
三峡水库蓄水过程中神农溪库湾营养盐的动态分布
2017-03-21 07:21:43宋林旭纪道斌刘德富
中国农村水利水电 2017年3期
李 欣,宋林旭,纪道斌,刘德富
(1.三峡大学水利与环境学院,湖北 宜昌 443002;2.三峡库区生态环境教育部工程研究中心(三峡大学),湖北 宜昌 443002;3.湖北工业大学河湖生态修复与藻类利用湖北省重点实验室,武汉 430068)
氮、磷等营养盐是水体富营养化的物质基础[1],且其存在形态,结构及其分布特征与水华生消密切相关。三峡水库蓄水之后,水动力条件发生改变,在不同水位运行期,库区支流库湾普遍存在不同形式干流倒灌现象[2]。倒灌异重流对支流库湾营养盐的贡献有可能成为补给支流营养盐的主要来源[3-6]。为研究干流倒灌对库湾营养盐的影响,本文选择神农溪库湾为研究对象,在蓄水期9-11月进行水动力及营养盐持续监测,研究蓄水过程中支流库湾水动力特性、营养盐变化过程,分析了水动力过程对库湾营养盐动态分布影响,并根据营养盐结构变化特征,评估蓄水过程中水华暴发风险。以期为研究三峡水库运行对库湾水体营养盐动态变化过程提供了基础。
1 材料与方法
1.1 神农溪概况
神农溪系长江三峡水库湖北省库区第二大支流,发源于神农架山脉南坡,地处鄂西北山区。其干流全长60.6 km,于西瀼口注入长江,流域总面积1 047 km2。三峡水库修建前,神农溪早期的污染源主要是人类的生活的污水以及农业生产过程中产生的污染物,且环境负荷较小,因此“水华”现象较为少见。三峡大坝修建后,受蓄水影响,神农溪从河流态变成回水库湾,175 m时形成离河口26 km回水区,呈现出内陆湖泊水文状况。河口至上游段面处水面基本上未有明显流速,水体流动稀释作用大大减弱,自我净化功能衰退,造成营养物质停滞于固定区域,给营养物质的浓度累积形成了条件,神农溪库湾近年来多次暴发水华。
1.2 样点布设
根据神农溪水流特点及河流形态,在神农溪库湾回水区内设置SN01~SN06共6个断面,另设置SNYT为上游入流控制断面,在长江干流中设置CJBD断面(见图1和表1)。
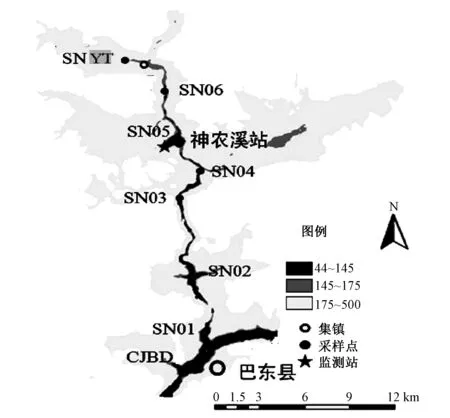
图1 监测断面布设示意图Fig. 1 Map of the monitoring section layout diagram
1.3 监测时段及采样时间
监测时段:2013年汛末蓄水期(9月10日-11月24日);根据三峡工程2013年175 m试验性蓄水计划安排的时间和进程将采样次数定为3次,分别是9月17日、10月20日和11月21日。

表1 监测断面设置说明Tab.1 The monitoring section setup instructions
1.4 监测指标及分析方法
监测指标如表2所示,包括水位、水深、水温、流速以及营养盐指标中的总氮(TN)、总磷(TP)、正磷酸盐(PO4-P)、硝氮(NO3-N)、氨氮(NH4-N)和溶解性硅酸盐(D-Si)。水深、水温和流速这三个指标现场监测后在电脑中导出数据并分析,营养盐指标参考《水和废水分析方法》(第四版)[7]和《湖泊生态调查观测与分析》[8]测定,在样品采集后的3 d内完成分析。
2 结果与分析
2.1 水动力特性
2.1.1 水位变化特点
监测期间水位变化如图2所示。根据三峡水库蓄水前后水位日变幅及变化趋势分为3个阶段:
第一阶段——9月10日至9月22日,历时13 d,水位从157.97 m变化至166.4 m,共计上升8.43 m,日均升幅0.648 m,日变幅达到最大值1.68 m,水位上升较快;
第二阶段——9月23日至11月10日,历时49 d,水位从166.47 m变化至174.89 m,共计上升8.42 m,日均升幅0.172 m,水位上升较慢;
