
燕子掌电信号随光照度变化的分析
2017-03-21 18:15王晓琳田立国李猛
江苏农业科学 2016年11期
关键词:光合作用
王晓琳+田立国+李猛
摘要:光环境是影响光合作用最重要的因素。不同植物所需的最适宜光照条件不同,通过检测计算燕子掌(Crassula obliqua)电信号的时域和频域特性,研究不同光照度下燕子掌电信号时域峰峰值、均值、频域功率谱的变化,以及表征它们的特征值谱重心频率、谱边缘频率和功率谱熵,探究它们最适宜的光照度。试验证明,燕子掌电信号是一种微弱低频信号,其光合作用的強弱与其植物电信号密切相关,燕子掌最适宜的光照度为24 000~29 500 lx,在此光照度范围其光合作用较强。
关键词:光合作用;植物电信号;燕子掌;时域和频域分析
中图分类号: TP274;S184 文献标志码: A
文章编号:1002-1302(2016)11-0064-03
植物电信号是一种能够在细胞、组织间传递的微弱信号[1],它是植物对环境变化刺激引起的反应[2],电信号激发植物产生运动、生长代谢及物质运输等生理变化,从而调节植物与外部环境的联系[3]。不同环境刺激会使植物产生不同的电位波动,因此可以通过观察植物电信号的变化来检测外部环境因子的改变[4-6],以此营造作物自身最适宜的生长环境,从而为温室调控提供一种可行的方法,为农业现代化的发展奠定基础。
光合作用是一切植物赖以生存的基础[7],光照是植物能量的来源,因此光照度是植物生长发育过程中最重要的环境影响因子之一,光环境对植物生长的各个方面都有着广泛的调节作用。光照的很多因素都会影响植物生长过程中的健康状态以及生长快慢[8],主要包括光照度(光量)、光质(光谱成分)、光周期(明/暗期时间[9])。本研究通过对不同光照条件下燕子掌(Crassula obliqua)电信号进行采集,并对其进行系统精确地分析,以确定光照度对燕子掌生长的具体影响。
1 材料与方法
1.1 测试仪器及材料
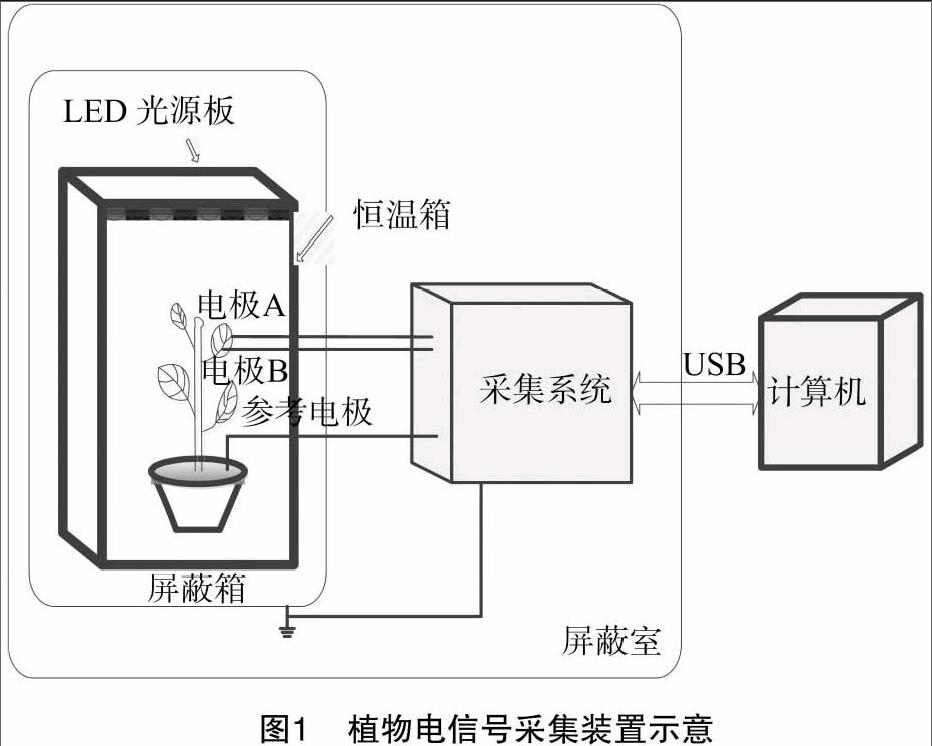
基于NI-DAQ数据采集和LabView的植物电信号检测及采集系统;可控LED灯光板(红蓝配比为8 ∶1,可调节发光亮度);自制铜网屏蔽室(150 cm×50 cm×150 cm);铂金电极(直径0.3 mm);生长状态良好的盆栽燕子掌。试验的环境参数保持为:温度保持在25~28 ℃,花盆土壤湿度为15%~25%,室内空气湿度为40%~50%,手持式光谱计MK350(可检测光照度)。
1.2 方法
将盆栽生长状况良好的燕子掌放入植物生长柜中,置于自制的铜网屏蔽箱内,在植物箱顶放置可控光的LED灯光板,屏蔽箱和屏蔽室外壳接地,铂金电极直接刺入植物叶片组织,正负电极之间的距离约为10 mm,将铂金电极接入植物电采集系统的正负两极,接地电极插入土壤中,信号经过两电极接入植物电信号采集系统,合理设置其参数,而后将转换的数据经过USB传输线输入计算机,采集装置图1所示。
2 时域和频域分析
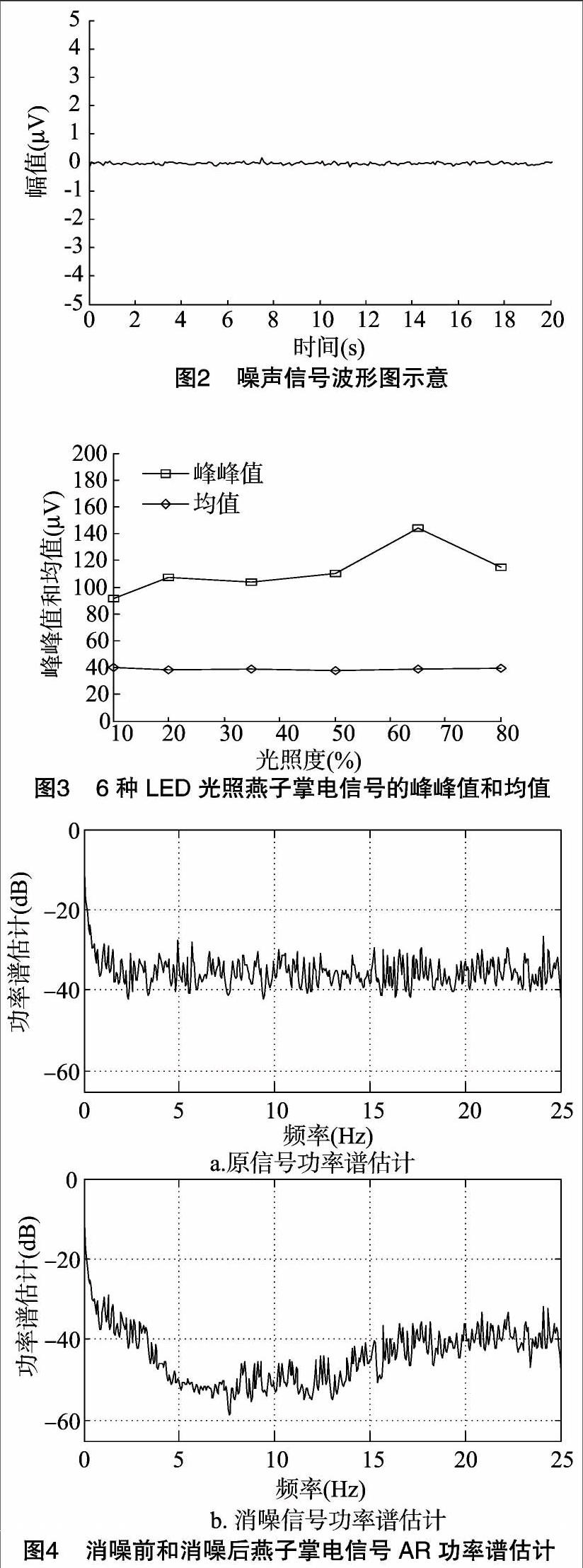
为了更好地说明采集到的电信号,首先让采集系统工作1 h,待工作稳定后先不接任何植物,看采集系统受到外界噪声干扰有多大,从噪声图(图2)可以看出,外界噪声很小,该系统可以很好地满足试验要求。
