
非O157产志贺毒素大肠埃希菌研究进展
2017-03-16 06:38巴鹏斌白向宁艾永循熊衍文
中国人兽共患病学报 2017年2期
巴鹏斌,孟 琼,白向宁,艾永循,熊衍文
非O157产志贺毒素大肠埃希菌研究进展
巴鹏斌1,2,孟 琼1,白向宁1,艾永循2,熊衍文1
产志贺毒素大肠埃希菌(Shiga toxin-producingEscherichiacoli,STEC)是一类能产生一种或一种以上志贺毒素的大肠埃希菌的总称,包括400余种血清型,其中以O157∶H7血清型为主。近年来,非O157 STEC在世界范围内引起散发感染和暴发的报道明显增加,但由于非O157 STEC血清型多样,菌株间表型差异较大,目前尚无一种有效的能用于所有非O157 STEC菌株分离的方法,因此非O157 STEC的流行情况可能被低估。本文就非O157 STEC 的病原学、致病机制、流行病学特征、实验室诊断、治疗和预防等方面的研究进展做一简要综述。
产志贺毒素大肠埃希菌;大肠埃希菌O157:H7;志贺毒素
产志贺毒素大肠埃希菌(Shiga toxin-producingEscherichiacoli,STEC)是一类能产生一种或一种以上志贺毒素的大肠埃希菌的总称,是重要的人兽共患病病原,可引起人类水样腹泻、出血性肠炎(hemorrhagic colitis,HC)以及高死亡率的溶血性尿毒综合征(hemolytic uremic syndrome,HUS)等[1]。
1977年Konowalchuk首次报道了一种对Vero细胞具有广泛不可逆损伤毒性的大肠埃希菌,随后O'Brien从大肠埃希菌中分离到一种与痢疾志贺菌产生的志贺毒素性质相似的毒素,称其为志贺样毒素(Shiga-like toxin,SLT),因能引起Vero细胞病变,也称Vero细胞毒素(Vero cytotoxin,VT)。目前已不再使用志贺样毒素这一名称,而直接称为志贺毒素(Shiga toxin,Stx)或VT毒素[1]。
自1982年美国首次报道从食物中毒患者以及HUS患者粪便标本中分离到产志贺毒素的大肠埃希菌O157∶H7以来,由O157∶H7所引起的大规模暴发流行及散发病例在世界范围内不断报道。O157∶H7成为STEC的主要血清型,也成为公共卫生领域面临的重要问题。近年来,非O157 STEC造成暴发流行以及散发感染的报道越来越多,尤其是2011年5月德国暴发了O104∶H4感染疫情,波及北美及欧洲16个国家,3 816人感染,其中54例死亡,22%的患者发生HUS,是迄今为止世界范围内造成HUS病例最多的一次非O157 STEC感染暴发,并由此引起人们对非O157 STEC的极大关注[2]。
1 病原学
STEC为革兰阴性短杆菌,形态与普通大肠埃希菌无异,大小为(0.4-0.7)μm×(1-3)μm,无芽胞,周身鞭毛,能运动,部分菌株无鞭毛,动力阴性。兼性厌氧,最适生长温度37 ℃,最适生长pH 7.0~7.4,在普通营养琼脂平板上生长良好,但对不同选择性培养基如麦康凯、山梨醇麦康凯、科玛嘉O157、科玛嘉STEC、科玛嘉ECC等中抑制剂的敏感性、糖的发酵能力不同而在生长状况、菌落大小及菌落颜色等方面有差异。2011年德国O104:H4暴发菌株除携带志贺毒素Stx2a外,还携带肠集聚性大肠埃希菌(enteroaggregativeE.coli,EAEC)的集聚性粘附质粒以及多重耐药质粒。因此该菌株同时具有产志贺毒素大肠埃希菌和肠集聚性大肠埃希菌的特点,故也称其为STEC EAEC O104∶H4[3]。
2 血清型别及其与疾病的关系