第三阶段——11月11日至11月24日,历时14 d,此阶段水位在174.58 m至174.99 m之间波动,变化幅度极小。

图2 蓄水期神农溪库湾水位变化Fig.2 Shennong Bay water level changes during the period of Impoundme

监测指标监测仪器预处理监测方法水位-- 从中国长江三峡集团公司网站(http:∥www.ctg.com.cn/inc/sqsk.php)直接获取水深美国产HydroLabDS5多参数水质监测仪- 现场测量,首先在水面停留35s,然后沿垂向缓慢向下放仪器,读数每1s更新一次水温美国产HydroLabDS5多参数水质监测仪- 现场测量,首先在水面停留35s,然后沿垂向缓慢向下放仪器,读数每1s更新一次流速挪威产“威龙”声学多普勒三维点式流速仪6MHzVectorADV-现场测量,沿垂向每2m监测一组数据,可监测采样点沿E、N及沿水深方向的流速大小总氮(TN)紫外分光光度计 将采集的300mL水样用GF/C滤膜过滤(孔径0.45μm)并低温保存 将预处理过的水样带回实验室进行水化学分析总磷(TP)紫外分光光度计 将采集的300mL水样用GF/C滤膜过滤(孔径0.45μm)并低温保存 将预处理过的水样带回实验室进行水化学分析正磷酸盐(PO4-P)紫外分光光度计 将采集的另一瓶300mL水样加入0.15mL浓H2SO4低温保存 将预处理过的水样带回实验室进行水化学分析硝氮(NO3-N)紫外分光光度计 将采集的另一瓶300mL水样加入0.15mL浓H2SO4低温保存 将预处理过的水样带回实验室进行水化学分析氨氮(NH4-N)紫外分光光度计 将采集的另一瓶300mL水样加入0.15mL浓H2SO4低温保存 将预处理过的水样带回实验室进行水化学分析溶解性硅酸盐(D-Si)紫外分光光度计 将采集的另一瓶300mL水样加入0.15mL浓H2SO4低温保存 将预处理过的水样带回实验室进行水化学分析
2.1.2 水流特点
图3为2013年蓄水过程中SN01断面不同阶段流速沿垂向分布规律。第一阶段中,干流水体以表层倒灌异重流形式潜入神农溪库湾。根据监测数据,干流水体从表层至水深38 m范围潜入库湾,水深38 m以下的库湾水体从上游流向河口,最大流速达到0.167 m/s,平均流速为0.045 5 m/s[图3(a)];第二阶段干流水体以中层倒灌异重流形式潜入库湾,干流水体从中层(水深10~35 m范围)潜入库湾,水深约35 m以下和10 m以上的水体从库湾流向河口,平均流速达到0.024 m/s[图3(b)];在第三阶段中,干流水体以底部倒灌异重流形式潜入库湾,11月21日干流水体从66m至河底范围潜入库湾,最大流速达到0.147 m/s,平均流速达到0.044 m/s[图3(c)]。蓄水过程中库湾是具有明显的分层异向流动特性,产生这种分层异向流动的最主要原因是由干支流水体密度差引起的[10]。
图4显示了各蓄水阶段的神农溪库湾水流特性,其中9月17日、10月20日分别为蓄水较快和较缓时段的水流特性。可以看出,蓄水过程中支流水流具有较明显的分层异向流动特性,整个蓄水过程中神农溪库湾流速整体小于0.1 m/s。第一阶段中,在距河口18 km以内的水域水体整体呈分层异向流特性,水库干流水体由表层至水深38 m范围潜入支流,水深30 m以下的支流水体流向干流,从距河口18km以上至回水末端水域,水体由上游流向下游,呈一维特性[图4(a)]。第二阶段水位日升幅缓慢,在距河口7 km以内的水域水体整体呈分层异向流特性,干流水体由水深10~35 m范围倒灌入支流,水深约35 m以下和10 m以上的支流水体流向干流(图4(b)),倒灌异重流方式由表层倒灌变为中上层倒灌,倒灌强度与潜入范围较第一阶段明显减小,这是因为在蓄水开始后,气温逐渐下降,神农溪库湾表层水体温度迅速降低,而干流水温降低较慢,干流水体从表层灌入神农溪库湾形成表层异重流。而随着蓄水过程的进行,干流水温逐渐降低,而神农溪库湾表层水体温度变化不如第一阶段的变化明显,因此,干流水体倒灌入支流的潜入点下降。
