
牦牛适应青藏高原环境的组织解剖学研究进展
2017-03-29 03:02丁学智钱娇玲吴晓云梁春年包鹏甲龙瑞军
中国畜牧杂志 2017年3期
杨 超,丁学智,钱娇玲,吴晓云,梁春年,包鹏甲,龙瑞军,阎 萍*
(1.兰州大学草地农业科技学院,草地农业生态系统国家重点实验室,甘肃兰州 730020;2.中国农业科学院兰州畜牧与兽药研究所,甘肃省牦牛繁育工程重点实验室,甘肃兰州 730050;3.兰州大学生命科学学院,甘肃兰州 730000)
牦牛适应青藏高原环境的组织解剖学研究进展
杨 超1,丁学智2,钱娇玲3,吴晓云2,梁春年2,包鹏甲2,龙瑞军3,阎 萍2*
(1.兰州大学草地农业科技学院,草地农业生态系统国家重点实验室,甘肃兰州 730020;2.中国农业科学院兰州畜牧与兽药研究所,甘肃省牦牛繁育工程重点实验室,甘肃兰州 730050;3.兰州大学生命科学学院,甘肃兰州 730000)
高寒、缺氧和牧草营养短缺是高原地区主要的生态限制因子,高原土著动物在长期的适应进化过程中形成了独特的低氧适应策略。牦牛对青藏高原低氧的适应,主要通过特定的生理、代谢及解剖学特征表现出来。近年来,随着低氧环境下从器官到细胞器水平研究的不断深入,牦牛高寒低氧适应在基因水平上的成果较多,但受分析方法和样本数量的限制,低氧适应的候选基因不尽相同,其明确的生理生化表型仍不是很确定。本文从牦牛对氧的运转和利用、高寒胁迫的适应以及应对牧草营养短缺等组织解剖学形态方面的适应性研究进行综述,以期为高原畜牧业的发展、高原动物和人群的疾病防治及揭示高原人群的生理生态适应机制提供重要的借鉴作用。
牦牛;高寒缺氧;解剖学;组织学;适应性
牦牛(Bos grunniens)是青藏高原的主体畜种,更是世界上唯一能在高寒、低氧和牧草营养短缺等极端环境中生存的大型反刍家畜。平均海拔超过4 000 m的青藏高原,是人类生存的极限区,而被誉为“高原之舟”的牦牛,千百年来不仅为当地游牧民提供衣、食、住、行等最基本的生产与生活资料,更是藏传佛教与藏文化传承的重要载体[1]。牦牛以其极高的生态适应性、独特的生物学效率和巨大的经济潜力,对维系整个青藏高原生态系统稳定、社会稳定发展以及国家生态安全起着关键作用[2]。因此,班禅大师曾说“没有牦牛就没有藏民族”。
近年来,牦牛对高原环境的适应性成为研究热点,其中形态结构、生理生化以及分子水平的研究成果颇多。本文主要从牦牛解剖学和组织学角度,对其在高寒环境下形成的特殊适应性进行综述,旨在为牦牛特殊种质资源的保护和进一步品种选育与改良提供新的思考。
1 低氧环境的适应性
氧是机体新陈代谢的重要物质基础,而牦牛分布的地区海拔在3 000~5 000 m。如图1所示,随海拨升高,大气中氧分压逐渐降低。与此同时,吸入气体氧分压(PIO2)、肺泡气氧分压(PAO2)以及动脉血氧分压(PaO2)均逐步下降,这种从大气到机体细胞线粒体的氧传送过程呈瀑布式逐级递减,故称“氧瀑布”[3]。高原哺乳动物不仅对于高海拔低氧环境显示出了极强的忍耐力,还能有效应对伴随氧分压降低而带来的限制有氧活动的潜在风险,其自身通过整体水平代偿机制调节保持机体的血氧稳定和能量需求,从而适应低氧环境。