目前发现可从人类分离的STEC血清型有400余种[4],O157∶H7仍然是致病力强、流行范围广、最为常见的血清型。近年来,非O157 STEC 感染率不断上升,1994-2004年间日本、美国、澳大利亚、爱尔兰、德国、意大利等国家均出现了非O157 STEC暴发流行。 国家间菌株血清型分布略有不同,但是常见的血清型不足10种,其中O26、O111、O128、O103、O145在暴发的非O157 STEC血清型中占主导地位[5]。
Karmali等[6]根据血清型及其导致人类疾病暴发及散发的严重程度,提出了将STEC分成A到E 5类致病血清型,归纳如表1。
表1 产志贺毒素大肠埃希菌的血清型与人类疾病严重程度及暴发相关性
Tab.1 STEC serotypes, severity of disease and potential to cause outbreaks in humans

致病血清型SeropathotypeSTEC血清型STECserotype与疾病相关性Associationwithdisease与暴发相关性Involvementinoutbreaks与HUS、HC相关性AssociationwithHUSandHCAO157∶H7,O157∶NM高常见有BO26∶H11,O103∶H2,O111∶NM,O121∶H19,O145∶NM中不常见有CO5∶NM,O91∶H21,O104∶H21,O113∶H21,O121∶NM,O165∶H25andothers低不常见有DO7∶H4,O69∶H11,O103∶H25,O113∶H4,O117∶H7,O119∶H25,O132∶NM,O146∶H21,O171∶H2,O172∶NM,O174∶H8andothers低罕见无EO6∶H34,O8∶H19,O39∶H49,O46∶H38,O76∶H7,O84∶NM,O88∶H25,O98∶H25,O113∶NM,O136∶NM,O143∶H31,O156∶NM,O163∶NMandothers无无无
A型为致病力最强的血清型,包括O157∶H7和O157∶NM,B到E为非O157 STEC 致病血清型。B型为能引起严重疾病和暴发流行、导致HUS但感染率较O157低的血清型,包括O26∶H11,O103∶H2,O111∶NM,O121∶H19,O145∶NM;C型包括O91∶H21,O113∶H21, O104∶H21以及另外的8种血清型与散发HUS病例相关,但通常不引起暴发流行;D型包括O7∶H4,O69∶H11等14种血清型,仅与腹泻有关,不导致HUS或引起暴发;E型包括了与人类疾病无关的血清型。随着认识的深入,越来越多的血清型及其与疾病的关系将会被进一步揭示,如O104∶H4。
3 致病机制
目前对于非O157 STEC致病性的认识,主要来自STEC O157:H7。STEC 的致病性主要包括两个重要策略:1)由粘附因子介导的细菌对宿主肠道上皮细胞粘附和定植,2)产生志贺毒素和其它毒力因子。
胃酸(低pH值)是宿主在胃肠道对感染的重要防御机制,大肠埃希菌普遍耐酸,O157:H7 以及非O157 STEC 均被证实具有耐酸性,不同菌株之间有差异,并且酸的耐受性通过诱导可以提高[7]。非O157 STEC耐受胃酸到达肠道后,部分菌株通过LEE(locus of enterocyte effacement)岛编码的三型分泌系统紧密粘附于肠道上皮细胞并对宿主细胞造成粘附/抹平损伤(attaching and effacing,A/E)。对于LEE阴性的非O157 STEC菌株,则通过其他不同的粘附分子粘附定植于肠道上皮细胞,如eae阴性的O113:H21可以造成上皮微绒毛损伤,但并未发现细胞骨架重排以及明显的A/E损伤[8]。
