
干旱胁迫对4种园林绿化树种幼苗光合参数的影响
2017-03-30 05:38谢腾芳
广东农业科学 2017年12期
谢腾芳,薛 立
(1.广州普邦园林股份有限公司,广东 广州 510600;2.华南农业大学林学与风景园林学院,广东 广州 510642)
公路两侧表面土壤水分蒸发快、持水能力弱,不利于植物生长和其他生物活动,生态恢复非常困难[1],因此抗旱性成为公路边坡绿化树种筛选的一项紧迫而极其重要的研究课题。植物对干旱胁迫的应答是多基因、多途径相互作用共同完成的,是一个复杂的多方调控的应激反应[2]。干旱胁迫既影响植物光合器官的形态结构,又对光合作用的生理、生化等过程产生重大影响[3-4]。研究表明,水分对叶片光合作用的影响巨大,即使在轻度水分胁迫下,也容易造成气孔关闭或植物叶片的气孔阻力和叶肉阻力增加,导致植物光合速率下降、生长受到抑制[5]。光合速率和蒸腾速率一定程度上反映植物光合作用和蒸腾作用的强度,而两者的比例水分利用效率直观反映植物的水分利用情况[6]。公路边坡保水能力较差,植被恢复困难,通过了解绿化树种本身在水分胁迫下的光合响应机理,筛选出抗旱能力强的树种,对公路边坡生态恢复具有重要意义。
海南蒲桃(Syzygium hainanense)、紫楠(Phoebe sheareri)、铁冬青(Ilex rotunda)、小叶榕(Ficus microcarpa)是热带、亚热带地区的重要园林绿化树种。尽管有些学者已研究了臭氧与干旱胁迫下海南蒲桃的幼苗光合生理特性[7],阴蔽条件下紫楠幼苗的光合生理特性[8],在镉[9]、SO2[10]、低温[11]胁迫下铁冬青幼苗的抗性以及小叶榕的生理生化[12]和光合生理特性[13]等,但对以上树种在干旱胁迫下光合参数的对比仍未见报道。