1.1 呼吸系统解剖学结构 牦牛鼻孔开张幅度大,有利于呼吸。气管短而粗大,断面呈半月形,周径为13~14.5 cm,由50~53块环状软骨组成。气管环状软骨两端的间距大且肌肉发达,有利于频速呼吸,保证牦牛能在高原低压少氧环境下奔跑、行走。同时,牦牛胸腔大,胸廓发育较好,肋骨开张,胸椎和肋骨比普通牛多1~2个。对高原牦牛的肺组织研究发现,高海拔牦牛肺呈灰红色,右肺通常较大,肺脏表面有一层肺胸膜,结缔组织伸入肺内,将实质分成许多完整的肺小叶,其中右肺分为尖叶、中叶、膈叶和副叶,也有的分为4~6叶;左肺多分为尖叶和膈叶,小叶间隔明显而连续[5]。肺脏实质由导气部(包括肺内支气管、小支气管、细支气管和终末细支气管)和呼吸部(包括呼吸性细支气管、肺泡管、肺泡囊和肺泡)组成,肺胸膜、小叶间隔以及肺泡隔共同组成肺间质。如表1所示,牦牛气管长,直径大,肺脏重量大,功能发育良好,两肺不对称系数较高,肺组织中毛细血管网发达,肺泡数多,这些结构特点有利于增加肺通气量和肺血流量,提高牦牛在缺氧条件下的气体交换机能[6]。
1.2 呼吸系统组织学结构
1.2.1 气管的组织结构 牦牛气管壁由黏膜层、黏膜下层和外膜层组成。牦牛气管各段均含有大量的杯状细胞,数量显著高于平原黄牛[7]。牦牛的气管软骨中软骨细胞基质较黄牛丰富,可以供给软骨充分的营养,增加软骨的弹性,有利于牦牛呼吸运动的进行[8]。气管中杯状细胞的存在可以加湿、加温进入呼吸道内的空气,而牦牛气管中杯状细胞的大量存在使其能够很快加热吸入的寒冷空气,适应严酷的高原环境。除此之外,杯状细胞还能够黏附吸入的异物,保证牦牛呼吸道的顺畅。
1.2.2 肺脏的组织结构 在高原低压、少氧的环境下,牦牛由于有特殊的肺泡结构才能够适应低压、少氧环境。牦牛肺泡壁由扁平立方上皮组成,Ⅰ型上皮和Ⅱ型上皮之间紧密连接(图2)。据张勤文[9]报道,生活在高海拔地区的牦牛单位体积内的肺泡数明显多于生活在低海拔地区的牦牛,并且高海拔地区牦牛肺泡隔厚度极显著大于后者。

图1 大气中氧含量随海拔高度变化曲线[4]
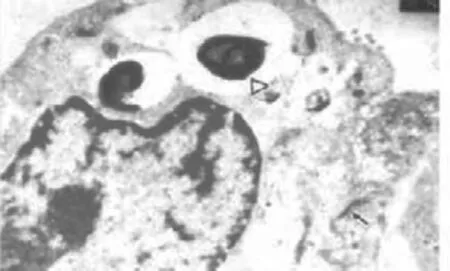
图2 肺泡壁电镜像(×1 000)[12]
由表2可知,不同日龄平原黄牛单位面积内肺泡数显著高于高原牦牛,而单个肺泡面积黄牛却小于牦牛;牦牛单位面积内肺泡面积显著大于平原黄牛;成年牦牛肺泡隔厚度均大于平原黄牛,相应的肺泡隔内毛细血管和弹性纤维等含量丰富。由于牦牛所处环境少氧,为了满足正常呼吸,故其在出生以后肺泡发育迅速,并且单个肺泡发育程度较生活在氧气供应充足的平原黄牛好,随着牦牛自身不断发育,肺泡的数量也迅速增加,以此来增加肺泡面积,扩大气体交换空间,为牦牛适应高原低压少氧环境提供组织学基础。