2.1 燕子掌信号的时域分析
时域分析是最基本的信号分析方法,它可以表现最基本的信号特征,LED灯光板的发光亮度可控(量程为1%~100%),满量程为100%,分别设置10%(8 003 lx)、20%(15 010 lx)、35%(19 230 lx)、50%(24 015 lx)、65%(27 819 lx)、80%(29 430 lx)6种不同的光照环境,他们对应的光照度由光谱计MK350测定,植物生长温度保持在25~28 ℃,相对湿度保持在45%。植物电信号是一种微弱信号,因此受噪声影响极大,如不进行消噪,很有可能使有用信号淹没于噪声中。目前小波软阈值消噪已被广泛应用于脑电、心电、肌电等微弱电信号的研究[10-11],并取得良好的试验效果,所以将采集到的信号先经过小波软阈值消噪,经过Matlab计算,时域分析统计特征中峰峰值和均值与光照关系的曲线为图3。
峰峰值的数值大小是最大值减去最小值,反映信号幅值的波动程度,而均值是信号的平均值,它可以反映出信号变化的基本趋向,这2个特征值可以表达出植物电信号在时域分析中的重要信息。由图3可以看出,在可变的光照条件下,燕子掌电信号的幅值单位大小为μV级,从LED灯光板亮度的10%(8 003 lx)一直到65%(27 819 lx),燕子掌电信号的峰峰值是不断上升的趋势,到80%(29 430 lx)时其特征值不如65%时大,表明植物电信号呈下降趋势,由植物电信号峰峰值先升后降,可知植物的光合作用先是增强,到达一个临界点,当光照继续增强时,植物受到光抑制[12],随后光合作用减弱,而均值变化的趋势不是很明显,基本呈现出水平直线形状,表明采集植物电信号的仪器系统稳定性良好,可以准确地检测提取植物电信号。
2.2 燕子掌电信号的频域分析
时域分析只能简单地从时域的角度出发分析信号特征,如果想要更加准确和客观地分析植物电信号,频域分析是必不可少的,因为植物电信号是一种包含多种不同频率的复杂信号,用功率谱分析法[13]可以很好地表征出电信号随光照度变化的频率特性,功率谱估计是把随时间变化的电信号幅值转化为功率谱随频率变化的功率谱图,比单纯从频率谱角度分析更加清晰直观,由于植物电信号的特性,经典的功率谱估计不适合植物电信号的频域分析,因为用此方法得出的估计值方差特性不好,频率分辨率低,而现代功率谱估计AR模型估计型估计的频率分辨率高,因此适合植物电信号短数据的处理,而且谱图平滑,有利于特征值的分析研究。鉴于此,本研究拟采用现代功率谱估计AR模型功率谱估计对植物电信号频域分析。
简单描述AR 模型功率谱估计:AR 模型是根据已观察到的样本数据来选择一个正确的模型,认为除样本数据外其他信号数据都是白噪声通过此模型产生的,因此与经典谱估计有区别(经典功率谱估计认为样本数据以外的数据全为0),这就有可能获取到更好的估计,避免了估计方差性能不好、分辨率低的不足,用这种方法得到的频率分辨率较高,因此它是一种高分辨率谱估计法。
图4为10%光照度下未消噪信号功率谱和经过小波软阈值消噪后的AR功率谱估计,可以看到植物电信号受外界噪声干扰严重,小波软阈值消噪可以很好地消除部分噪声,同时也说明使用AR功率谱估计植物电信号频域特性切实可行。