本研究将海南蒲桃、紫楠、铁冬青、小叶榕等4种幼苗置于温室,采用自然干旱胁迫法模拟干旱环境,测定其光合参数,比较4种树种抗旱能力,探讨各树种干旱胁迫下光合作用响应机理,对于筛选抗旱性强的公路边坡植被恢复树种有一定的参考价值。
1 材料与方法
1.1 试验材料
试验地设在华南农业大学林学院温室,所用苗木为1年生营养袋实生苗,营养袋直径16 cm,深17 cm,基质为1∶1的荷兰土与黄心土。苗木生长基本情况见表1。
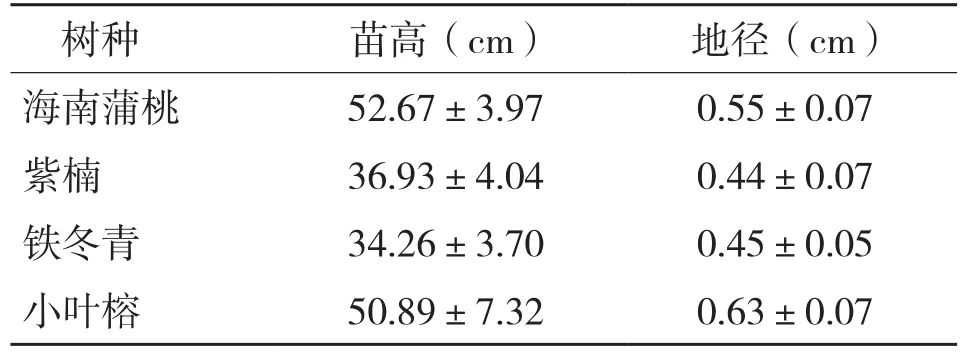
表1 各树种幼苗生长基本情况
1.2 试验方法
采用自然干旱胁迫法对试验幼苗进行0(CK)、4、8、12 d及复水4 d(R4)等5种控水处理。采用LI-6400便携式光合仪(LI-Cor. Inc,美国)测定幼苗针叶在饱和光强1 600 μmol/m2·s下净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci),并计算气孔限制值(Ls)和水分利用率(RWUE)。测定时使用开放气路。空气流速为0.5 L/min,叶温25℃,相对湿度60%,外界CO2浓度(Ca)为 360 μmol/mol。选用幼苗顶部开始的第3~8枚叶中5枚成熟叶进行挂牌标记,测定时间在9:30~11:00进行,所有参数测定6次重复。RWUE和Ls计算公式如下:
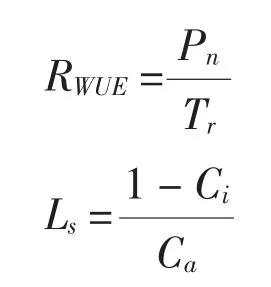
1.3 数据处理
为综合评价不同树种的抗旱性,采用SAS8.1软件系统对数据进行主成分分析[14],比较4个树种的抗旱性。