气-血屏障是肺泡中气体和血液之间的一层分隔组织,它可以让肺泡中氧气进入毛细血管网,同时让组织产生的二氧化碳进入肺内经呼吸道排出体外,而不让毛细血管中血液流出。牦牛肺中气-血屏障由Ⅰ型肺泡上皮、基膜和肺泡隔毛细血管膜组成。陈秋生等[12]研究发现,牦牛肺泡中的气-血屏障非常薄,厚度显著低于猪、狗、绵羊[13]以及平原黄牛,同时牦牛肺泡上皮也非常薄,并且在某些位置出现间断(图3),形成一些上皮小孔。但魏青等[14]经过大量的观察认为,在牦牛肺泡Ⅰ型上皮未发现有间断现象,但是可以看到许多凹陷以及空泡状结构。特殊的气-血屏障以及上皮中空泡状结构减缓了气体交换时的阻力,加快气体进入毛细血管网进行交换的速度,让更多的氧气能够及时运送到各个组织、器官,让牦牛能够在低氧环境下呼吸效率增高,保证其进行正常的生命活动。

表1 牦牛呼吸器官的测定(n=5)[5]
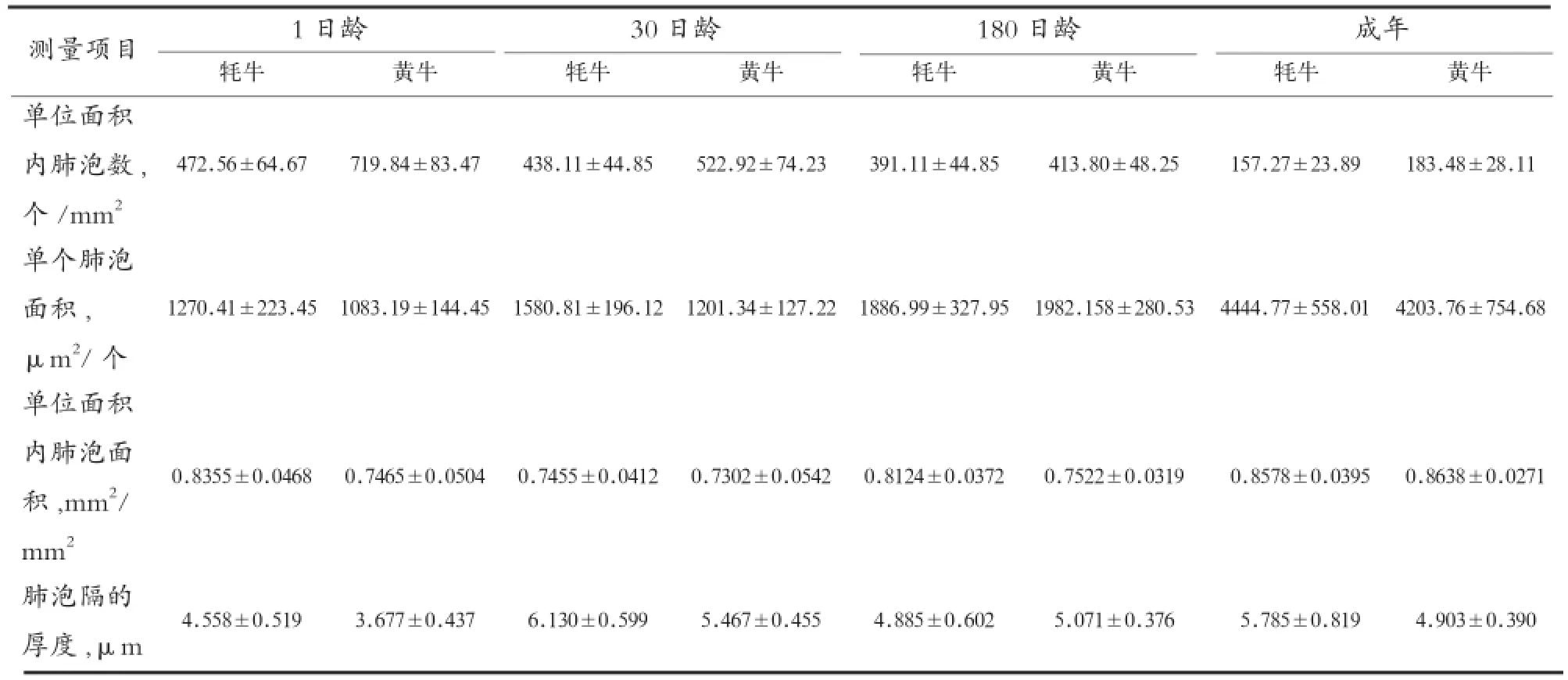
表2 不同日龄高原牦牛和平原黄牛肺脏显微结构相关测量值比较[10-11]

图3 肺泡壁电镜像(×10 000)[12]
2 高寒环境的适应性
高原地区昼夜温差大,常年温度较低,平均气温为0℃,牦牛能在-30℃甚至更低的温度下正常生活。随着海拔的不断升高,牦牛的绝食产热平稳不变,是由于牦牛的体型较小,而且比较紧凑,四肢比较短,体表褶皱较少,这样明显减少了单位面积的散热量[15]。
2.1 皮、毛的解剖学结构 牦牛皮肤较厚,青海牦牛皮肤平均厚度为6.50 mm,天祝白牦牛皮肤平均厚度为6.23 mm。牦牛全身被毛密而长,由粗毛、两型毛和绒毛组成,属于混合型被毛。在寒冷季节,粗毛间生长出绒毛和两型毛,身体各突出和着地部位长有粗而长的裙毛,其间充满了空气,比单类型被毛的空气层厚,形成了天然的隔热层,有利于抵御高寒[16]。
2.2 皮肤的组织学结构 牦牛皮肤的结构主要由表皮和真皮构成。表皮由角质层、颗粒层、棘细胞层和基底层构成。牦牛表皮层极其薄,并且角质化程度低,各部位厚度不同;基层细胞内含有大量色素颗粒,可防止强紫外线照射[17]。牦牛真皮层特别厚,由乳头层和网状层构成,真皮层中含有丰富的胶原纤维、相对较少的网状纤维和弹性纤维,在成年牦牛真皮中还含有大量的脂肪细胞,对阻止体内热量的散失有重要作用。另外,Zhang等[18]发现,青海牦牛皮肤中的血管主要在真皮的乳头层和网状层有少量分布,减少血液对流散热。牦牛竖毛肌比较发达,其一端附着于毛囊,另一端位于真皮乳头层附近。在寒冷环境中,竖毛肌收缩,使毛干直立,形成一层绝热的空气层,减少散热。牦牛的毛囊由一个初级毛囊和4~9个次级毛囊组成,如此多的次级毛囊将在寒冷季节产出大量绒毛夹在粗毛之间,为牦牛维持正常体温。岳静[19]研究发现,牦牛的皮肤厚度随年龄变化逐渐增加,成年牦牛背部皮肤的厚度大于其他部位,而且在其真皮中,存在脂肪细胞浸润的现象,这样可以让其抵御寒冷。另外,牦牛汗腺不发达,故其排汗能力较弱,减少了热量的散失。综上所述,从组织学角度来看,牦牛脂肪浸润细胞、次级毛囊发达、汗腺不发达等特点使其能够抵御寒冷的侵袭,适应严酷的高原环境。
