
淹水胁迫对火龙果植株形态和生理特性的影响
2017-03-30 05:37韦双双阮云泽
广东农业科学 2017年12期
闫 臻,齐 钊,徐 敏,熊 睿,程 玉,韦双双,阮云泽,汤 华
(海南大学热带农林学院/海南热带生物资源可持续利用国家重点实验室,海南 海口 570228)
火龙果(Hylocereus undatus)属仙人掌科(Cactaceae)量天尺属(Hylocereus Britton et Rose)植物[1],是热带亚热带的名优水果,主要有红皮红肉、红皮白肉和黄皮白肉3种类型。火龙果营养丰富,富含多种有益于人体的物质,深受消费者喜欢。其中,红皮红肉火龙果品种台湾大红具有产量高、肉质细腻、甜度高、耐储存、口感好[2]、不需人工授粉[3]、易种植等特点,受到火龙果种植业与消费者的青睐。近年来,随着不断增加的火龙果消费需求,火龙果的栽培面积日益扩大。海南优越的光照和气候条件,十分有利于发展火龙果种植业[4]。然而,海南地区因台风而引起的强降水以及沿海、沿江地带可能会发生的洪涝灾害都会造成土壤中的含水量过高,而当土壤含水量超过土壤田间持水量时,就会对火龙果造成淹水胁迫,这种非生物胁迫严重威胁了火龙果植株的正常生长发育,极不利于火龙果种植业的发展。但目前国内外对此方面的研究较少,尚未见到相关报道,鉴于此,本研究以台湾大红火龙果品种为材料进行淹水胁迫处理,对相关的形态学指标、生理特性进行研究和对比分析,探索淹水胁迫对火龙果植株的影响,以期为火龙果的涝害防治提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为台湾大红火龙果品种,由海南万钟实业有限公司提供。供试土壤取自海南大学热带农林学院海甸基地。
1.2 试验方法
试验于2017年3~7月在海南大学热带农林学院海甸基地大棚中进行。取火龙果枝条扦插育苗,选取根系良好、生长势一致、无病虫害的火龙果苗进行盆栽1个月,然后进行淹水胁迫处理:将盆栽火龙果苗放入水槽中进行淹水胁迫处理,使水面高出盆栽土壤表面2 cm,试验设6个淹水胁迫处理阶段,分别为淹水10、20、30、40、50、60 d,以不淹水栽培为对照。每个处理3次重复,每个重复10株幼苗。
对未淹水对照及淹水胁迫10、20、30、40、50、60 d处理,每个处理取10株幼苗,测定主根数量、侧根数量、最长根长、气生根数量及生长状态等根系形态指标。对未淹水对照及淹水胁迫10、20、30、40 d处理取样,测定地上部分的超氧化物歧化酶(SOD)活性(氮蓝四唑光化还原法)、过氧化物酶(POD)活性(愈创木酚法)、可溶性糖含量(蒽酮比色法)、可溶性蛋白含量(考马斯亮蓝G-250染色法)、脯氨酸含量(磺基水杨酸提取法)、叶绿素含量(无水乙醇提取的分光光度法)以及地下部分的根系活力(氯化三苯基四氮唑TTC还原法)。