2 结果与分析
2.1 干旱胁迫对4种园林绿化树种幼苗叶片净光合速率(Pn)的影响
由图1可知,随着干旱天数增加,4种幼苗叶片的Pn都显著下降,均在干旱处理12 d降到最低值,复水4 d后又显著升高。与0 d相比,干旱处理4 d时,4种幼苗叶片的Pn下降范围在20%~22%;干旱处理8 d时,海南蒲桃、紫楠、铁冬青和小叶榕的Pn分别下降了81%、29%、39%和39%;干旱处理12 d时,海南蒲桃、紫楠、铁冬青和小叶榕的Pn降至最低值,分别下降93%、66%、69%和85%。由此可见,干旱处理12 d时,Pn降幅排序为海南蒲桃>小叶榕>铁冬青>紫楠。除海南蒲桃的Pn降幅出现在干旱处理4~8 d外,其他幼苗的Pn在干旱处理8~12 d降幅最明显,可见海南蒲桃的光合系统功能在干旱处理4~8 d后产生的破坏较其他幼苗严重。
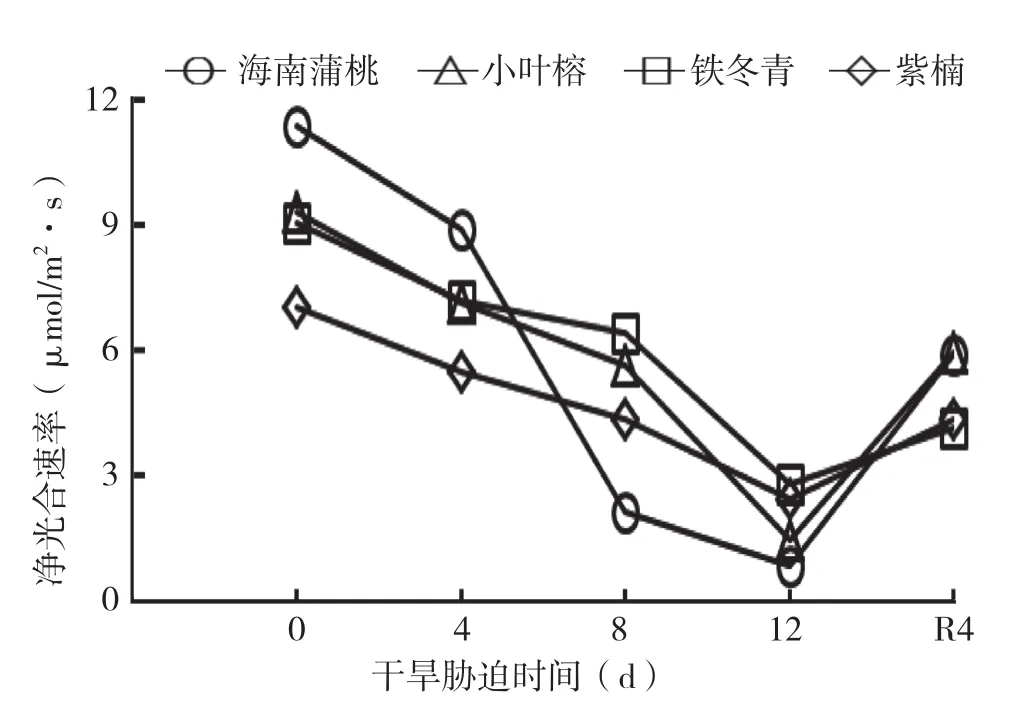
图1 干旱胁迫中4种幼苗叶片净光合速率变化
与0 d相比,复水4 d后,4种幼苗的Pn均显著恢复,Pn降幅排序为铁冬青>海南蒲桃>紫楠>小叶榕,较对照分别下降54%、48%、38%和36%。说明4种幼苗经干旱处理后光合作用虽受抑制,但仍可在复水后逐渐恢复。
2.2 干旱胁迫对4种园林绿化树种幼苗叶片气孔导度(Gs)的影响
由图2可知,随着干旱胁迫的天数增加,4种幼苗的Gs均呈下降趋势,干旱处理12 d时最低,复水后均显著增加,但仍显著低于0 d。与0 d相比,海南蒲桃在干旱处理4、8、12 d时的Gs分别下降24%、92%和93%,紫楠分别下降为34%、62%和76%,铁冬青分别下降29%、45%和87%,小叶榕分别下降4%、66%和93%。