3 应对牧草营养短缺的适应性
在高原地区,由于太阳辐射较强、温度较低、无霜期较短,使得牧草生长期较短,牧草生长受到限制,营养价值较低。为了更好地适应高原营养短缺的状况,牦牛经过不断地进化,形成了一套特点突出的消化系统,但是牦牛皱胃在解剖学结构上与普通牛相似,没有明显差异,仅是分泌腺体较多。3.1 口、牙齿的解剖学结构 牦牛的唇厚而灵活,口裂小,可以啃食5 cm高的牧草。其上唇黏膜比较光滑,靠近唇缘有许多小而圆的乳头,向口角处乳头数增多且由圆顶变成锥体,下唇近口角处的乳头呈锥状,尖端向后。牦牛牙齿坚硬,门齿齿面较宽呈铲形齿冠,有利于切割牧草。
3.2 舌的解剖学结构 牦牛的舌与黄牛相似,分为舌根、舌体、舌尖3部分,其中舌体上有很多肌肉发达的突起,表面和侧面分布着丝状、轮廓、菌状、豆状、圆锥状乳头(图4)。食入口中的食物在突起上的丝状乳头、圆锥状乳头、豆状乳头以及上颚共同作用下,经研磨变碎。圆锥状乳头比较发达,顶端被角质化上皮覆盖而且数量较多,有很好的机械研磨功能。豆状乳头位于舌突起表面,只是起到单纯的机械消化作用。丝状乳头在牦牛舌突起表面有少量分布,在形状上和总体数量上跟其他动物没有太大区别[20],它可以将食物固定、理顺,除此之外,还可以清理牙齿和下颚之间的杂物。丝状乳头顶端被连续的、有规律的锯齿形上皮环绕,提高了其机械消化功能。菌状乳头底部为圆形,外围被丝状乳头环绕的结构为其提供了保护。轮廓状乳头有简单型和复合型两种,在牦牛舌体上主要为复合型,分布较为广泛[21]。相比之下,黄牛舌较短,舌体上突起不发达,圆锥状乳头分布较少,豆状乳头表面没有被角质化上皮覆盖,机械功能不及牦牛(图5)。牦牛在采食时,主要用口唇,不直接用舌来接触食物。因为在青藏高原寒冷的冬天,气温平均-30~-40℃,如果牦牛用舌将草拽进口中,可能会因为霜吸热而使舌受伤。
3.3 舌的组织学结构 从组织学角度看,牦牛舌上的各类乳头表面均附有较厚的角质化上皮,保证了其一定的机械功能;在圆锥状、豆状、丝状、菌状乳头上无味蕾分布,可以观察到一些腺体存在,而在轮廓状乳头上有味蕾分布,各类乳头上有大量的黏液分泌小孔分布[22]。黄牛舌上各类乳头角质层较薄,分布有大量的味蕾,味觉更敏感。因此,牦牛由于味觉较差、黏液分泌孔分布较多等特点,对于枯黄期纤维含量高的饲草产生了一定的适应性。
