
杂草稻资源研究现状及利用进展
2017-03-27 21:14刘丹柴永山孙玉友魏才强解忠李洪
湖北农业科学 2017年2期
刘丹+柴永山+孙玉友+魏才强+解忠+李洪亮+张巍巍+孙国宏+侯国强
摘要:杂草稻是一类具有杂草特性的野生资源,常与栽培稻一起伴生在稻田里,对水稻产量和品质有着重要的影响。由于长期生长在野生状态中,经受着各种不良环境的选择,杂草稻具有极强的抗逆能力。因此,对杂草稻植物学特征、分类、起源、危害与防治以及优异基因的挖掘等5个方面进行了综述,旨在为更深入了解和利用杂草稻野生资源奠定基础。
关键词:杂草稻;种质资源;杂草危害;抗逆性
中图分类号:S451 文獻标识码:A 文章编号:0439-8114(2017)02-0201-04
DOI:10.14088/j.cnki.issn0439-8114.2017.02.001
杂草稻(Weedy Rice)又名杂草型稻或杂草种系,早在1846年美国就已有发现,当地居民因其果皮呈红色而称其为“红稻”(Red Rice)。近年来,几乎世界上所有的稻作生产区都有杂草稻的发生[1,2]。调查发现,杂草稻常与栽培稻伴生在稻田里,与栽培稻具有十分相似的形态特征,通常田间很难进行鉴别,只是出穗比栽培稻早,边成熟边落粒,第二年成为田间的恶性杂草[3]。
近年来,由于农村劳动力人口的大量流失以及水稻精简栽培技术的不断推广,中国杂草稻发生日益频繁,已经严重影响到水稻产量和国家粮食安全。有数据统计,中国黑龙江省仅2 000年因杂草稻造成的经济损失就有3亿~4亿元人民币[4]。此外,杂草稻是研究栽培稻起源和进化的重要材料,对于水稻遗传改良也具有重要作用。大量研究表明,杂草稻长期处于野生环境,具有丰富的遗传基础和变异类型,且有着许多优良的抗性性状,诸如抗寒性、抗旱性、耐盐碱性等[5-13]。研究和利用杂草稻资源,有助于开展栽培稻的抗性改良和丰富栽培稻日益狭窄的遗传基础。因此,本研究从杂草稻的形态特征、分类、起源、危害与防治以及优异基因挖掘等方面入手,对杂草稻的国内外研究现状进行了综述,旨在为进一步了解杂草稻,有效防治和利用杂草稻奠定基础。
1 杂草稻的植物学特征
杂草稻的生育期较早,田间表现为抽穗开花较早,落粒性强,边成熟边落粒。杂草稻成熟后,茎秆、芒以及颖尖等器官由于花色素类物质的沉积而表现为多种颜色,具有丰富的变异类型[14,15]。研究表明,杂草稻的株高存在显著差异,其变化范围在62~192 cm之间[7,16,17]。杂草稻的穗部形态多样,穗型多为中等或开放,颖果中型或长型,多有长芒,内外稃稻草色,黑色,具褐色沟纹或斑点,果皮红色或白色[18]。杂草稻的分蘖能力、单株穗数以及芒长等生物学特性变异较大,不同地区杂草稻间差异明显。孙建昌等[19]研究表明,宁夏杂草稻农艺性状的表型变异较小,变异类型较单一,大多数农艺性状表现与当地栽培稻相近或相似。而马殿荣等[20]研究表明,辽宁省杂草稻的分蘖能力、单株穗数及芒长等生物学特性变异较大。