由此可见,在干旱胁迫处理12 d时,Gs降幅排序为小叶榕>海南蒲桃>铁冬青>紫楠。此外,海南蒲桃和小叶榕在干旱处理4~8 d时Gs降幅最大,而紫楠则在0~4 d急剧下降,铁冬青在8~12 d时降幅最大。说明铁冬青的Gs较其他幼苗对干旱胁迫的耐受能力强。
与0 d相比,复水4 d后4种幼苗的Gs降幅排序为铁冬青>海南蒲桃>紫楠>小叶榕,分别比对照低47%、36%、17%和10%。说明小叶榕在复水后呈现较强的恢复能力。
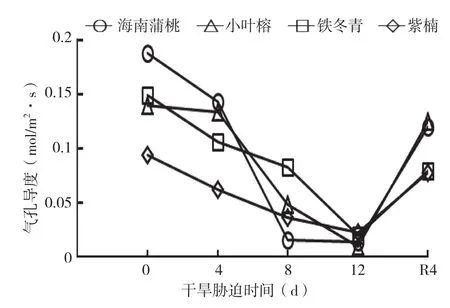
图2 干旱胁迫中4种幼苗叶片气孔导度变化
2.3 干旱胁迫对4种园林绿化树种幼苗叶片蒸腾速率(Tr)的影响
由图3可知,随着干旱天数的增加,4种幼苗叶片的Tr呈下降趋势,至干旱处理12 d降到最小值,复水4 d后显著增加。与0 d相比,在干旱处理8 d时,海南蒲桃、紫楠、铁冬青和小叶榕的Tr分别下降了91%、60%、47%、71%;干旱处理12 d时,则分别下降至93%、75%、88%和93%,降幅排序为小叶榕>海南蒲桃>铁冬青>紫楠。由此可知,除铁冬青的Tr最大降幅出现在干旱处理8~12 d外,其他幼苗的最大降幅均出现在干旱处理4~8 d。说明铁冬青较其他幼苗在干旱胁迫后呈现更强的抗蒸腾能力。
复水4 d后,海南蒲桃、紫楠、铁冬青和小叶榕Tr分别为0 d的55%、34%、51%和31%。说明小叶榕在复水后呈现较强的恢复能力。
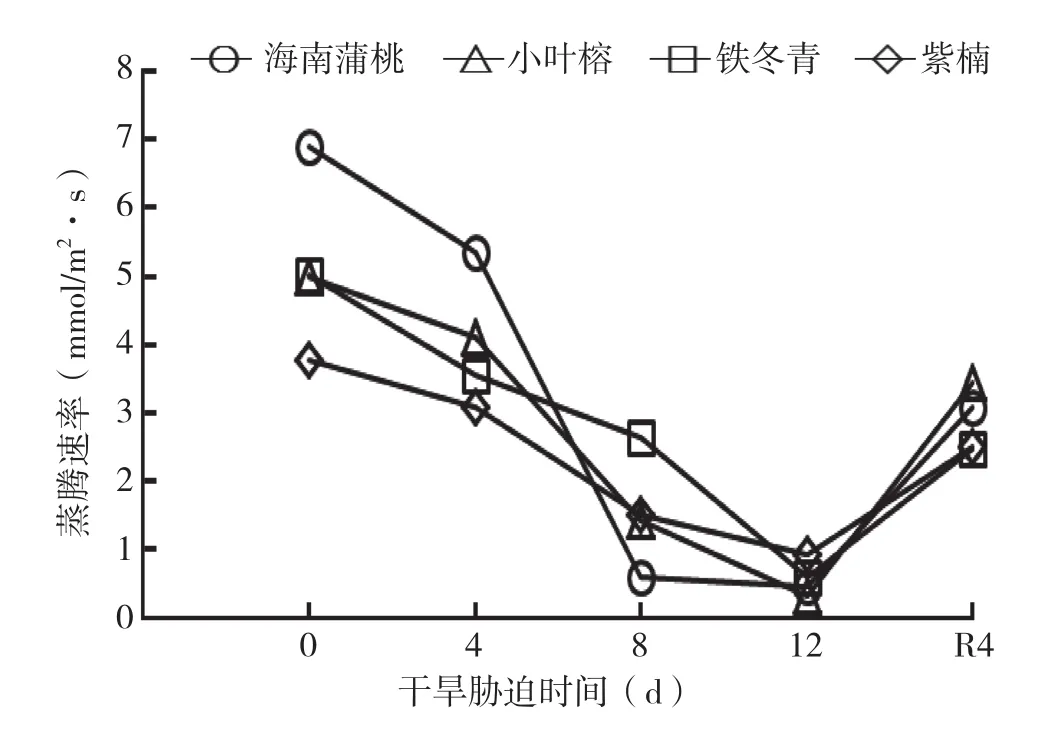
图3 干旱胁迫中4种幼苗叶片蒸腾速率变化
2.4 干旱胁迫对4种园林绿化树种幼苗叶片胞间CO2浓度(Ci)的影响