采用Excel 2007进行原始数据整理计算、图表绘制,辅以DPS 7.05进行方差分析及差异显著性检验。
2 结果与分析
2.1 不同淹水时间对火龙果植株形态的影响

表1 不同淹水时间对火龙果植株形态的影响
由表1可知,淹水胁迫10~30 d处理火龙果植株的地上部分在形态上与未淹水时无明显区别;淹水30~40 d时,植株侧茎出现褶皱和萎蔫(图1B,彩插一),主茎基部有病斑出现(图1D,彩插一);淹水40~50 d后植株主茎基部开始腐烂(图1E,彩插一)。火龙果植株的地下部分在淹水胁迫10~20 d后,其形态与未淹水时无明显区别;淹水20 d时,侧根数量开始减少(图2B,彩插一);淹水40 d时,主根开始明显变短(图2C,彩插一)。

图1 淹水胁迫对火龙果植株表型的影响
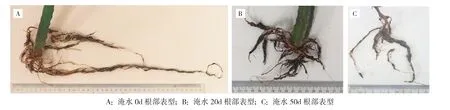
图2 淹水胁迫对火龙果植株根系的影响
2.2 不同淹水时间对火龙果植株地上部SOD、POD活性的影响
由图3可知,火龙果植株地上部SOD活性随淹水时间的延长整体呈先上升后下降再上升的趋势,其中淹水20 d时SOD活性达到第一个峰值,比未淹水的对照增加146.28%;淹水40 d时到达第二个峰值,比对照提高73.36%。火龙果地上部POD活性随淹水时间的延长呈上升趋势,淹水10、20、30、40 d分别比对照增加48.03%、117.70%、209.52%、318.22%。
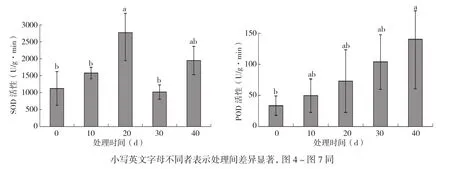
图3 不同淹水时间对火龙果植株地上部SOD、POD活性的影响
2.3 不同淹水时间对火龙果植株地上部可溶性糖、可溶性蛋白含量的影响
从图4可以看出,火龙果植株地上部可溶性糖含量随淹水时间的延长呈先上升后下降的趋势,其中淹水30 d时可溶性糖含量达到峰值,为未淹水对照的10.01倍。图4显示,火龙果地上部可溶性蛋白含量随淹水时间的增加整体呈先上升后下降再上升的趋势,其中淹水20 d时可溶性蛋白含量达到第一个峰值,为对照的1.72倍;淹水40 d时到达第二个峰值,为对照的1.18倍。
2.4 不同淹水时间对火龙果植株地上部脯氨酸含量和叶绿素含量的影响
由图5可知,火龙果地上部脯氨酸含量随淹水时间增加整体呈上升趋势,淹水40 d的脯氨酸含量比未淹水的对照提高120.88%。