志贺毒素是STEC的主要致病物质,是由糖苷酶活性的A亚基(32 kDa)与B亚基(7.5 kDa)构成的AB5型多肽亚基结构,其受体为神经酰胺三己糖苷(globotriaosylceramide, Gb3)。Stx通过与Gb3受体结合,干扰细胞蛋白质的合成,导致细胞损伤和死亡[9]。近年来也发现志贺毒素在引起细胞凋亡中发挥了重要作用。志贺毒素包括免疫反应不交叉的两类毒素,即志贺毒素1(Stx1)和志贺毒素2(Stx2),分别由stx1和stx2基因编码。志贺毒素可进一步分为不同的亚型,至今已报道的Stx2亚型有Stx2a,Stx2b,Stx2c,Stx2d,Stx2e,Stx2f,Stx2g,而Stx1相对保守,主要有Stx1a,Stx1c,Stx1d。每一菌株可以同时携带两种类型的毒素,也可携带多种毒素亚型[10]。
pO157质粒虽然普遍存在于O157∶H7分离株中,但在某些非O157的STEC菌株中也存在类似的质粒,其编码的肠溶血素(Ehx)等多种毒力因子可能与疾病有关[11]。其他毒素如枯草杆菌毒素SubAB是O113:H21菌株98NK2中的一种毒素,是一种丝氨酸蛋白酶,可引起小鼠微血管血栓、肾脏及其他器官坏死[8]。表2列举了目前人们了解的非O157 STEC毒力因子以及粘附相关因子。由于STEC菌株间的差异较大,不同菌株间携带的毒力因子种类会有很大差异,Stx是STEC的主要致病因素,而其他致病因子也在疾病进程中起重要作用。
4 流行概况
1982美国首次报道O157 STEC暴发感染病例,主要原因是食用未煮熟的牛肉汉堡。目前STEC感染呈世界性分布,各年龄段人群均可感染,儿童和老人通常易感且临床症状较重。STEC感染全年均能发病,夏秋两季是高发季节。美国1983-2002年间的960例非O157 STEC感染患者年龄中位数为12岁,四分位间距为3~37岁,且散发感染和暴发的高峰多出现在夏季[12]。
反刍动物是STEC的主要宿主,从牛体内分离出超过435种血清型,而从人体内分离到超过470种血清型的菌株[13]。在北美,牛是STEC最主要的宿主,而在有些国家,如澳洲,绵羊、山羊、鹿是STEC的主要宿主[14]。近年来,在猪、鸡、鹿、鸽子等家禽家畜等动物体内也陆续分离到非O157 STEC菌株[15-17]。我国饲养的猪,青藏高原的牦牛、鼠兔以及市售生肉类均为非O157 STEC的重要来源[18-21]。带菌动物往往是动物来源食品污染的来源,也可通过排泄的粪便污染当地的食物、草场、水源或其他水体及场所,造成交叉污染和感染,使病菌的传播机会和范围增大。人通过摄入受污染的食物或水,或直接与动物接触,导致散发感染或暴发。2007年3月丹麦暴发O26∶H11疫情,感染源来自有机发酵的牛肉香肠。2011年5月德国O104∶H4疫情暴发高峰时间集中在5月21日至5月22日,且感染者多为中年妇女,有22%的患者发展成为HUS,其中有88%HUS患者为成年人(平均年龄42岁),传染源来自有机芽苗菜(fenugreek sprouts)[22-23]。在2012-2013年间,日本由O26、O111、O103、O121、O145感染的病例也有报道,感染来源于幼儿园[24]。表3列举了自1986年以来全球部分由非O157 STEC引起暴发的报道。我们目前尚无非O157 STEC引起暴发的报道。
表2 非O157 STEC毒力因子及粘附相关因子
Tab.2 Virulence factors and adherence associated factors in non-O157 STEC