2.1.3 水温变化特点
在蓄水过程中,各个阶段表层水温差异明显(图5),第一阶段支流水温比干流水温高3.27 ℃,主要是因为干流流速大,水体垂向混合较强而不易形成温度分层,支流流速小,水温分层明显,从河口到上游各断面的水温逐渐升高;在第二阶段中,水库干支流的水温差迅速减小,各断面间水温波动较小;而蓄水结束后,干支流的温度差趋于0。整个蓄水过程中,水库干支流水温整体呈下降趋势,这主要是受气温的影响,但是在整个蓄水过程中水温均大于20 ℃,满足藻类生长的温度条件[9]。
2.2 营养盐沿水流方向的时空分布特征
2.2.1 磷的纵向时空分布特征
在蓄水过程中,神农溪库湾的TP和PO4-P纵向时空分布如图6示。蓄水第一阶段由图6(a) 可知,干流CJBD断面的TP浓度略高于库湾各断面平均值,略低于河口SN01断面,从SN01至SN04断面呈快速递减趋势,而上游区域的变化趋势则与之相反,TP浓度呈逐渐升高的趋势;蓄水第二阶段中,干流CJBD断面的TP浓度略低于河口附近SN01断面,从SN01至SNYT呈缓慢递减趋势;蓄水第三阶段干流CJBD断面至SN03断面处TP浓度无明显变化,而SN04处TP浓度达到最小值,上游区域TP浓度逐渐升高。

图3 SN01断面流速沿深度方向分布Fig.3 SN01 sectional velocity distribution along the depth direction注:水流流入库湾流速为正,流出库湾为负

图4 流速分布纵剖面图Fig. 4 Profile of velocity distribution profile注:图中白色箭头表示由河口流向支流,黑色箭头表示由支流流向河口,箭头长度表示流速大小(m/s)

图5 蓄水期神农溪库湾水温变化Fig.5 Shennong Bay water temperature changes during the period of Impoundment
由图6(b) 可知,蓄水第一阶段干流CJBD断面的PO4-P浓度明显高于库湾各断面,从SN01断面至SNYT断面PO4-P浓度整体上逐渐降低;第二阶段中,干流CJBD断面的PO4-P浓度明显低于下游SN01与SN02断面,而从SN01至SN06断面,PO4-P浓度呈现逐渐降低的趋势,而上游SNYT断面的PO4-P浓度则高于库湾平均值;到了第三阶段,从干流CJBD断面到SN03断面的TP浓度无明显差异,在SN04处PO4-P浓度达到最小值,从SN04断面到上游区域呈线性递增趋势。
蓄水末期TP平均浓度比初期略有降低,由0.056 mg/L降至0.049 mg/L;而PO4-P则由0.011 mg/L上升至0.026 mg/L,PO4-P占TP百分比从19.6%升高至约53%,吸附态P的浓度从蓄水初期的0.045 mg/L降至0.023 mg/L,这是由于在蓄水过程中水流减缓,水体中碎屑物质迅速沉降[11],被悬浮泥沙吸附的颗粒态磷酸盐沉降[12-14]。
2.2.2 氮的纵向时空分布特征
图7为神农溪库湾蓄水期间TN与NO3-N动态分布。由图7(a)可知,在蓄水第一阶段,从干流CJBD断面到SN06断面,TN浓度呈逐渐降低的趋势,上游SNYT断面的TN浓度远远低于库湾平均值;到了蓄水第二阶段,干流CJBD断面的TN浓度略低于河口SN01断面,略高于库湾平均值,而从SN01到SN06断面TN浓度上下波动,无明显变化趋势,上游SNYT断面的TN浓度还是远远低于库湾平均值;在蓄水第三阶段中,干流CJBD断面与河口SN01断面的TN浓度大致相等,而从SN01到SN04断面TN浓度缓慢降低,在上游区域的TN浓度趋于一致。