整体来看,杂草稻与栽培稻存在形态学差异[21]。前人的研究表明,尽管在苗期杂草稻与栽培稻很难区分,但大田移栽后,进入分蘖期,杂草稻就表现出了一些与栽培稻相区别的特殊性状,如生长快、叶片两面有长毛、植株高、容易落粒、有长芒等[22]。袁晓丹等[23]对几种东北地区杂草稻的形态性状特征进行了调查分析,认为杂草稻的株高伸长速度基本上快于普通稻,分蘖能力均强于普通水稻,杂草稻子粒褐色居多,且有芒,芒较长,成熟期较早,生育期短。吴川等[24]对中国辽宁省和江苏省的杂草稻植物性性状进行了对比分析,认为在生殖生长期的果皮色、谷粒长宽比、百粒重、落粒性、秆硬度、剑叶宽、剑叶长、有效穗数等性状与相应的当地栽培稻存在差异。由此可见,与栽培稻相比,杂草稻前期具有明显生长优势,与栽培稻争夺养分,这也是它在田间影响栽培稻生长的最主要原因。
2 杂草稻的分类
亚洲栽培稻可分为籼粳两个亚种已是研究者的广泛共识。研究发现,杂草稻也存在籼粳分化[25-26]。Tang等[27]对来源于不同国家的24份杂草稻材料同栽培稻和野生稻一起,在落粒性、种子休眠性、千粒重等方面进行比较研究,将其划分为3种类型,即拟籼型,籼型自生型和粳型自生型。陈晓锋等[28]研究发现,江苏省杂草稻主要以籼型和偏籼型为主,其中极少数为粳型。刘丹等[29]利用SRAP分子标记结合程氏指数判定法认为中国杂草稻呈南籼北粳的分布状态。研究发现,一些籼粳特异性分子标记如SSILP和InDel标记被认为可以高效鉴定杂草稻的籼粳分化[30]。此外,杂草稻细胞质也存在籼粳亚种的分化,Sun等[31]研究表明,中国东北杂草稻在ORF100和ORF29两个细胞质位点上基因型均为粳稻类型。杨杰等[32]对江淮流域杂草稻叶绿体DNA的籼粳分化进行了研究,发现江淮流域杂草稻的细胞质来源既有粳稻类型,又有籼稻类型。这些研究结果将为进一步阐明杂草稻的分子进化机理和杂草稻的来源奠定基础。
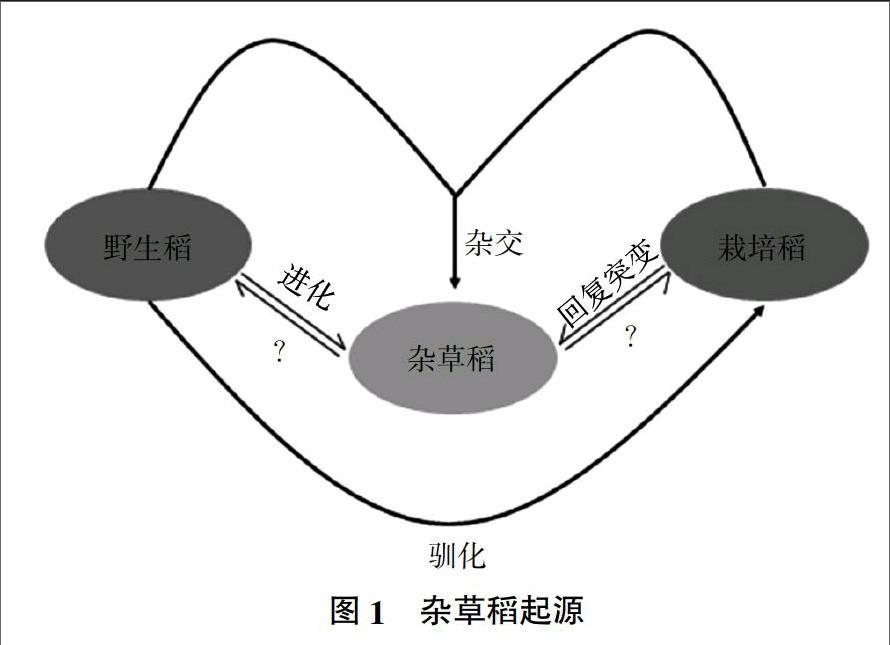
3 杂草稻的起源
杂草稻的起源一直是研究者关注的焦点,相关研究结果不仅有助于杂草稻的有效防治,对于阐明和丰富水稻驯化的历史也具有重要意义。Cho等[14]认为杂草稻是栽培稻与野生稻或籼稻与粳稻自然串粉野化而来的一类特殊材料。Heu等[33]的研究认为韩国杂草稻可能来源于粳稻间的回复突变。Reagon等[34]研究表明,美国杂草稻可能是由栽培稻演化而来。Bres-Patry等[35]利用分子生物技术手段对法国杂草稻起源分类进行了研究,结果表明,法国杂草稻与栽培稻的亲缘关系较近,很可能起源于籼粳栽培稻品种间的远缘杂交。马殿荣等[20]研究也表明辽宁杂草稻与当地粳型栽培稻血缘关系很近,与籼稻和野生稻的遗传关系较远,辽宁省杂草稻的起源很可能与当地长期的籼粳稻杂交育种和地理远缘杂交育种有关。Londo等[36]认为美国杂草稻中,一些是野生稻,另一些来源于栽培稻的变异,还有一些来源于野生稻与栽培稻的杂交后代,并认为美国的杂草稻起源于亚洲。
目前来看,有关杂草稻起源主要有3种假说(图1)。①由野生稻进化而来:经进化后具备栽培稻的部分特性;②栽培稻的回复性突变:栽培稻遗落于田间的种子,经演化具备杂草特性;③栽培稻与野生稻的杂交:栽培稻与野生稻经1次或多次杂交后代演化而来。
4 杂草稻的危害与防治
4.1 杂草稻的危害