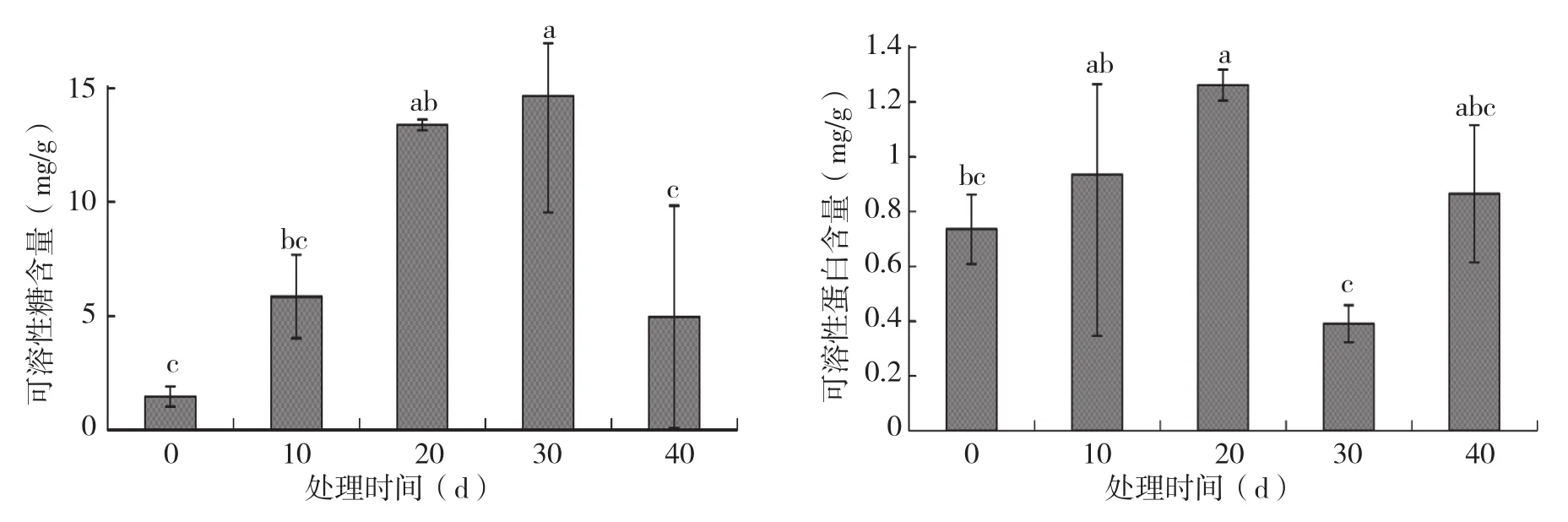
图4 不同淹水时间对火龙果植株地上部可溶性糖、可溶性蛋白含量的影响
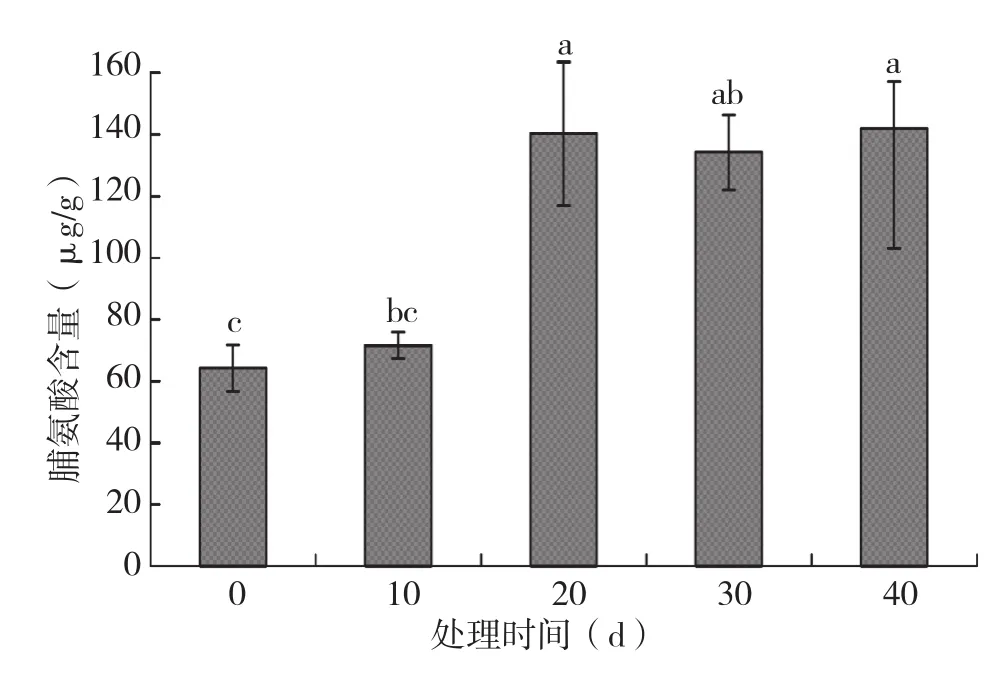
图5 不同淹水时间对火龙果植株地上部脯氨酸含量的影响
由图6可知,火龙果植株地上部叶绿素a和类胡萝卜素的含量随淹水时间增加变化未达到显著水平,叶绿素b含量在淹水30 d时达最小值、仅为对照的0.51。在所有处理中,植株地上部均表现为叶绿素a含量>叶绿素b含量>类胡萝卜素含量。
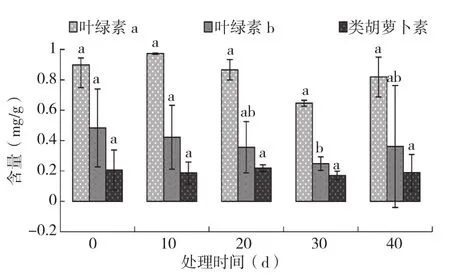
图6 不同淹水时间对火龙果植株地上部叶绿素含量的影响
2.5 不同淹水时间对火龙果植株根系活力的影响
由图7可知,淹水胁迫不同时间火龙果植株的根系活力均显著减弱,淹水10、20、30、40 d后火龙果植株的根系活力分别为未淹水对照火龙果植株的33.21%、38.81%、41.12%、47.34%。
3 结论与讨论
水分是影响着植物生长发育的关键生态因子,过多水分会阻碍植物与大气环境间的气体交换,造成植物受淹组织缺氧,而火龙果必须从环境中获取氧气来维持正常的生理代谢及生长发育过程,所以最终会导致严重减产[5-6]。火龙果属仙人掌科植物,普遍认为其较为耐旱而不耐涝。本研究通过对火龙果进行淹水胁迫处理发现,火龙果植株对淹水胁迫的耐受性已经远远超出试验最初预期的1~3 d。本研究发现,长时间的淹水并未直接导致火龙果植株的死亡,火龙果对淹水胁迫存在较强的耐受性。
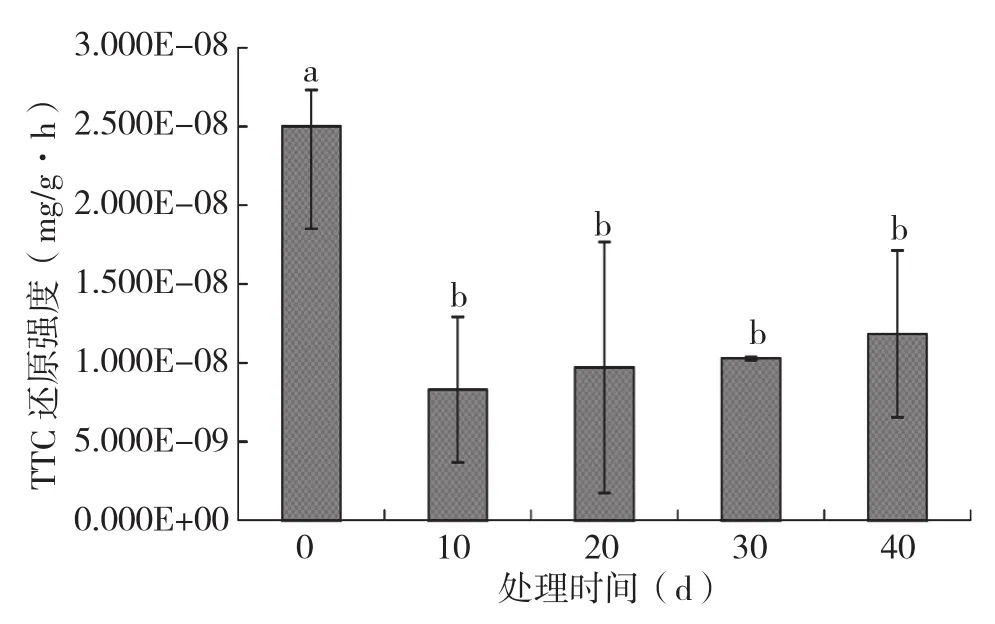
图7 不同淹水时间对火龙果植株根系活力的影响
淹水胁迫使植株的气体扩散受限、叶绿素降解、光合速率下降,为了适应淹水生境,植物形态往往发生改变以保障其正常的生理功能和新陈代谢[7]。本研究发现,火龙果在长期淹水胁迫情况下会产生不定根(气生根),其形态适应机制常表现为产生以下3种形态的不定根:第一类不定根与火龙果正常生境下产生的不定根相似,由侧茎近顶端的部位产生,较短小;第二类不定根同样自侧茎产生,垂入水中,产生放射状的侧根;第三类不定根自根部产生,伸出水面以上,颜色为绿色。不定根大量出现于淹水30 d和60 d处理,但个体差异较为明显,出现大量不定根的侧茎其褶皱情况较未出现不定根的侧茎轻微。有报道指出,不定根的形成本质可能是由于淹水引起生长素和乙烯之间的相互作用,大量不定根的形成有利于保持根系的活力和功能[8-9]。因此,不定根很可能对火龙果适应淹水胁迫有帮助。