3.4 前胃的解剖学结构 牦牛与其他反刍动物一样有4个胃。瘤胃、网胃和瓣胃被称为前胃。瘤胃最大,占全部胃容积的75%~78%,成年牦牛瘤胃容积可达到42~45 L。瘤胃上皮黏膜大部分为棕色或黑色,背囊黏膜呈灰白色;其上皮分布有大量的乳头,腹囊乳头最为发达,表现出密度大、长度大的特点;背囊乳头相对较少,长度较小,但宽度较大。网胃处于瘤胃前方,是前胃中最小的,其上皮黏膜形成许多褶皱,并且相互连接成四边、五边或六边形网格。在网格上还有许多次级褶皱,形成许多小网格,在小网格底部分布着大量角质化乳头。瓣胃内部有很多叶片状结构,食糜流入叶片状结构之间,在瓣胃收缩下,进行机械消化。研究发现,牦牛网瓣孔直径显著小于瓣胃和皱胃孔,并且随着季节变化而变化[23]。在暖季,牧草中纤维量下降的时候,网瓣孔变大,加快消化速度;在秋季和冬季,由于草场中牧草开始进入枯黄期,纤维素和木质素含量逐渐升高,为了保证其在瘤胃和网胃中充分发酵,网瓣孔直径变小,减慢向瓣胃流动的速度,使纤维物质被充分降解,产生更多的挥发性脂肪酸来供能,以适应冬季牧草营养不足的情况。

图4 牦牛舌解剖结构[22]

图5 黄牛舌解剖结构[22]
3.5 前胃的组织学结构 在冬季,牦牛采食的牧草中粗纤维含量较高,经瘤胃发酵产生大量挥发性脂肪酸,会刺激瘤胃上皮乳头的发育[24],使得瘤胃表面扩张系数增大。因此,瘤胃中乳头较发达,单位面积内乳头数量比放牧条件下的普通牛多,挥发性脂肪酸的吸收面积也随之增大。瘤胃上皮为复层扁平上皮,角质化程度较高,具有很强的机械保护作用,这是牦牛能够采食大量粗饲料,适应恶劣环境的消化特征之一。牦牛瘤网胃上皮中,肌层非常发达(图6-7),这种结构保证了一定的机械消化强度,增强了牦牛的消化能力,使其能够耐粗饲[25]。
3.6 小肠的解剖学结构 肠道是消化和吸收营养物质的主要场所,特别是小肠,由十二指肠、空肠和回肠组成。牦牛在各个发育阶段中,回肠的长度最短,空肠最长,各段长度与平原黄牛差异不显著;但是其十二指肠和回肠长度均显著小于同龄的双峰驼,空肠长度则极显著大于双峰驼[26]。同时,小肠内壁黏膜上形成了许多皱襞和绒毛,它们在消化和吸收营养物质的过程中起重要作用。
3.7 小肠的组织学结构 小肠的绒毛长度、隐窝深度以及肌层厚度等都是衡量其消化吸收功能的重要指标,此外绒毛长度和隐窝深度比值(V/C值)也是衡量其消化吸收能力的重要指标[27]。研究发现,V/C值能够反映出小肠的功能状态,当比值上升表明小肠的消化吸收功能增强,有利于动物的生长;比值下降,说明其消化吸收能力下降[28]。小肠各段的绒毛高度会影响其消化吸收功能,而黏膜厚度则会影响营养物质的吸收和转运速度,黏膜中肌层厚度会对小肠的蠕动和机械消化产生影响[29]。牦牛小肠中空肠绒毛高度最大,隐窝最深;十二指肠肌层厚度最大,绒毛高度/隐窝深度的比值最高;3段小肠绒毛高度、隐窝深度、V/C值和肌层厚度之间差异较大[30]。由表3可知,在5个不同牛种中,牦牛的空肠绒毛长度最大,其吸收表面积也最大;隐窝深度显著大于其他4种牛,说明其隐窝上皮成熟速度下降,分泌功能变弱,具有较大的迁移和分化潜能,迁移到绒毛上皮补充正常脱落的细胞增多;肌层厚度最大,其节律性收缩强度最大,机械消化效率最高[31]。