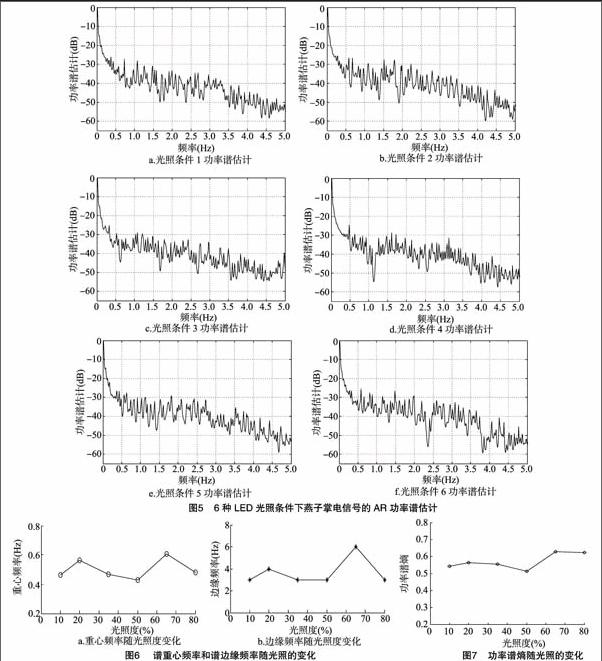
为了更好地显示频域特性,横轴的频率范围选择到 5 Hz,6种不同光照条件下功率谱估计图在Matlab软阈值消噪后信号的AR模型功率谱见图5。
由图5可知,植物电信号是一种低频信号,大部分能量集中在0.5 Hz以下,高频处也存在一些功率谱线,可以认为主要是外界噪声干扰造成的。从6组光照度图来看,前4组燕子掌功率谱特性曲线随光照度变化不大,但也可以明显看出在光照条件5时功率谱有明显变化,那就是0.5~2.5 Hz之间的功率谱比其他图要大,功率谱更集中,功率谱曲线有向高频变化的趋势,说明在光照条件5时比其他光照条件反应强烈,大致趋势也间接证明光合作用随光照度的不断增强先是增强,而后下降,这与时域峰峰值反映的信息基本一致。
为了更好地反映植物随光照变化的频域特性,分别计算谱重心频率(信号功率按频率的分布重心)、谱边缘频率(0 Hz 到该频率时的信号功率占总功率的95%)和功率谱熵 (反映信号的复杂程度),制成图表,如图6与图7。
由图6与图7可以看出,谱重心频率、谱边缘频率和功率谱熵变化规律呈现一致性,在光照度10%~50%变化不是很大,在50%~80%比前一范围反应要大,而且在选取的6个点处65%光照度下特征值最大,谱重心和谱边缘频率越大表明植物受抑制越小,功率谱熵大表明植物电信号复杂度变大,进而说明燕子掌电信号受光照变化而变化,光照度增强电信号不断增强,当光照度到达一定强度时,光合作用达到最强,之后各特征值呈现下降趋势,表明达到了光饱和,光合作用不再增强。
综合时域和频域各特征值的分析,各特征值反映的总体趋势较为一致,那就是在50%~80%光照度下植物电信号反应较为强烈,根据介值定理可知,在这一光照范围条件下必存在1个临界值点,植物电信号达到最大,光合作用最强。
3 结论
通过研究燕子掌的时域和频域特性,可以得出以下结论:燕子掌电信号是一种微弱低频信号,其植物电信号随着光照度的增强而增强,而且必存在1个临界点,但由于试验条件有限,不能准确得到,而且在实际环境条件下长期保持在某一精确的数值上比较困难,意义不大,而选取某一段时间对应的一定范围内的光照条件变得十分有意义,试验可得出燕子掌(Crassula obliqua)电信号最适宜的光照度范围为24 000~29 500 lx。从而为以后建立植物与各种环境因子模型打下了良好的基础。
参考文献:
[1]娄成后. 高等植物中电化学波的信使传递[J]. 生物物理学报,1996,12(4):739-745.
[2]王兰州,李海霞,林 敏,等. 植物微弱电波信号的时、频域分析[J]. 中国计量学院学报,2005,16(4):294- 298.
[3]卢善发. 植物电信号的传递途径与方式[J]. 植物学通报,1996,13(4):23-27.
[4]Hlavackova V,Krchnak P,Naus J,et al. Electrical and chemical signals involved in short-term systemic photosynthetic responses of tobacco plants to local burning[J]. Planta,2006,225(1):235-244.