由图4可知,随着干旱胁迫处理天数的增加,4种幼苗叶片的Ci均呈下降趋势,在干旱处理4~8 d时急速下降,在干旱处理8~12 d降速减缓,逐渐下降至最小值。待复水4 d后,叶片Ci增加,除海南蒲桃外,均显著大于对照,说明海南蒲桃较其他幼苗的恢复能力弱。
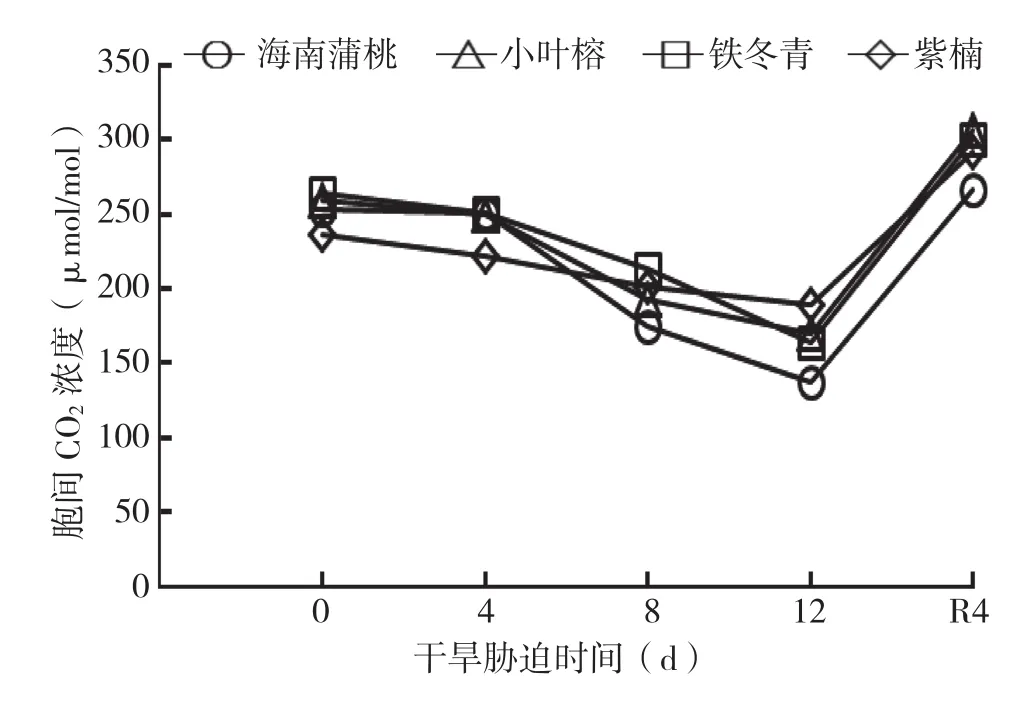
图4 干旱胁迫中4种幼苗叶片胞间CO2浓度变化
2.5 干旱胁迫对4种园林绿化树种幼苗叶片气孔限制值(Ls)的影响
由图5可知,干旱胁迫处理期间,4种幼苗的Ls随着干旱胁迫时间延长而逐渐上升,除小叶榕外,均在干旱处理12 d时增加到最大值。复水4 d后,叶片Ls显著下降,除海南蒲桃外,其余3种幼苗的Ls显著小于对照水平。
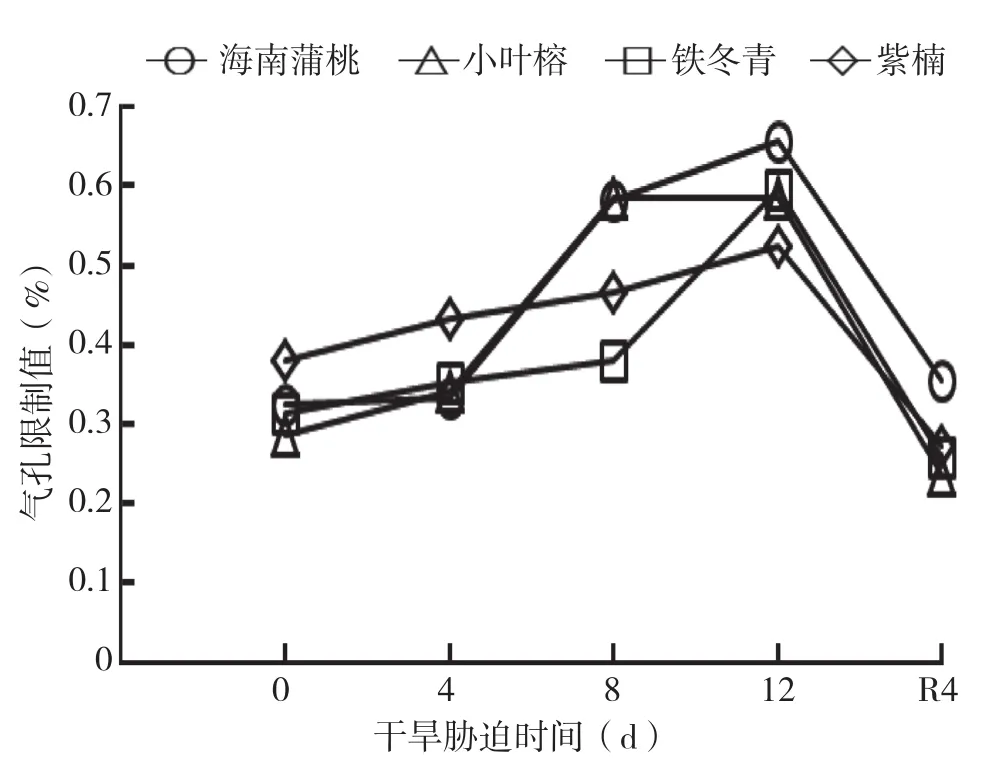
图5 干旱胁迫中4种幼苗叶片气孔限制值变化
2.6 干旱胁迫对4种园林绿化树种幼苗叶片水分利用率(RWUE)的影响
由图6可知,干旱胁迫处理期间,铁冬青的RWUE呈上升趋势,在干旱处理12 d达到最大值,显著大于对照。小叶榕幼苗的RWUE经小幅波动后呈上升趋势,在干旱处理12 d达到最大值。紫楠幼苗的RWUE呈上升趋势,干旱处理8 d达最大值,在干旱处理12 d略有下降。而海南蒲桃的RWUE先逐渐增加,干旱处理8 d达到最大值后,在干旱处理12 d时显著下降至对照水平。与对照相比,复水处理4 d后,除小叶榕外,其余树种均恢复到对照水平。
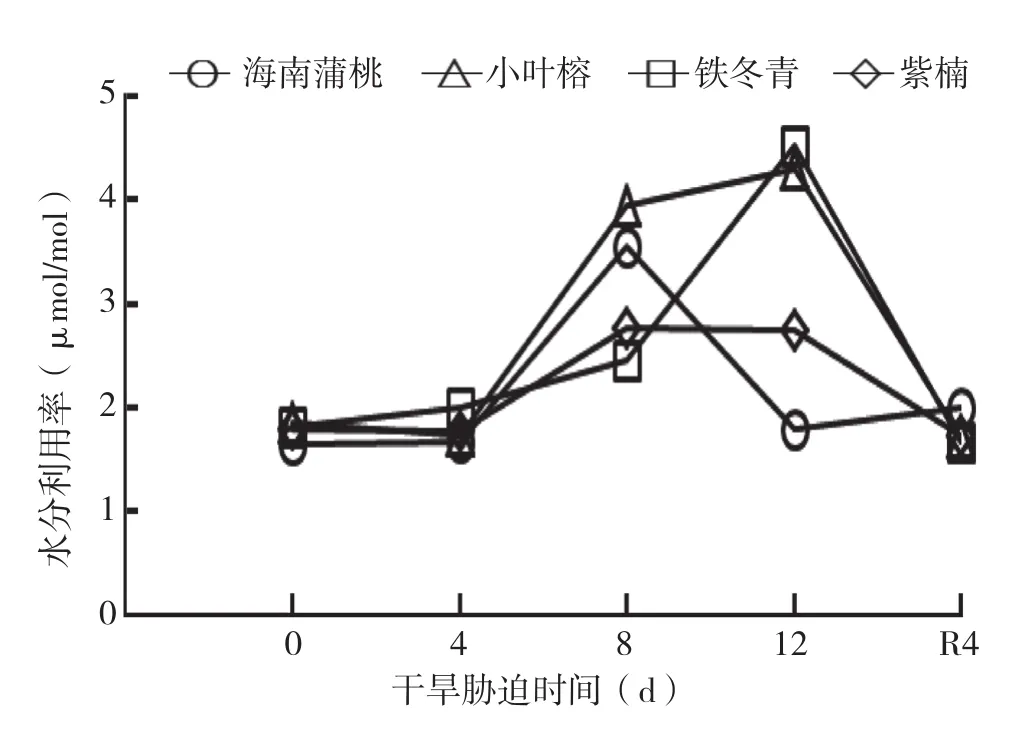
图6 干旱胁迫中4种幼苗叶片水分利用率的变化
2.7 4种幼苗的抗旱能力
