, 说明该海域大型底栖动物群落结构出现了一定变化。
3.1.2与邻近海区的比较调查区以北为烟台周边海域, 与调查区域相邻。王全超等(2013)在2010—2011年对烟台近海的调查共获大型底栖动物176种,与本海域的调查相差不大。但优势种的组成则有较大的差别。烟台海域主要优势种为短叶索沙蚕(Lumbrineris latreilli)和不倒翁虫, 在四个航次中均为优势种, 且短叶索沙蚕在群落中的优势地位极显著。但本海域不倒翁虫未作为优势种出现, 索沙蚕属的种类优势度也较低。可能是因为烟台调查海域位于半岛北部, 水流相对平缓, 调查站位靠近沿岸, 群落结构出现不同。
2006—2007年, Li(2009)等对乳山湾近岸海域的调查研究共鉴定出大型底栖动物236种, 与宁津海域2007年的调查相差不大。乳山湾 4个季度航次共有优势种为寡鳃齿吻沙蚕(Nephtys oligobranchia)和不倒翁虫, 其优势种的组成与宁津海域明显不同, 寡鳃齿吻沙蚕未出现在宁津海域的优势种中, 且不倒翁虫仅在 2007年出现过, 宁津海域的优势种由个体更小的多毛类拟特须虫和丝异蚓虫等组成。
3.2 生物量和丰度的年度和季节变化
3.2.1本海域年度和季节变化2007年调查海域年平均生物量和丰度分别为为9.5g/m2和219.6ind./m2(甘志彬等, 2012), 本次调查年平均生物量和丰度分别为 9.144g/m2和 233.90ind./m2。两者比较, 本调查生物量稍低, 但丰度增大, 变化不是十分明显。本调查秋季生物量和栖息丰度均值分别为15.302g/m2和146.00ind./m2, 与2007年秋季调查相比,平均生物量增高, 其原因是采到了个体较大的澳洲鳞沙蚕所致; 平均丰度降低, 其原因与调查站数目和位置的选取有关, 也与采集到的软体动物减少有关。春季平均生物量和栖息丰度为3.397g/m2和178.33ind./m2, 与 2007年春季调查相比存在较大差异, 平均生物量降低, 其原因主要是 2007年在 SE4站采到了个体较大的心形海胆(Echinocardiumcordatum)所致。各季度之间平均生物量和平均丰度存在较大差异, 个体较大的底栖动物对生物量影响明显。冬季和夏季由于航次调查站位较少, 变化较大,未予对比。
3.2.2与邻近海域比较烟台近海大型底栖动物平均生物量为 16.72g/m2, 明显大于宁津近海的平均生物量。烟台海域生物量以软体动物贡献最高, 而宁津海域主要以多毛类为主, 群落结构的不同影响了生物量的数值。分季节看, 烟台海域生物量季节间变化明显, 夏、秋两季的平均生物量(分别为 25.09g/m2和 25.76g/m2) 高于春、冬两季(分别为 6.04g/m2和9.97g/m2)(王全超等, 2013)。而宁津海域平均生物量也表现为夏、秋季明显大于冬、春季, 两者的季节变化规律相同。烟台海域大型底栖动物平均丰度为1022ind./m2, 宁津海域平均丰度仅为 233.90ind./m2,远小于烟台海域, 两区域丰度主要贡献类群均为多毛类动物。两区域物种数相近, 但宁津海域生物量和丰度明显小于烟台海域, 说明两者群落结构差异较大。宁津近岸部分站位的细沙底质不利于底栖生物的生存。
乳山湾的平均生物量和丰度分别为100.2g/m2和248.6ind./m2。丰度和宁津海域相差不大, 但生物量相差达到了10倍。出现这种状况主要是因为乳山湾海域底质以软泥为主, 水流较小, 生态环境良好, 生物资源远较宁津海域丰富, 个体较大的软体动物等较多。而宁津主要以小个体的多毛类和钩虾类占优。
3.3 次级生产力
宁津海域较低的生物量导致了次级生产力也较低, 平均值仅为 1.903g(AFDW)/(m2·a), 远低于北黄海獐子岛海域的次级生产力 15.335g(AFDW)/(m2·a)(王金宝等, 2016), 也低于南黄海的 4.98g(AFDW)/(m2·a) (Liet al, 2005a)。说明该海域底栖生物相对贫瘠, 这可能与流经该处较强的沿岸流有关, 较强的海流导致海底有机物质减少, 不利于底栖生物的生存发展。
3.4 群落结构变化分析