图6 蓄水期间神农溪库湾的TP和PO4-P纵向时空分布Fig. 6 TP and PO4-P longitudinal time and space distribution in Shennong Bay during the period of impounding

图7 蓄水期间神农溪库湾的TN和NO3-N纵向时空分布Fig.7 TN and NO3-N longitudinal time and space distribution in Shennong Bay during the period of impounding
由图7(b)可知,在蓄水第一阶段中,从干流CJBD断面到库湾下游区域NO3-N浓度逐渐降低,上游区域的NO3-N浓度趋于一致;库湾的NO3-N浓度在蓄水第二阶段中无明显变化规律,上游SNYT断面远远低于库湾平均值;而到了蓄水第三阶段,NO3-N浓度则无明显变化规律,整体上相当。
TN与NO3-N的变化范围分别为0.924~2.569和0.608~1.796 mg/L,均值分别是1.759 mg/L和1.341 mg/L。蓄水末期与初期比较,库湾中总氮含量由1.302 mg/L上升到1.77 mg/L,NO3-N浓度也由0.843 mg/L上升至1.563 mg/L,NO3-N占TN百分比从65%升高至约88%。蓄水使NO3-N比例增加。而NH4-N对环境变化敏感,受此地区人为活动干扰大[15]。蓄水期间变化没有明显差异,调查期间NH4-N的变化范围0.002~0.295 mg/L,平均值0.143 mg/L。
2.2.3 硅的纵向时空分布特征
神农溪库湾D-Si动态分布如图8所示,由图可知,蓄水第一阶段各断面的D-Si浓度均大于第二阶段和第三阶段,而第二阶段和第三阶段各断面间D-Si浓度相近且空间差异较小,蓄水末期的D-Si浓度较蓄水初期下降了约1/3。蓄水第一阶段到第二阶段中,库湾D-Si浓度变化较大,这是由于上游来流量减少,水流流速减缓,水体中碎屑物质沉积使水体中溶解性硅酸盐的补给量减少,同时可溶性硅酸盐自身的脱水作用[16]和生物沉积作用[17]使水体中的D-Si浓度迅速降低。

图8 蓄水期间神农溪库湾的D-Si纵向时空分布Fig.8 D-Si longitudinal time and space distribution in Shennong Bay during the period of impounding
2.2.4 叶绿素的时空分布特征
在整个蓄水过程中神农溪库湾叶绿素a浓度变化显著,干流CJBD断面与上游控制断面SNYT的叶绿素a浓度基本保持不变,在4 mg/m3以下(图9),峰值出现在蓄水第一阶段的上游SN05水域,且蓄水第一阶段的上游区域和蓄水第二阶段除了河口SN01断面外的其余断面的叶绿素a浓度都超过了“水华”暴发叶绿素a浓度的阀值10 mg/m3,使水体呈中富营养化状态[18]。而到了蓄水第三阶段,神农溪库湾与干流水体的叶绿素a浓度接近。

图9 蓄水期间神农溪库湾的Chl.a时空分布Fig.9 Chl.a longitudinal time and space distribution in Shennong Bay during the period of impounding
2.3 营养盐结构分析
2.3.1 N/P的动态分布特征