已有研究表明,利用主成分分析对幼苗光合参数指标进行抗旱能力评价已获得验证[15-16]。由表3可知,干旱胁迫下4种幼苗的抗旱性大小顺序为:铁冬青>小叶榕>紫楠>海南蒲桃。
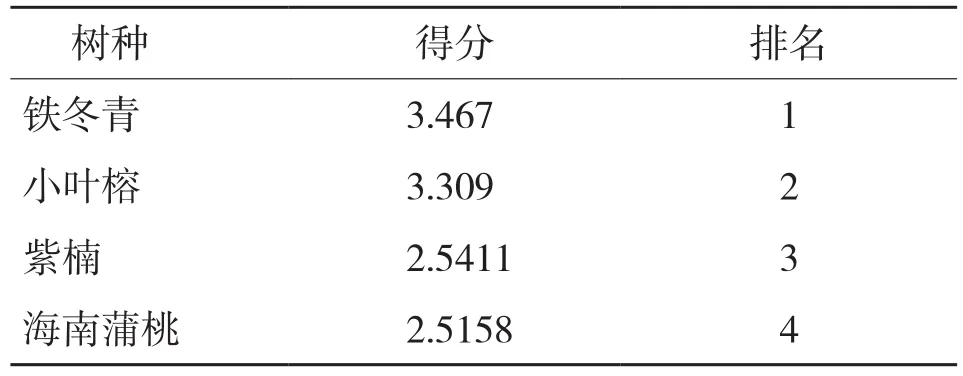
表3 干旱胁迫处理对4个树种光合指标主成分得分及排名
3 结论与讨论
3.1 光合速率、气孔导度和蒸腾速率与植物抗旱性之间的关系
植物受到干旱胁迫时,叶片水分散失和叶片水势下降,导致气孔开度减小,气孔阻力增加,CO2进入叶片受阻,同时光合产物输出变慢,叶片中淀粉水解加强,糖类积累,引起光合速率下降[17-18]。研究表明,Pn、Gs、Tr作为光合作用的重要指标,随干旱胁迫天数的增加而不断降低[15,19-20]。在干旱胁迫期间,4种幼苗的Pn、Gs、Tr持续下降,12 d时显著小于对照,复水处理4 d后增加。与0 d相比,紫楠的Pn、Gs、Tr降幅最小,说明其抗旱能力较强;此外,除光合速率外,铁冬青的Gs、Tr的最大降幅较其他3种植物延迟出现,说明其对干旱胁迫的耐受能力较强;此外,小叶榕在复水处理4 d后的Pn、Gs、Tr较对照降幅最小,说明其在干旱胁迫后的恢复能力较强。
3.2 胞间CO2浓度和气孔限制值变化与植物抗旱性的关系
研究表明,如果Pn和Ci同时减小,且Ls增大,光合速率降低的主要原因为气孔限制[21]。本研究中,干旱胁迫造成4种幼苗的根系导水能力下降而使气孔阻力增加、光合速率减缓,进一步限制CO2吸收,造成Ci下降和Ls增加,说明限制4种幼苗光合作用的主要是气孔限制。此时光合能力降低是短暂的可恢复性的,对植物生理伤害较小[22],说明4种幼苗均具有较强的耐旱能力。