近年来,杂草稻已经成为世界上公认的稻田恶性杂草[2]。随着耕作制度的变化和轻简栽培新技术的推广,杂草稻的发生日益严重,它不仅影响着栽培稻的产量,对稻米的等级、品质与商品价值也有着巨大的影响。相关研究表明,杂草稻平均密度为2株/m2便会影响栽培稻的产量[37],5株/m2可以使栽培稻减产22%[38],10~20株/m2可导致栽培稻产量降低50%[39],35~40株/m2可使高秆栽培稻的产量减产60%[40]。进一步研究发现,随着杂草稻密度的逐渐增加,栽培稻分蘖数、叶面积指数、叶绿素含量和干物质积累均有不同程度的减少,且水稻株高、有效穗数、穗长、一次枝梗数、穗粒数、千粒重、结实率、实际产量等也均随杂草稻密度的增加而呈现下降趋势[41-43]。杂草稻在与栽培稻竞争养分、水分和光照以及其他生长资源上占有较大优势,是造成栽培稻减产的主要原因。
杂草稻存在对栽培稻米质的影响。一方面是通过混收对稻米品质纯度造成影响,杂草稻红褐色果皮及在稻米加工过程中容易破碎从而降低了稻米的商品等级。另一方面,杂草稻影响栽培稻碾磨品质和营养品质,使得食味品质稍有下降,而外观品质变化不明显。杂草稻对栽培稻群体微生态环境的负面影响,可能是导致栽培稻产量和品质降低的间接因素[41]。
此外,杂草稻还有一个潜在危害即抗性基因的漂移。由于雜草稻的柱头外露率极高,较栽培稻易于发生串粉和基因漂流现象,一旦转基因水稻中转抗除草剂基因、抗虫基因等向杂草稻中漂移,便会使杂草稻秧苗出现抗性,有可能增强杂草稻的竞争优势,增加潜在危害,给水稻生产造成更大危害。
4.2 杂草稻的防治
由于杂草稻与栽培稻的亲缘关系极近,在形态结构和生理特性上具有许多相同的特性,因此,很难在不损害栽培稻的前提下通过某种单一的途径进行有效控制。基于杂草稻的抗逆性较强,且苗期与栽培稻很难区分的特点,张峥等[21]建议在杂草稻与栽培稻出现明显形态学差异的时期,如杂草稻的分蘖初期、扬花期时进行人工拔除是杂草稻防治的有效手段。此外,由于杂草稻的越冬能力较强,可以通过落粒进行自身繁衍,但是如果冬季进行深翻土地,能有效减少杂草稻的发生。同时,对已有杂草稻危害的稻田,稻田播种前一般进行土壤灌水等处理,促使土壤中杂草稻种子提早萌发,再用非选择性除草剂灭杀也是一种行之有效的方法。
通过转基因技术培育抗除草剂水稻,也为杂草稻以及顽固杂草的防除带来了灵活性,能有效控制杂草稻和其他杂草,并可以减少使用传统除草剂及其替代品转而使用对环境较安全的化合物,但其潜在的危害是抗性基因可能从抗性品种流向杂草稻及其他杂草,因此应该充分考虑对未来环境的潜在危害[44]。
5 杂草稻有利性状基因的利用进展
杂草稻由于长期处于恶劣的野生状态中,经受各种灾害和不良环境的选择,使得其具有不少优良性状,如抗病、耐寒、耐旱、耐盐碱等。利用杂草稻的这些有利性状,可为水稻遗传改良和抗性育种提供新的思路。至今为止,前人已经开展了相关的研究。
丁国华等[11,45]采用盆栽试验,在人工设置干旱胁迫条件下,对不同地区杂草稻幼苗期的耐旱性进行了鉴定。发现不同的杂草稻之间抗旱性不同,大部分杂草稻在苗期具有较强的抗旱能力,少数杂草稻在苗期具有极强的抗旱能力。陈惠哲等[5]对丹东杂草稻种子在抗冻和低温发芽方面进行研究,发现杂草稻与常规栽培稻相比具有较明显的差异,丹东杂草稻种子在遗传基础上有自身的特性能适应较严重的冻害。邹德堂[46]研究认为黑龙江杂草稻比栽培稻更耐低温。高男等[47]研究表明,北方杂草稻具有较强的耐低温发芽特性,参试19份材料中,5份杂草稻材料耐冷性为1级,耐冷性强。袁晓丹等[48]试验发现在室外播种过冬后杂草稻有较高的发芽率。