N/P对藻类的暴发性生长具有重要意义,是水中浮游植物营养结构特点的重要反映[19]。据此学者提出富营养化评估的营养物限制性划分标准:①当水中N/P<7-10时,藻类生长表现为氮限制状态,生物固氮作用有可能发生以调节N/P,消纳水体中相对较多的磷;②N/P>22.6~30时,磷将成为藻类生长的限制性因子[20,21],较低的磷含量水平可能使得氮素的有机合成过程受到抑制,无机氮向有机氮形态转化速率降低;③N/P介于两者之间时为藻类生长的合适范围,通过对硝氮的利用和摄取以实现对氮素有机合成的生态过程较为明显。
由图10所示,蓄水期间神农溪库湾N/P空间差异很大,蓄水第一阶段从河口到SN04断面N/P逐渐升高,而从SN04到上游SN07断面的N/P则逐渐降低,干流处N/P略高于河口;第二阶段从SN01到SN06断面N/P总体上是缓慢增加的,上游SN07断面的N/P低于库湾平均值,干流的N/P同样略高于河口;到了第三阶段,从干流断面到SN06断面,N/P总体上呈逐渐升高的变化趋势,而上游SN07断面N/P明显低于SN06断面。在整个蓄水过程中,N/P的最大值出现在蓄水第三阶段的SN06断面为57.406,最小值出现在蓄水第一阶段的河口SN01断面为12.75。根据Justic提出的营养盐浓度限制法则,有57%的N/P大于22,43%的介于10和22之间。

图10 蓄水期间神农溪库湾表层水体N/P动态分布Fig.10 Surface layer waters N/P dynamic distribution in Shennong Bay during the period of impounding
2.3.2 Si/N的动态分布特征
图11所示蓄水期间神农溪库湾表层水体Si/N动态分布,从时间上看,在蓄水第一阶段中,从下游到上游Si/N在逐渐增大,最大值出现在上游SN07断面为8.14;到了蓄水的第二阶段和第三阶段,从干流到SN06断面的Si/N整体上相当,无明显的变化规律。从空间分布来看,蓄水初期干流水体Si/N小于神农溪,到了蓄水末期干支流Si/N总体上相当,并且库湾各断面的Si/N在蓄水末期都比初期有了显著的降低。总的来说,蓄水末期比蓄水初期Si/N减小。有国外学者研究表明,浮游植物对D-Si的吸收比例与N相近,对营养盐的吸收比例是16(Si)∶15(N)∶1(P),当环境中(Si/N)<1时,表明Si为潜在的限制因子[22]。而在此次监测过程中,Si/N的变化范围为1.68~8.14,(Si/N)>1,表明在神农溪蓄水过程中,硅不是藻类生长的限制因子。

图11 蓄水期间神农溪库湾表层水体Si/N动态分布Fig.11 Surface layer waters Si/N dynamic distribution in Shennong Bay during the period of impounding
2.3.3 Si/P的动态分布特征
图12显示的是蓄水期间神农溪库湾表层水体Si/P动态分布,可以看出Si/P的时空分布差异很大,变化范围为23~264。整个蓄水过程中,下游略小于上游,库湾各断面沿水流方向先降低然后升高。
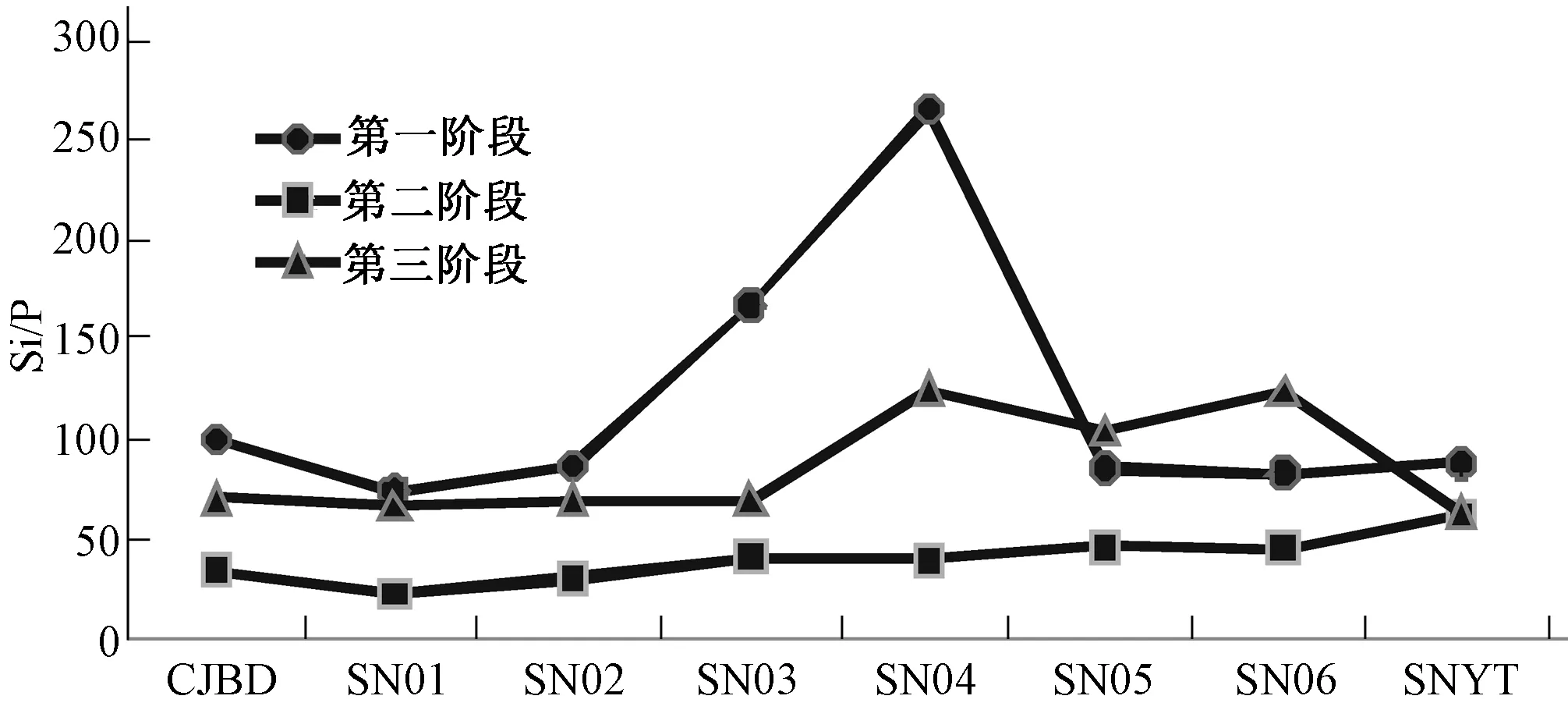
图12 蓄水期间神农溪库湾表层水体Si/P动态分布Fig.12 Surface layer waters Si/P dynamic distribution in Shennong Bay during the period of impounding
3 讨 论
3.1 水动力特性分析及其对营养盐分布的影响