火龙果作为一种浅根系植物,其根系往往分布在10~30 cm的土壤浅层,侧根较为发达,然而,火龙果的主根在淹水10 d时就已经开始明显缩短,根系活力明显降低,20 d时侧根已经明显减少。而淹水30 d时,火龙果茎的表型无明显变化,且地上部分叶绿素含量也未发生显著变化,无变黄现象。可见,淹水胁迫主要作用于火龙果根部,导致根系缺氧,影响水分和营养的吸收。
淹水胁迫影响细胞的膜系统及其功能,在涝害发生时植物所产生的一些毒害物质会破坏植物体内活性氧代谢系统的平衡,导致细胞膜损伤,最终导致植物体内的叶绿素降解、电导率上升、内含物外流、脯氨酸积累等一系列现象发生。植物在进化过程中产生了相应的抗氧化防御系统,包括活性氧清除酶类(SOD、POD等)、非酶清除剂等[10]。SOD和POD能够清除过量的活性氧,维持活性氧代谢平衡,保护膜结构[11],从而使植物在一定程度上抵御胁迫。对耐淹水植物而言,其SOD和POD活性会随淹水时间的延长而升高[12],可见,耐水淹的植物具有更高的SOD和POD活性[13]。本研究中火龙果在淹水胁迫过程中,SOD活性先升高后降低再升高,POD活性显著上升,这与刘文革等[14]在西瓜上的研究结果一致。淹水初期,SOD和POD的活性均显著增加,这是火龙果对淹水胁迫的一种应激反应,但随着淹水时间的延长,SOD活性开始下降,这可能是由于长时间淹水胁迫导致抗氧化酶合成受阻[15-17],后期SOD活性的增加可能与不定根的产生有关。
可溶性糖和可溶性蛋白均参与植物的各项生命代谢活动,其含量高,意味着植物体可以积累更多营养物质用于来年的开花结果。研究表明,在淹水前后,植株的存活率与其可溶性糖及淀粉含量显著相关,植株的耐淹性和耐淹后的恢复生长与其碳水化合物含量显著相关[18]。在淹水之前,如果植物有较高的储备量,淹水解除后能够快速恢复生长。逆境条件下,植物体内的脯氨酸含量会明显增加,其积累量与植物的抗逆性有关[19]。
综上所述,火龙果植株对淹水胁迫有一定的耐受能力,淹水胁迫会导致植株的SOD活性、POD活性、可溶性糖含量、可溶性蛋白含量、脯氨酸含量显著增加,叶绿素含量无显著变化,根系活力显著降低;淹水时间过长会导致火龙果根部变短、侧根变少、主茎基部腐烂、侧茎褶皱萎焉或多生气生根。因此,脯氨酸、可溶性糖和可溶性蛋白的积累,抗氧化酶活性的增加,不定根的产生等,可能有利于增强火龙果对淹水胁迫的抗性。
[1]申世辉,马玉华,蔡永强. 火龙果研究进展[J].中国热带农业,2015(1):48-52.