研究还发现杂草稻具有较强的耐盐碱能力。赵娜等[10]选用在辽宁省各地收集到的47份杂草稻为材料,用不同浓度的NaCl溶液处理,对辽宁省杂草稻发芽期的耐盐性进行了初步评价。结果表明,不同浓度NaCl溶液处理对杂草稻芽长影响较小。余柳青等[9]通过对辽宁丹东杂草稻的抗逆性进行研究,发现落粒粳可以忍耐0.5%的盐碱浓度。张丽丽等[49]研究表明,杂草稻WR 03-12的吸胀种子在各浓度盐碱胁迫下发芽率和发芽势受影响较小。
此外,杂草稻具有极强的耐老化能力。李茂柏等[50]通过对13份上海地区收集的杂草稻的耐储藏特性进行分析,发现所有杂草稻比栽培稻更耐老化处理,其中2份杂草稻种子在老化处理40 d后仍保持一定的发芽率,为相当优良的耐储藏特性材料,暗示杂草稻是一种良好的耐储藏特性改良资源,有较好的育种利用价值。
6 展望
杂草稻作为稻田杂草种系严重危害着水稻的产量和品质,开展杂草稻的综合防治工作依旧是今后工作的重点。同时,由于杂草稻自身变异类型丰富,利用杂草稻特异稻属资源将有助于丰富现有栽培稻品种的遗传基础。杂草稻具有极强的抗逆境能力,也是栽培稻抗性育种中重要的抗性基因来源。因此,进一步收集和整理杂草稻野生资源,建立杂草稻野生资源圃,挖掘杂草稻资源中的优良基因尤其是对抗旱、耐寒以及耐盐碱等抗逆性状调控基因的定位和克隆,将为开展栽培稻的抗性遗传改良提供基础“元件”。此外,结合野生稻和栽培稻的驯化过程,分析杂草稻的起源,阐明野生稻、栽培稻与杂草稻之间的亲缘关系也将是杂草稻研究中亟待解决的问题。
参考文献:
[1] FERRERO A,FINASSI A.Viability and soil distribution of red rice(Oryza sativa L. var. sylvatica) seeds[J].In Med.Fac. Landbouw,Rijksunv. Gent,1995:205-211.
[2] BAKI B B,CHIN D V,MORTIMER M.Wild and weedy rice in rice ecosystems in Asia-A review[J].Internationl Rice Research Institute,2000:118.
[3] ARRIETA-ESPINOZA G,SANCHEZ E,VARGAS S,et al. The weedy rice complex in Costa Rica.I. morphological study of relationship between commercial rice varieties,wild Oryza relatives and weedy types[J].Genetic Resources and Crop Evolution,2005,52(5):575-587.
[4] 孟 英,魏永海,栾浩文,等.寒地穞生稻发生原因及防御对策[J].黑龙江农业科学,2005(2):55-56.