蓄水第一阶段,坝前水位快速升高,干流水体从表层倒灌入支流,根据监测数据可知,蓄水第一阶段在距河口18 km以内的水域水体整体呈分层异向流特性,给整个库湾补给了大量的营养盐,但是在库湾中上游区域,由于水温、光照等环境因子和缓慢的流速适宜浮游植物生长,致使下游营养盐整体上大于中上游。不过上游来流也补给了部分营养盐给库湾上游区域,因此库湾上游区域的营养盐略高于库湾中游区域。第二阶段,坝前水位上升缓慢,干流水体由表层倒灌异重流逐渐过渡至中层倒灌异重流潜入神农溪库湾,在距河口7 km以内的水域水体整体呈分层异向流特性,对库湾中下游区域营养盐补给大于对中上游区域的营养盐补给。在蓄水第三阶段,坝前水位已到达175 m的正常蓄水位。倒灌潜入点继续下移,干流水体从中层倒灌异重流变为底部倒灌异重流,在距河口16 km以内的水域水体整体呈分层异向流特性,整个库湾表层的营养盐浓度大致持平,无明显变化趋势。Schemel[23~24]等认为,水体的各种运动形式造成了生物有机体和生源要素在不同空间尺度的分布,使水体环境发生改变并影响着库区和河流水生生物的组成、数量以及分布。干流倒灌异重流的持续存在,使库湾表层各营养盐指标整体呈现自下游向上游逐渐降低的变化趋势,说明倒灌异重流在给神农溪库湾输入水量的同时,也为库湾补给了大量的营养盐[25]。在整个蓄水过程中,水温始终大于20℃,满足藻类增殖水温条件,因此温度不是影响营养盐分布的主要因素。
3.2 蓄水过程对神农溪库湾水华暴发风险的分析
罗专溪等人[26]通过研究三峡水库大宁河回水河段氮磷的来源、数量及时空特征,指出氮、磷营养盐受干流倒灌影响明显,并提出了干流逆向影响效应,这种干流逆向影响效应可能就是异重流影响效应。而蓄水期间水流水动力环境有很大改变,长江水体泥沙含量较大,由于水库水位的不断抬升,密度较大的水流更容易进入库湾,与库湾水体进行掺混和交换,也就必然会改变营养盐浓度和结构分布特征。国外有学者研究发现[27~28],饱和营养生长的藻细胞中N和P的原子比约为16∶1,当(N/P)>30时,P为限制因子;(N/P)<8时,N为限制因子;而N/P在8~30之间属于正常的波动,15~16是藻类生长最佳水平。Justic等[29~30]提出:可溶性Si=2 μmol/L、N=1 μmol/L、P=0.1 μmol/L为限制浮游植物生长的营养盐浓度阈值;当水体中(Si/P)>22和(N/P)>22时,P是浮游植物生长的营养限制;当(N/P)<10和(Si/N)>1时,为N限制;当(Si/P)<10和(Si/N)<1,Si是限制因子。数据显示,蓄水期间N/P大多集中在8~30之间,N和P都不是限制因子,而在蓄水的第三阶段中,中上游区域的(N/P)>30,P为限制因子。蓄水过程中神农溪库湾(Si/P)>22,(Si/N)>1,可见Si不是神农溪库湾的营养限制。
蓄水过程中,支流水体呈现分层异向流态的特性,干流水体分别从库湾的表层、中层、底层倒灌入库湾,并且水位上升越快,倒灌强度则越大。所以当干支流进行水体交换时,由于干流叶绿素a含量较低,故会对库湾水体的叶绿素a含量起稀释作用。在蓄水第一阶段中,水位迅速提升,干流水体以表层倒灌入库湾,且倒灌强度直至中游,从河口到库湾中游区域的叶绿素a浓度大于干流,小于10 mg/m3,而上游区域叶绿素a浓度明显高于10 mg/m3,达到了中富营养化状态;蓄水第二阶段,干流水体从中层倒灌入库湾,除了河口SN01断面,库湾其余断面叶绿素a含量都高于10 mg/m3,此阶段整个库湾都达到中富营养化状态;在蓄水末期整个库湾叶绿素a含量都低于5 mg/m3,处于贫营养化状态,暴发水华的风险较小。