[2]黄樟华,彭代勇,覃群明. 台湾大红火龙果丰产栽培技术[J]. 甘肃农业科技,2017(1):91-92.
[3]台湾大红—— 不用人工授粉,亩产达万斤,最值得种植[J]. 世界热带农业信息,2016(12):45.
[4]李向宏. 海南火龙果产业发展现状及对策//现代果业标准化示范区创建暨果树优质高效生产技术交流会论文汇编[C]. 中国园艺学会,2015.
[5]钟雪花,杨万年,吕应堂. 淹水胁迫下对烟草、油菜某些生理指标的比较研究[J]. 武汉植物学研究,2002(5):395-398.
[6]任佰朝,张吉旺,李霞,等. 淹水胁迫对夏玉米籽粒灌浆特性和品质的影响[J]. 中国农业科学,2013,46(21):4435-4445.
[7]潘澜,薛晔,薛立. 植物淹水胁迫形态学研究进展[J]. 中国农学通报,2011,27(7):11-15.
[8]Wang S G,He L R,Li Z W,et al. A comparative study on the resistance of barley and wheat to waterlogging[J]. Acta Agronomica Sinica,1996,22(2):228-232.
[9]Kaowano N,Ella E,Ito O,et al. Metabolic changes in rice seedlings with different submergence tolerance after desubmergence[J].Environmental and Experimental Botany,2002,47:195-203.
[10]潘澜,薛立. 植物淹水胁迫的生理学机制研究进展[J]. 生态学杂志,2012,31(10):2662-2672.
[11]雷菲,解钰,张文,等. 海水浇灌对不同品种火龙果植株生长和若干生理指标的影响[J]. 热带作物学报,2017,38(6):1052-1057.
[12]陈鹭真,林鹏,王文卿. 红树植物淹水胁迫响应研究进展[J]. 生态学报,2006,26(2):586-593.
[13]叶勇,卢昌义,谭凤仪. 木榄和秋茄对水渍的生长与生理反应的比较研究[J]. 生态学报,2001,21(10):1654-1661.
[14]刘文革,阎志红,王川,等. 西瓜幼苗抗氧化系统对淹水胁迫的响应[J]. 果树学报,2006(6):860-864.
[15]汪贵斌,蔡金峰,何肖华. 涝渍胁迫对喜树幼苗形态和生理的影响[J]. 植物生态学报,2009,33(1):134-140.
[16]何嵩涛,刘国琴,樊卫国. 银杏对水涝胁迫的生理反应(Ⅰ):水涝胁迫对银杏膜脂过氧化作用及保护酶活性的影响[J]. 山地农业生物学报,2000,19(4):272-275.
[17]吴麟,张伟伟,葛晓敏,等. 植物对淹水胁迫的响应机制研究进展[J]. 世界林业研究,2012,25(6):27-33.
[18]Kawano N,Ito O,Sakagami J I. Relationship between shoot elongation and dry matter weight during submergence in Oryza sativa L. and O. glaberrima Steud rice cultivars[J]. Plant Production Science,2008,11(3):316-323.
[19]文晓鹏,郑思成,樊庆杰,等. 火龙果试管苗对干旱胁迫的生理生化反应[J]. 西南大学学报(自然科学版),2013,35(4):8-13.
猜你喜欢
作物研究(2022年1期)2022-11-27
中老年保健(2022年3期)2022-11-21
作物学报(2022年10期)2022-07-21
发明与创新(2021年39期)2021-11-05
现代农业研究(2021年10期)2021-11-02
棉花学报(2020年3期)2020-08-08
海峡姐妹(2020年4期)2020-05-30
科学与财富(2019年15期)2019-10-21
天然产物研究与开发(2018年4期)2018-05-07
天然产物研究与开发(2018年2期)2018-04-04