牦牛随着日龄的增加(1~30 d),小肠黏膜结构也随之发生变化,表现为各段绒毛长度下降,隐窝深度增加,黏膜肌层厚度增加,V/C值下降;30 d后,随日龄增加,小肠绒毛长度增加,黏膜肌层厚度增加,隐窝深度减小,V/C值上升。相比之下,黄牛在各个阶段均表现出小肠各段绒毛长度、隐窝深度不断减小,V/C值不断降低,黏膜厚度和肌层厚度增加[32]。通过对比发现,随着牦牛不断地发育,其小肠逐渐发达,V/C值不断升高,对于营养物质的消化吸收能力不断增强,而黄牛却恰恰相反。这种变化趋势能够有效地说明牦牛在小肠形态上对高原牧草营养短缺产生了适应性,吸收营养物质的能力更强,因此它能在牧草营养短缺的环境下正常生活。
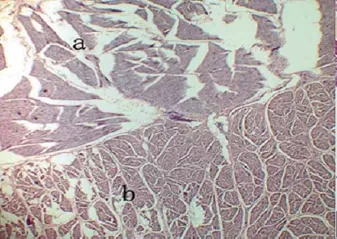
图6 瘤胃肌层(HE×40)[25]

图7 网胃肌层(HE×40)[25]

表3 不同品种成年牛空肠组织相关测量值比较[30,32-33]
4 问题与展望
目前,牦牛对青藏高原地区高寒、低氧、营养短缺环境在解剖学与组织学方面的特殊适应性已被较为深入地研究,但是其适应性机制是一个较为复杂的体系,有很多问题仍旧不清楚,如其解剖学、组织学方面的适应性与机体的分子作用机理是如何协同作用的有待进一步探讨。牦牛瘤胃对于挥发性脂肪酸的吸收效率与其瘤胃内部结构以及瘤胃上皮中转运载体的种类和数量之间的相互作用关系未见详细报道。另外,有大量研究表明,牦牛对高原环境的适应性不仅仅在于解剖学与组织学方面,在氮素循环机制、瘤胃微生物、肠道微生物及微生物宏基因组学等角度均表现出对高寒地区的特殊适应性,但它们与牦牛的解剖学、组织学适应性的共同作用途径以及调控网络尚不清楚。
针对以上问题,今后应当利用生物信息学和先进的分子生物学技术来研究牦牛适应性在解剖学、组织学以及分子机理的联合作用机制。应用蛋白质组学、基因组学以及免疫学等手段对瘤胃上皮中挥发性脂肪酸的转运载体数量、种类以及转运途径进行深入探讨,结合微生物宏基因组学来说明牦牛能适应冷季牧草营养短缺的微生物消化特点和脂肪酸吸收利用特点。同时,还可利用上述技术手段来研究牦牛肠道结构、肠上皮细胞中与氨基酸、小肽等物质的吸收和转运相关载体的分布特点。以期将牦牛对青藏高原环境在各方面的特殊适应性整合成一个完整的系统,为牦牛特殊种质资源的保护和进一步品种选育与改良提供新的思考。
[1] Wiener G, Han J L, Long R J. The Yak second edition [M]. FAO: Regional Office for Asia and the Pacif i c, 2003:460.
[2] Long R J, Apori S O, Castro F B,et al. Feed value of native forages of the Tibetan Plateau of China[J]. Anim Feed Sci Tech, 1999, 80(2):101-113.