[5] 陈惠哲,玄松南,王渭霞,等.丹东杂草稻种子的耐冻能力和低温发芽特性研究[J].中国水稻科学,2004,18(2):109-112.
[6] 苗 微,王国骄,马殿荣,等.辽宁省杂草稻幼苗对低温胁迫的生理响应[J].中国水稻科学,2011,25(6):639-644.
[7] 許 聪,吴万春.海南岛杂草稻的生态考察和鉴定[J].中国水稻科学,1996,10(4):247-249.
[8] 张忠林,谭学林,邓安凤.杂草稻种质资源的综合评价[J].植物遗传资源科学,2002,3(4):47-50.
[9] 余柳青,MORTIMER A M,玄松南,等.杂草稻落粒粳的抗逆境特性研究[J].应用生态学报,2005,16(4):717-720.
[10] 赵 娜,马殿荣,陈温福.北方杂草稻发芽期耐盐性的初步评价[J].中国稻米,2007(3):20-24.
[11] 丁国华,马殿荣,马 巍,等.杂草稻幼苗期耐旱性的初步筛选与评价[J].北方水稻,2009,40(1):11-14.
[12] 张丽丽,孙 健,马殿荣.耐盐杂草稻3个锌指蛋白基因家族的实时定量分析[J].植物生理学通讯,2010(6):529-536.
[13] VINOD K S,NILDA R B,MARITES A S,et al. Polymorphisms in the ALS gene of weedy rice (Oryza sativa L.) accessions with differential tolerance to imazethapyr[J].Crop Prot,2010,29:336-341.
[14] CHO Y C,CHUNG T Y,PARK Y H,et al. Genetic polymorphisms and phylogenetic relationships of Korean red riee (weedy rice in Oryza sativa L.) based on randomly amplified polymorphic DNA (RAPD) makers[J].Korean J Breed,1995,27:86-93.
[15] KWON S L,SMITH R J,TALBERT R E. Comparative growth and development of red rice(Oryza sativa)and riee(O.sativa)[J].Weed Sci,1992,40:57-62.
[16] 张忠林,彭桂峰,田卫东,等.杂草稻种质资源农艺性状的主成分及聚类分析[J].西南农业学报,2004,17(S1):236-240.
[17] GEALY D R,BOCKELMAN H. Southern U.S. weedy rice(Oryza sativa) accessions for entry into the national small grains collection[A].5th International Weed Science Congress Abstract[C].USA:International Weed Science Society,2008.111.
[18] 杨 琳,戴伟民,强 胜,等.杂草稻穗部形态及休眠特性的初步研究[J].江苏农业科学,2009,37(4):121-123.
[19] 孙建昌,王学龙,马 静,等.宁夏杂草稻农艺性状的鉴定评价[J].作物杂志,2014(3):129-132.
[20] 马殿荣,李茂柏,孙 健,等.辽宁省杂草稻生物学多样性及其遗传分化研究[J].沈阳农业大学学报,2008,39(3):265-269.
[21] 张 峥,戴伟民,章超斌,等.江苏沿江地区杂草稻的生物学特性及危害调查[J].中国农业科学,2012,45(14):2856-2866.
[22] FERRERO A. Biology and control of red (Oryza sativa L.var.sylvatica) infesting European rice fields[J].Medoryzae,2001,9:2-4.
[23] 袁晓丹,赵国臣,柳参奎,等.东北地区杂草稻主要农艺性状的评价[J].吉林农业科学,2006,31(6):6-9.
[24] 吴 川,戴伟民,宋小玲,等.辽宁和江苏两省杂草稻植物性状多样性[J].生物多样性,2010,18(1):29-36.
[25] SUH H S,SATO Y I,MORISHIMA H. Genetic characterization of weedy rice(Oryza sativa L.)based on morpho-physiology,isozymes and RAPD makers[J].Theoretical and Applied Genetics,1997,94:316-321.
[26] CHEN L J, LEE D S, SONG Z P. Gene flow from cultivated rice(Oryza sativa)to its weedy and wild relatives[J].Annals of Botany,2004,93(1):67-73.
[27] TANG L H,MORISHIMA H. Genetic characterization of weedy rice and the inference on their origins[J].Breeding Science,1997,7(2):153-160.