大型底栖动物的群落结构与底质类型、深度等环境因子密切相关。SA2为泥沙碎壳底质、SE1为细沙底质、SE3为硬质泥等, 这几个站位靠近岸边, 未采集到标本或采集到极少, 与其他软泥底质站位有明显不同, 群落结构差别较大。另外, 受到海底地形、水流、人为扰动、深度等因素的影响, 宁津海域的大型底栖动物丰度和生物量呈斑块状分布, 但总体表现为由近岸向外海, 丰度和生物有增加趋势。除近岸的个别站位外, 大部分站位的构成差异不明显。聚类分析和MDS分析也表明各站位的群落结构差异不明显。由于该海域靠近岸边, 水深较小, 不同季节的温差大, 对群落的影响较大, 不同季节的平均生物量和丰度差异较大, 说明了该海域底栖动物数量受季节变化影响较大。底栖动物行动力较弱, 从群落结构上看, 又保持了一定的稳定性。宁津周边有数个大型近岸海洋工程正在建设或筹备建设, 通过工程建设前、建设中和建设完成后运行阶段的底栖动物调查, 了解其群落结构变化, 可以评估海洋工程对生态系统功能的影响, 为以后工程建设的环境影响评价提供借鉴。
4 结论
相对于周边海域, 调查海域大型底栖动物种类较少且有逐渐减少的趋势, 优势种保持基本稳定, 并出现一定变化, 物种多样性指数较低且较往年出现减小的趋势。宁津邻近海域年平均生物量和丰度均较低且存在季节性差异。次级生产力平均值仅为1.903g(AFDW)/(m2·a)。秋、春航次群落分布格局均包含5个左右的群落, 主要有近岸群落、中间过渡群落、远岸群落以及桑沟湾群落等群落, 远岸群落包含的站位最多, 各群落间界线不明显, 对各群落贡献率较大的种主要是环节动物多毛类。大型底栖动物群落结构受污染或扰动情况不明显。
致谢李宝泉、蔡文倩、董栋、董超、马林、王晓晨、寇琦、隋吉星、彭松耀、徐勇、许鹏、张鹏弛等参加或部分参加了本研究的野外考察、实验室样品鉴定和处理、数据分析工作, 并在论文写作过程中给予良好建议和大力帮助, 中国科学院海洋生物标本馆为本研究的样品处理给予了大力支持和协助, 谨致谢忱。
王全超, 李宝泉, 2013. 烟台近海大型底栖动物群落特征. 海洋与湖沼, 44(6): 1667—1680
王金宝, 李新正, 王洪法, 2016. 北黄海獐子岛附近海域大型底栖动物数量分布和季节变化. 海洋科学集刊, 51(1):258—268
王金宝, 李新正, 王洪法等, 2007. 黄海特定断面夏秋季大型底栖动物生态学特征. 生态学报, 27(10): 4349—4358
王金宝, 李新正, 王洪法等, 2011. 2005—2009年胶州湾大型底栖动物生态学研究. 海洋与湖沼, 42(5): 728—737
王洪法, 李新正, 王金宝, 2011. 2000—2009年胶州湾大型底栖动物的种类组成及变化. 42(5): 738—752
甘志彬, 李新正, 王洪法等, 2012. 宁津近岸海域大型底栖动物生态学特征和季节变化. 应用生态学, 23(11):3123—3132
古丽娅诺娃 E Y, 吴宝铃, 1963. 胶州湾的底栖生物群落. 见:中国科学院海洋研究所主编. 海洋科学集刊(3). 北京: 科学出版社, 51—61
曲方圆, 于子山, 刘卫霞等, 2009. 北黄海春季大型底栖生物群落结构. 中国海洋大学学报, 39(S1): 109—114
刘卫霞, 2009. 北黄海夏、冬两季大型底栖生物生态学研究.青岛: 中国海洋大学硕士学位论文, 80—84
刘卫霞, 于子山, 曲方圆等, 2009. 北黄海冬季大型底栖动物种类组成和数量分布. 中国海洋大学学报, 39(S1): 115—119
刘录三, 李新正, 2003. 南黄海春秋季大型底栖动物分布现状.海洋与湖沼, 34(1): 26—32
刘瑞玉, 徐凤山, 1963. 黄、东海底栖动物区系的特点. 海洋与湖沼, 5(4): 306—321
李新正, 2011. 我国海洋大型底栖生物多样性研究及展望: 以黄海为例. 生物多样性, 19(6): 676—684
李新正, 王金宝, 王洪法, 2016. 北黄海獐子岛邻近海域大型底栖动物群落结构和季节变化. 海洋科学集刊, 51(1):245—257
李新正, 王洪法, 张宝琳, 2005. 胶州湾大型底栖动物次级生产力初探. 海洋与湖沼, 36(6): 527—533
陈亚瞿, 徐兆礼, 王云龙等, 1995. 长江口河口锋区浮游动物生态研究 I生物量及优势种的平面分布. 中国水产科学,2(1): 49—58