[3] Storz J F, Runck A M, Moriyama H,et al. Genetic dif f erences in hemoglobin function between highland and lowland deer mice[J]. J Exp Biol, 2010,123(15):2565-2574.
[4] Beall C M. Two routes to functional adaptation: Tibetan and Andean high-altitude natives [J]. Proc Natl Acad Sci USA, 2007,104 Suppl 1(20):8655-8660.
[5] 李诗洪. 试谈牦牛的适应性[J].中国牦牛, 1985, (3):22-25.
[6] 崔刚, 陆天才, 李洪安, 等. 牦牛心肺的解剖学特点[J].中国牦牛, 1991, (2):29-30.
[7] 魏青, 俞红贤, 张勤文, 等. 高原牦牛气管组织学结构特征[J].黑龙江畜牧兽医, 2013, (15):153-154.
[8] 成令忠, 钟翠平, 蔡文琴. 现代组织学[M].上海:上海科学技术文献出版社, 2003:20-100.
[9] 张勤文. 不同海拔地区牦牛的肺组织形态学比较[J].中国兽医科技, 2004, 34 (2):73-74.
[10] 魏青. 不同发育阶段高原牦牛和平原黄牛肺泡显微结构的研究[D].青海:青海大学, 2007:33-39.
[11] 魏青, 俞红贤, 张勤文, 等. 不同发育阶段黄牛肺泡组织结构的研究[J].青海大学学报:自然科学版, 2010, (6):1-6. [12] 陈秋生, 冯霞, 姜生成. 牦牛肺脏高原适应性的结构研究[J].中国农业科学, 2006, (10):2107-2113.
[13] Meban C. Thickness of the air-blood barriers in vertebrate lungs[J]. J Anat, 1980, 131(2): 299-307.
[14] 魏青, 俞红贤, 张勤文, 等. 高原牦牛肺泡组织结构的观察和比较[J].黑龙江畜牧兽医, 2011, (9):47-49.
[15] 郭宪, 杨博辉, 李勇生, 等. 牦牛的生态生理特性[J].中国畜牧杂志, 2006, (1):56-57.
[16] 施奇静, 王静, 孙军平, 等. 中国牦牛资源保护及可持续利用:驯化与品种选育[J].家畜生态学报, 2016, 37(1):81-85.
[17] 欧阳熙, 王茜飞. 牦牛耐寒力的研究—牦牛皮肤组织学结构的初步观察[J].西南民族大学学报:畜牧兽医版, 1984, (3):13-21.
[18] Zhang Q W, Yu H X, Jing H X,et al. Study on the Datong yak’sskin development [J]. Agri Sci Tech, 2010, 11(9-10) :118-120.
[19] 岳静. 不同年龄牦牛皮肤的组织结构观察[D].兰州:甘肃农业大学, 2013:18-25.
[20] Ⅰwasaki S. Evolution of the structure and function of the vertebrate tongue[J]. J Anat, 2002, 201(1):1-13.
[21] Kumar S, Bate L A. Scanning electron microscopy of the tongue papillae in the pig (Sus scrofa)[J]. Microsc Res Tech, 2004, 63(5):253-258.
[22] Shao B, Long R, Ding Y,et al. Morphological adaptations of yak (Bos grunniens) tongue to the foraging environment of the Qinghai-Tibetan Plateau [J]. J Anim Sci, 2010, 88(8):2594-2603.
[23] 王郑媛.牦牛胃的结构与功能研究[D].兰州:兰州大学, 2005:12-14.
[24] Beiranvand H, Ghorbani G R, Khorvash M,et al. Ⅰnteractions of alfalfa hay and sodium propionate on dairy calf performance and rumen development[J]. J Dairy Sci, 2014, 97(4):2270-2280.
[25] Wang J B, Li H Y, Zhang L,et al. Histomorphometric Charac -terization of Forestomach of Yak (Bos grunniens)in the Qinghai-Tibetan Plateau[J]. Ⅰnt J Mophol, 2014, 32(3):871-881.
[26] 祁珊珊.双峰驼小肠黏膜免疫的形态学特征及其抗感染能力的研究[D].兰州:甘肃农业大学, 2009:15-20.
[27] 李可洲,李宁,黎介寿,等.短链脂肪酸对大鼠移植小肠形态及功能的作用研究[J].世界华人消化杂志, 2002, (6):720-722.