[28] 陈晓锋,强 胜,杨金玲,等.江苏省杂草稻的传播与籼粳分化研究[J].中国水稻科学,2015,29(1):82-90.
[29] 刘 丹,孙 健,马殿荣,等.利用SRAP分子标记分析47份杂草稻样品遗传多样性[J].中国水稻科学,2012,26(1):70-76.
[30] 徐 群,许红云,魏兴华,等.基于SSILP、InDel和SSR标记的杂草稻籼粳分类[J].中国水稻科学,2012,26(6):686-692.
[31] SUN J,QIAN Q,MA D R,et al. Introgression and selection shaping the genome and adaptive loci of weedy rice in northern China[J].New Phytologist,2013,197:290-299.
[32] 杨 杰,王 军,曹 卿,等.江淮流域杂草稻叶绿体DNA的籼粳分化[J].中国水稻科学,2009,23(4):391-397.
[33] HEU M H,CHO Y C,SUH H S. Cross affinity of Korean weedy rice to the cultivars Korean[J].J Crop Sci,1990,35:233-238.
[34] REAGON M,THURBER C S,GROSS B L,et al. Genomic patterns of nucleotide diversity in divergent populations of US weedy rice[J]. BMC Evolutionary Biology,2010,10:180.
[35] BRES-PATRY C,LORIEUX M,CLMENT G,et al.Heredity and genetic mapping of domestication domestication-related traits in a temperate japonica weedy rice[J].Theor Appl Genet,2001, 102:118-126.
[36] LONDO J P,SCHAAL B A.Origins and population genetics of weedy red rice in the USA[J].Mol Ecol,2007,16(21):4523-4535.
[37] KWON S L,SMITH R J,TALBERT R E. Interference of red rice (Oryza sativa) densities in rice (O. sativa)[J].Weed Science,1991,39(2):169-174.
[38] DIARRA A,SMITH R J,TALBERT R E.Interference of red rice (Oryza sativa L.) with rice (O. sativa)[J].Weed Science,1985,33(5):644-649.
[39] 杨 庆,马殿荣,宋冬明,等.不同密度杂草稻对栽培稻群体形态特征及产量的影响[J].北方栽培稻,2008,38(5):28-31.
[40] SMITH R J.Weed thresholds in southern US rice(Oryza sativa)[J].Weed Technology,1988,2(3):232-241.
[41] 宋冬明,马殿荣,杨 庆,等.杂草稻对栽培粳稻产量和品质及群体微生态环境的影响[J].作物学报,2009,35(5):914-920.
[42] 宋冬明,马殿荣,杨 庆,等.杂草稻密度对栽培稻生长发育和产量的影响[J].沈阳农业大学学报,2008,39(3):270-273.
[43] 温广月,沈国辉,钱振官,等.杂草稻对水稻生长及产量的影响[J].杂草科学,2011,29(2):51-53,59.
[44] 杨 巍,吴其褒,陈惠哲,等.杂草稻生物学特性、危害与防治方法[J].中国稻米,2008(3):50-53.
[45] 丁国华,马殿荣,刘晓亮,等.中国北方杂草稻幼苗对干旱胁迫的生理响应[J].干旱地区农业研究,2013,31(6):127-132.
[46] 邹德堂.黑龍江省杂草稻的特征特性及耐冷性分析[J].农业现代化研究,2008,29(2):235-238.
[47] 高 男,马殿荣,陈温福.北方杂草稻耐低温发芽特性的初步研究[J].中国稻米,2007(3):5-8.
[48] 袁晓丹,王 亮,王丽丽,等.东北地区杂草稻的抗逆性及遗传特性[J].延边大学农学学报,2006,28(4):12-15.
[49] 张丽丽,马殿荣,陈温福,等.杂草稻种子芽期对盐碱胁迫的响应[J].种子,2011,30(8):13-16.
[50] 李茂柏,王 慧,朴钟泽,等.杂草稻人工老化和耐储藏特性的初步研究[J].作物杂志,2010(5):30-33.
猜你喜欢
现代交际(2016年20期)2017-02-15
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
现代园艺(2016年3期)2016-03-24
中国果菜(2015年2期)2015-03-11
农业科技与装备(2014年11期)2015-02-02