徐 勇, 隋吉星, 李新正等, 2016. 南黄海大型底栖动物群落划分及变化. 广西科学, 23(4): 339—345
隋吉星, 李新正, 王洪法等, 2013. 石岛海域大型底栖生物群落特征. 海洋科学, 37(3): 17—21
Brey T, 1990. Estimating productivity of macrobenthic invertebrates from biomass and mean individual weight.Archive of Fishery and Marine Research, 32: 329—343
Li X Z, Li B Q, Wang H Fet al, 2009. Community structure of macrobenthos in coastal water off Rushan, southern Shandong Peninsula, and the relationships with environmental factors. Acta Oceanologica Sinica, 28(5):81—93
Li X Z, Yu Z S, Wang J Bet al, 2005a. Study on the secondary production of macrobenthos from southern Yellow Sea.Chinese Journal of Applied and Environmental Biology,11(6): 702—705
Li X Z, Zhang B L, Wang H F, 2005b. Secondary production of macrobenthos from the anchovy spawning ground in the southern Yellow Sea. Chinese Journal of Applied and Environmental Biology, 11(3): 324—327
Xu Y, Li X Z, Wang H Fet al, 2016a. Seasonal and spatial variations of macrobenthic community structure and diversity in the South Yellow Sea. Aquatic Ecosystem Health & Management, 19(1): 92—100
Xu Y, Li X Z, Wang H Fet al, 2016b. Status of macrobenthic diversity and distribution in Subei Shoal, China. Aquatic Ecosystem Health & Management, 19(4): 411—419
Xu Y, Sui J X, Yang Met al, 2017. Variation in the macrofaunal community over large temporal and spatial scales in the southern Yellow Sea. Journal of Marine Systems, 173:9—20 Zhang J L, Xu F S, Liu R Y, 2012. Community structure changes of macrobenthos in the South Yellow Sea. Chinese Journal of Oceanology and Limnology, 30(2): 248—255
猜你喜欢
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
今日农业(2020年19期)2020-12-14
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
中国科技信息(2015年2期)2015-11-16
集装箱化(2014年10期)2014-10-31
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