[28] Beers-schreurs H M, Nabuurs M J, Vellenga L,et al. Weaning and the weanling diet influence the villous height and crypt depth in the small intestine of pigs and alter the concentrations of short-chain fatty acids in the large intestine and blood[J]. J Nutr, 1998, 128(6): 947-953.
[29] Kato Y, Yu D, Chwart M Z. Glucagon like peptide-2 enhances small intestinal absorptive function and mucosal mass in vivo [J]. J Pediatr Surg, 1999, 34(1):18-21.
[30] 左愈臻.牦牛小肠结构特点及黏膜免疫功能细胞分布变化研究[D].兰州:甘肃农业学,2011:18-27.
[31] Owens D, Mcgee M, Boland T. Ef f ect of grass regrowth interval on intake, rumen digestion and nutrient fl ow to the omasum in beef cattle [J]. Anim Feed Sci Tech, 2008, 146(1-2) :21-41.
[32] 薛乾.大通牦牛小肠黏膜组织结构及黏膜免疫相关细胞的发育学研究[D].青海:青海大学, 2011:11-17.
[33] 王利民,孙青松,张惠,等. 3个品种肉牛小肠组织结构差异性研究[J].吉林农业大学学报, 2010, (3):326-329.
Research Progress on Adaptation on the Histology and Anatomy in Yak(Bos grunniens) in Qinghai-Tibetan Plateau
YANG Chao1, DⅠNG Xue-zhi2, QⅠAN Jiao-ling3, WU Xiao-yun2, LⅠANG Chun-nian2, BAO Peng-jia2, LONG Rui-jun3, YAN Ping2*
(1.State Key Laboratory of Grassland and Agro-Ecosystems, College of Grassland Agriculture Science and Technology, Lanzhou University, Gansu Lanzhou 730020, China; 2. State Key Laboratory of Yak Breeding Engineering, Ⅰnstitute of Husbandry and Pharmaceutical Sciences, Chinese Academy of Agricultural Sciences, Gansu Lanzhou 730050,China; 3. School of Life Sciences, Lanzhou University, Gansu Lanzhou 730000, China)
Ⅰn high altitude areas, there are so many negative factors to life, especially severe cold and hypoxia. Nevertheless, the indigenous animals own a unique adaptive strategy to adapt the hypoxia condition by long time evolution. The adaptation of the yak to hypoxia in the Qinghai-Tibet Plateau is mainly manifested by the specific physiological, metabolic and anatomical features. Recently, many achievements have obtained on the adaption to severe cold and hypoxia at the genetic level of yak with the continuous study of the level from organ to organelle. However, the candidate genes involved in hypoxia adaptation are not the same due to the limitation of the analytical methods and the number of samples, and the specif i c physiological and biochemical phenotypes remain unclear. The objective of this paper was to summarize the adaptability of the yak to high attitude and hypoxia condition on morphology and anatomy of tissue, which may provide a reference for the development of highland animal husbandry, disease prevention and control, as well as reveal the mechanism about oxygen transportation and utilization, the adaption to cold stress and innutritious forage.
Yak; Severe cold and hypoxia; Anatomy; Histology; Adaptabilit
S823.8
A
10.19556/j.0258-7033.2017-03-018
2016-08-12;
2016-09-01
甘肃省杰出青年基金计划项目(1308RJDA015);国家自然基金委国际合作与交流项目(31461143020)
杨超(1992-),男,甘肃酒泉人,硕士,研究方向为牦牛高原适应性,E-mial:yangch15@lzu.edu.cn
* 通讯作者:阎萍,女,山西人,博士生导师,研究方向为牦牛遗传育种,E-mail:pingyanlz@163.com
猜你喜欢
中国临床解剖学杂志(2022年6期)2022-12-30
青海湖(2022年3期)2022-06-09
滇池(2022年5期)2022-04-30
散文诗(2021年22期)2022-01-12
世界科学技术-中医药现代化(2021年7期)2021-11-04
海洋通报(2021年2期)2021-07-22
小哥白尼(野生动物)(2020年9期)2021-01-18
山西教育·招考(2019年3期)2019-09-10
中国兽医杂志(2019年2期)2019-06-25
武术研究(2019年12期)2019-04-20