3.3 水分利用效率变化与植物抗旱性的关系
高水分利用率是植物适应干旱环境的一种重要特征[23]。其效率越高,说明植物抗旱能力越强[24]。大量研究表明,干旱胁迫下,许多植物可以通过改变自身的生理生态特性以提高自身的水分利用效率[25]。随着干旱天数的增加,铁冬青和小叶榕的叶片水分利用率持续增加,表明其对水分亏缺具有超强的适应能力,抗旱性较强。但海南蒲桃、紫楠的叶片水分利用率持先增后减,这可能是因为干旱胁迫的时间,超过了植物的忍受能力,植物受到了伤害。
利用主成分分析法可以全面而准确地分析植物各种生理指标的综合抗旱性[14]。本研究运用主成分分析可知,其抗旱性由大到小顺序为:铁冬青、小叶榕、紫楠、海南蒲桃,该结果为其在边坡绿化应用提供参考。但植物的抗旱性是多种指标综合作用的结果[26],因此在边坡绿化引进植物时需结合专业工作经验进行评判筛选与耐旱性相关的关键指标,对完善植物抗旱性评价方法具有重要意义。
[1]朱谦. 山区公路边坡绿化及生态恢复种植材料的选择与处理[J]. 安徽科技,2008,19(9):34-35.
[2]李洁. 植物干旱胁迫适应机制研究进展[J]. 广东农业科学,2014,41(19):154-159.
[3]谢深喜,张秋明,熊兴耀,等. 水分胁迫对柑橘叶片和根系细胞超微结构的影响[J]. 湖南农业大学学报(自然科学版),2008,34(2):168-172.
[4]韩博,李志勇,郭浩,等. 干旱胁迫下5种幼苗光合特性的研究[J]. 林业科学研究,2014,11(1):92-98.
[5]陈建,张光灿,张淑勇,等. 辽东楤木光合和蒸腾作用对光照和土壤水分的响应过程[J]. 应用生态学报,2008,31(6):1185-1190.