[5]Shvetsova T,Mwesigwa J,Labady A,et al. Soybean electrophysiology effects of acid rain[J]. Plant Science,2002,162(5):723-731.
[6]Gallé A,Lautner S,Flexas J,et al. Environmental stimuli and physiological responses:the current view on electrical signalling[J]. Environmental and Experimental Botany,2014,114:15-21.
[7]Brenner E D,Stahlberg R,Mancuso S,et al. Plant neuro-biology:an integrated view of plant signaling [J]. Trends in Plant Science,2006,11(8):413-419.
[8]Volkov A G,Baker K,Foster J C,et al. Circadian variations in biologically closed electrochemical circuits in Aloe vera and Mimosa pudica[J]. Bioelectrochemistry,2011,81(1):39-45.
[9]Chatterjee S K,Ghosh S,Das S,et al. Forward and inverse modeling approaches for prediction of light stimulus from electrophysiological response in plants[J]. Measurement,2014,53:101-116.
[10]Luo Z Z,Li Y F,Meng M. EMG dnoising basedon the lifting wavelet transform[J]. Bioinformatics and Biomedical Engineering,2009,10:256-259.
[11]游榮义,徐慎初,陈 忠. 多通道脑电信号的盲分离[J]. 生物物理学报,2004,20(1):77-82.
[12]李新国,孟庆伟,赵世杰. 强光胁迫下银杏叶片的光抑制及其防御机制[J]. 林业科学,2004,40(3):56-59.
[13]王春兴. 基于MATLAB实现经典功率谱估计[J]. 曲阜师范大学学报,2011,37(2):59-62.
猜你喜欢
科学(2022年4期)2022-10-25
发明与创新(2019年43期)2019-11-16
学苑创造·A版(2018年5期)2018-05-28
现代园艺(2018年3期)2018-02-10
求学·理科版(2016年4期)2016-09-02
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