在蓄水第一阶段中,从干流到库湾中游区域的N/P整体上大于22,而从SN05断面到上游SN07断面的N/P值则介于10~22,说明在蓄水第一阶段中库湾上游区域富营养化比较严重,水华暴发风险较大;到了蓄水第二阶段,从干流到上游断面的N/P几乎都集中在14~20之间,整个库湾水华暴发的风险最大;而在蓄水末期即蓄水第三阶段中,库湾各断面的N/P都大于22,说明蓄水第三阶段富营养化较之前两个阶段有所减弱,不利于水华的暴发。而有学者提出富营养化评估的营养物限制性划分标准中说到当N/P介于10到22之间时为藻类生长的合适范围,因此我们可以认为当氮磷比的值在10~22之间时,水华的暴发风险将会增大。总的来说,在蓄水第一阶段中,库湾上游区域为水华暴发提供了有利的条件,而蓄水第二阶段是整个蓄水阶段库湾最有可能暴发水华的时期,到了蓄水第三阶段则不利于水华的暴发,这与库湾叶绿素a含量时空分布的分析结果一致。
4 结 论
(1)2013年神农溪库湾蓄水过程中,水位最大日升幅为1.68 m,水库干流水体分别以表层、中层和底层异重流的形式倒灌入支流库湾。
(2)蓄水初期,神农溪库湾营养盐分布差异很大;蓄水完成后,神农溪库湾和长江营养盐含量趋于一致。库湾TP含量由0.056 mg/L降至0.049 mg/L;而PO4-P则由0.011 mg/L上升至0.026 mg/L,PO4-P占TP百分比从19.6%升高至约53%;TN含量由最初的1.302 mg/L上升到1.767 mg/L,NO3-N浓度也由0.843 mg/L上升至1.563 mg/L,NO3-N占TN百分比从65%升高至约88%;D-Si浓度相比于蓄水初期下降了约1/3。
(3)营养盐结构方面,蓄水期间神农溪库湾(Si/P)>22,(Si/N)>1,表明Si不是神农溪库湾的营养限制。在蓄水第一阶段,库湾上游区域的(N/P)介于10到22之间,上游区域暴发的风险大于下游;而在蓄水第二阶段,整个库湾的(N/P)几乎都集中在14到20之间,满足藻类生长的最佳条件,水华暴发风险;到了蓄水第三阶段,整个库湾(N/P)>22,此阶段由于磷营养盐限制相较于前两个阶段,水华暴发的风险明显降低。
□
致谢本研究的采样和监测工作由三峡水库支流水质监测重点站神农溪基层站成员协助完成,在此表示感谢。
[1] 李锦秀,廖文根. 三峡库区富营养化主要诱发因子分析[J]. 科技导报,2003, 21(0309): 49-52.
[2] 谢 涛. 三峡水库调度对香溪河库湾水温特性的影响研究[D]. 湖北宜昌:三峡大学,2014.
[3] Holbach A, Wang L, Chen H, et al. Water mass interaction in the confluence zone of the Daning River and the Yangtze River-a driving force for algal growth in the Three Gorges Reservoir[J]. Environmental Science and Pollution Research, 2013,20(10):7 027-7 037.
[4] 陈媛媛,刘德富,杨正健,等. 分层异重流对香溪河库湾主要营养盐补给作用分析[J]. 环境科学学报,2013,33(3):762-770.
[5] 吉小盼,刘德富,黄钰铃,等. 三峡水库泄水期香溪河库湾营养盐动态及干流逆向影响[J]. 环境工程学报,2010,(12):2 687-2 693.
[6] 张 宇,刘德富,纪道斌,等. 干流倒灌异重流对香溪河库湾营养盐的补给作用[J]. 环境科学,2012,33(8):2 621-2 627.
[7] 复 盛,国家环境保护总局,水和废水监测分析方法委员会. 水和废水监测分析方法[M]. 北京:中国环境科学出版社,2002.
[8] 黄祥飞,孙鸿烈,刘光崧. 湖泊生态调查观测与分析[M]. 北京:中国标准出版社,2000.
[9] 黄钰铃,陈明曦,郭 静. 不同水温下蓝藻水华生消模拟与预测[J]. 三峡大学学报: 自然科学版,2009,31(1):84-88.