[6]苑华臻,郭建斌,刘岩,等. 抗蒸腾叶面肥对国槐幼苗水分利用效率的影响[J]. 西北林学院学报,2017,32(5):29-32.
[7]郝云亭,林敏,薛立,等. 臭氧与干旱胁迫对海南蒲桃和盆架子幼苗光合生理的影响[J]. 安徽农业大学学报,2014,41(2):193-197.
[8]陈模芳,韦小丽,张怡. 紫楠实生幼苗的光合生理特性[J]. 贵州农业科学,2013,14(10):55-58.
[9]胡仲义,章建红. 重金属镉对铁冬青幼苗的毒害效应[J]. 浙江农业科学,2008,23(3):306-309.
[10]张卫强,甘先华,殷祚云,等. 二氧化硫胁迫对园林植物幼苗生理生态特征的影响[J]. 水土保持研究,2012,34(6):247-253.
[11]毛志滨,谢晓金,汤庚国. 7种冬青树种耐低温能力比较[J]. 南京林业大学学报(自然科学版),2006,15(1):33-36.
[12]覃勇荣,潘振兴,黄江滨,等. 桂西北岩溶地区11种桑科植物生理生化特性比较[J]. 山地农业生物学报,2011,30(5):383-390.
[13]夏磊. 重庆市常见园林植物光合生理生态特性与生态效应研究[D]. 重庆:西南大学,2011.
[14]冯慧芳,薛立,任向荣,等. 4种阔叶苗木对PEG模拟干旱的生理响应[J]. 生态学报,2011,31(2):371-382.
[15]杨振意,薛立,郭淑红,等. 干旱对4种幼苗气体交换参数的影响[J]. 中南林业科技大学学报,2012,32(5):67-72.
[16]卢广超,许建新,薛立,等. 干旱胁迫下4种常用植物幼苗的光合和荧光特性综合评价[J]. 生态学报,2013,24(8):7872-7881.
[17]Yordanoyi I,Tsoney T,Goltsey V,et al.Interactiveeffect of water deficit and high temperature on photosynthesis in sunflower and maize plants. 1[J]. Photosynthetica,1997,33(3/4):391-402.
[18]刘志媛,党选民,曹振木. 土壤水分对黄秋葵苗期生长及光合作用的影响[J]. 热带作物学报,2003,24(1):70-73.
[19]张金政,张起源,孙国峰,等. 干旱胁迫及复水对玉簪生长和光合作用的影响[J]. 草业学报,2014,23(1):167-176.
[20]李永秀,申双和,李丽,等. 土壤水分对冬小麦生育后期叶片气体交换及叶绿素荧光参数的影响[J]. 生态学杂志,2012,18(1):74-80.
[21]Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33(5):317-345.
[22]夏江宝,孙景宽,张光灿. 水分胁迫对美国凌霄单叶水分利用效率的影响[J]. 西北植物学报,2009,29(1):154-159.
[23]杨全,孟平,李俊清,等. 土壤水分胁迫对杜仲叶片光合及水分利用特征的影响[J]. 中国农业气象,2010,31(1):48-52.
[24]张向峰,王玉杰,王云琦. 水分胁迫对马尾松光合特性的影响[J]. 中南林业科技大学学报,2012,15(7):58-63.
[25]闫海龙,张希明,许浩,等. 塔里木沙漠公路防护林3种植物光合特性对干旱胁迫的响应[J].生态学报,2010,30(10):2519-2528.
[26]张朋飞,武军艳,孙万仓,等. 干旱胁迫对白菜型冬油菜苗期生理特性的影响[J]. 西北农业学报,2015,13(2):84-90.
猜你喜欢
作物研究(2022年4期)2022-09-21
High Technology Letters(2021年4期)2022-01-09
中国糖料(2021年3期)2021-07-13
种子(2020年11期)2020-12-05
河北果树(2020年4期)2020-11-26
中华建设(2020年5期)2020-07-24
海峡姐妹(2019年1期)2019-03-23
中成药(2018年9期)2018-10-09
小学生必读(低年级版)(2018年6期)2018-09-08
农村百事通(2016年24期)2017-01-07