[10] 纪道斌,刘德富,杨正健,等. 三峡水库香溪河库湾水动力特性分析[J]. 中国科学: G辑,2010,(1):101-112.
[11] 钱 宁,万兆慧. 泥沙运动力学[J]. 2003.
[12] 曹承进,秦延文,郑丙辉,等. 三峡水库主要入库河流磷营养盐特征及其来源分析[J]. 环境科学,2008,29(2):310-315.
[13] 王圣瑞,金相灿,庞燕. 不同营养水平沉积物在不同 pH 下对磷酸盐的等温吸附特征[J]. 环境科学研究,2005,18(1):53-57.
[14] 周济福,曹文洪,杨淑慧,等. 河口泥沙研究的进展[J]. 2003,6:75-83.
[15] 曹 明,蔡庆华,刘瑞秋,等. 三峡水库库首初期蓄水前后理化因子的比较研究[J]. 水生生物学报,2006,30(1):12-19.
[16] 杨东方,高振会,陈 豫,等. 硅的生物地球化学过程的研究动态[J]. 海洋科学,2002,26(3):39-42.
[17] 杨东方,高振会,秦 洁,等. 地球生态系统的营养盐硅补充机制[J]. 海洋科学进展,2006,24(4):568-576.
[18] 郑丙辉, 张 远, 富 国, 等. 三峡水库营养状态评价标准研究[J]. 环境科学学报, 2006,26(6):1 022-1 030.
[19] 李 哲,郭劲松,方 芳,等. 三峡水库小江回水区不同 TN/TP 水平下氮素形态分布和循环特点[J]. 湖泊科学,2009,21(4):509-517.
[20] 彭近新,陈慧君. 水质富营养化与防治[M]. 北京:中国环境科学出版社,1988.
[21] Guildford S J, Hecky R E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship?[J]. Limnology and Oceanography, 2000,45(6):1 213-1 223.
[22] Conley D J, Malone T C. Annual cycle of dissolved silicate in Chesapeake Bay: implications for the production and fate of phytoplankton biomass[J]. Marine ecology progress series, 1992,81(2):121-128.
[23] Schemel L E, Sommer T R, Müller-Solger A B, et al. Hydrologic variability, water chemistry, and phytoplankton biomass in a large floodplain of the Sacramento River[J]. Hydrobiologia, 2004,513(1-3):129-139.
[24] Leitao M, Morata S M, Rodriguez S, et al. The effect of perturbations on phytoplankton assemblages in a deep reservoir (Vouglans, France)[C]∥Phytoplankton and Equilibrium Concept: The Ecology of Steady-State Assemblages. Springer Netherlands, 2003:73-83.
[25] Huang Y L, Zhang P, Liu D F, et al. Nutrient spatial pattern of the upstream, mainstream and tributaries of the Three Gorges Reservoir in China[J]. Environmental monitoring and assessment, 2014,186(10):6 833-6 847.
[26] 罗专溪,朱 波,郑丙辉,等. 三峡水库支流回水河段氮磷负荷与干流的逆向影响[J]. 中国环境科学,2007,27(2):208-212.
[27] Reynolds C S. The ecology of freshwater phytoplankton[M].London:Cambridge University Press, 1984.
[28] 韩新芹,叶 麟,徐耀阳,等. 香溪河库湾春季叶绿素a浓度动态及其影响因子分析[J]. 水生生物学报,2006,30(1):89-94.
[29] Justic D, Rabalais N N, Turner R E, et al. Changes in nutrient structure of river-dominated coastal waters: stoichiometrie nutrient balance and its consequences [J].Estuarine Coastal and Shelf Science, 1995,40:339-356.
[30] 李 军,刘丛强,王仕禄,等. 太湖水体溶解营养盐(N、P、Si)的冬、夏二季变化特征及其与富营养化的关系[J]. 地球与环境,2005,33(1):63-67.
猜你喜欢
中国资源综合利用(2022年5期)2022-06-06 01:52:32
科技与创新(2021年10期)2021-06-01 07:10:24
中国水利(2020年23期)2020-12-24 06:24:40
水利规划与设计(2017年11期)2017-12-23 06:34:19
水利技术监督(2017年6期)2017-12-19 13:28:17
化工管理(2017年9期)2017-03-05 12:05:20
水利科技与经济(2016年3期)2016-04-22 01:04:28
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
湿地科学与管理(2015年3期)2015-12-07 07:57:24
应用海洋学学报(2015年3期)2015-11-22 07:39:10