名称(Name)主要功能(Mainfunction)1.志贺毒素(Shigatoxin)STEC的主要致病因子,细胞毒性蛋白 Stx1a志贺毒素,与痢疾志贺的志贺毒素基本相同 Stx1c部分eae阴性STEC菌株产生的Stx1毒素变种 Stx1dStx1毒素变种 Stx2aStx2的原毒素,细胞毒性蛋白,可造成严重的人类疾病 Stx2bStx2毒素变种 Stx2cStx2毒素变种,可造成腹泻和HUS Stx2dStx2毒素变种,从绵羊中分离到的毒素变种,可导致人类疾病 Stx2eStx2毒素变种,导致猪水肿病的毒素变种,很少导致人类疾病 Stx2fStx2毒素变种,从鸽子中分离到的毒素变种,可导致人类疾病 Stx2gStx2毒素变种,从牛和污水中分离到的毒素变种2.其他毒素因子(Othertoxinfactors) CDT细胞致死肿胀毒素 EAST1肠聚集性耐热肠毒素 EHEChemolysin肠溶血素 Subtilasecytotoxin枯草杆菌毒素3.粘附相关因子(Adhesionrelatedfactors) LEE编码紧密素及受体,Ⅲ型分泌系统,诱导A/E损伤 Nle非LEE岛编码的效应分子3.1菌毛相关粘附因子(Pilusrelatedadhesionfactors) LpfO157-OI-141OI-141编码的O157∶H7极性长菌毛,在菌落形成中起重要作用 LpfO157-OI-154OI-154编码的O157∶H7极性长菌毛,在菌落形成中起重要作用 LpfO113O113∶H21粘附相关菌毛 OI-1fimbrialoperon推测的O157∶H7菌毛,其亚基与鼠伤寒沙门菌的StcA同源 OI-47fimbrialoperon推测的O157∶H7菌毛,其亚基与鼠伤寒沙门菌的StcA同源 SfpA可发酵山梨醇的O157∶NM菌毛蛋白3.2非菌毛粘附相关因子(Non-pilusadhesiverelatedfactors) EfaI介导STEC粘附于肠上皮细胞 Iha毒力岛(PAI)编码的粘附因子 OmpA外膜蛋白A,介导O157∶H7粘附HeLa和Caco2细胞 Saa由质粒编码的凝集粘附因子 ToxB质粒编码的粘附因子,可促进Ⅲ型分泌系统分泌蛋白4.蛋白酶(Proteases) EpeA质粒编码的丝氨酸蛋白酶,具有粘蛋白酶活性 EspP/PssA胞外丝氨酸蛋白酶,裂解胃蛋白酶A,人凝血因子V,对Vero细胞有细胞毒性 EspI/NleA与EspP功能相关的丝氨酸蛋白酶,降低胃蛋白酶A及人载脂蛋白A-I的水平 KatP质粒编码的过氧化氢酶 StcE质粒编码的,通过Ⅱ型分泌系统分泌的金属蛋白酶,可提高粘附性5.其他(Others) Etp质粒编码的Ⅱ型分泌系统 H7flagellin鞭毛素蛋白,促进与肠道上皮细胞的粘附,刺激肠上皮细胞产生前炎症细胞因子 H21flagellin鞭毛素蛋白,提高O113∶H21与结肠上皮细胞紧密结合 LPS脂多糖,诱导肠上皮细胞,血管内皮细胞产生促炎细胞因子,活化血小板 Urease尿素酶/脲酶,多存在于eae阳性菌株中,在体外通常不表达
表3 全球非O157 STEC引起暴发的部分报道
Tab.3 Some outbreaks of non-O157 STEC in the United States and worldwide
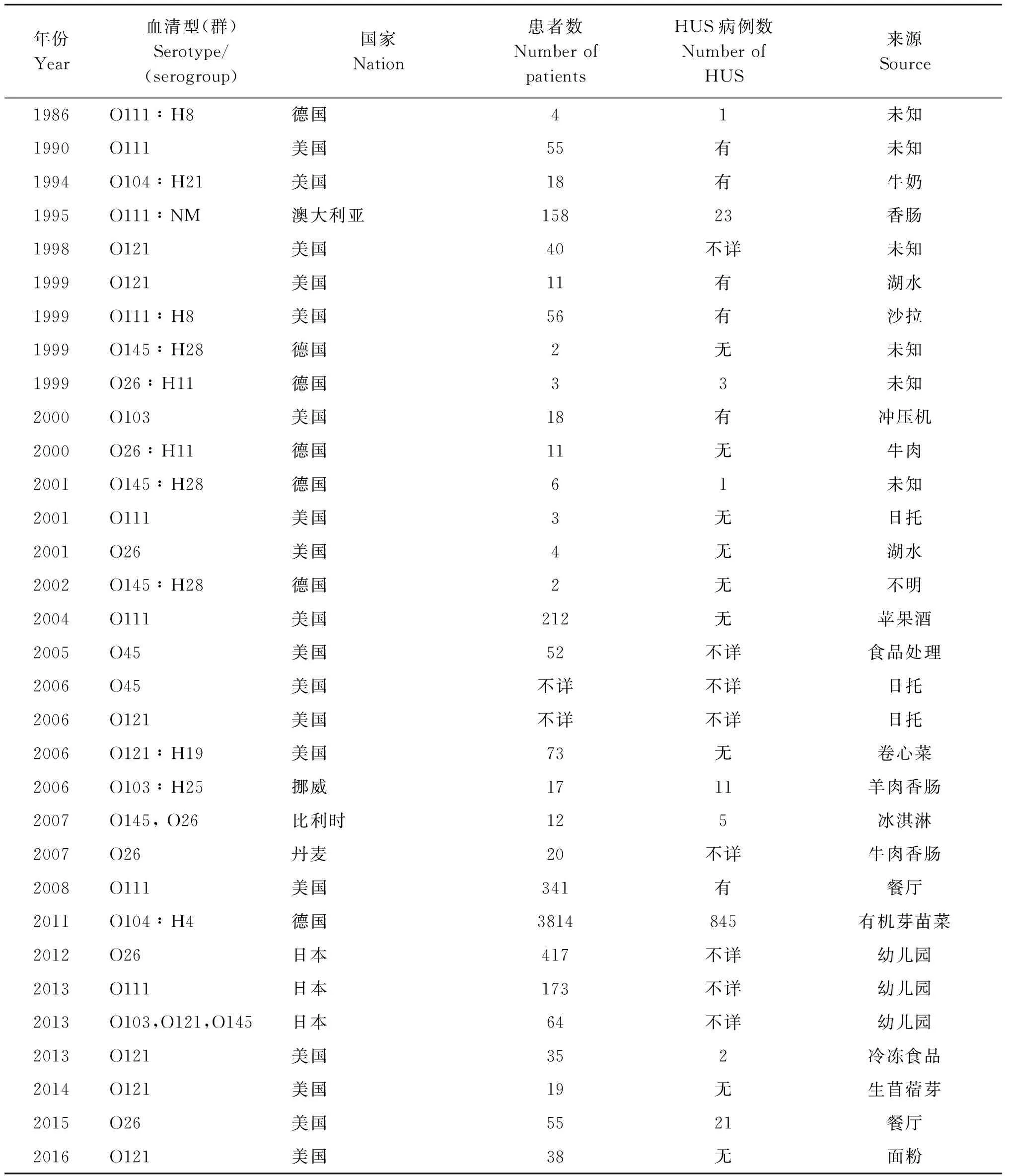
年份Year血清型(群)Serotype/(serogroup)国家Nation患者数NumberofpatientsHUS病例数NumberofHUS来源Source1986O111∶H8德国41未知1990O111美国55有未知1994O104∶H21美国18有牛奶1995O111∶NM澳大利亚15823香肠1998O121美国40不详未知1999O121美国11有湖水1999O111∶H8美国56有沙拉1999O145∶H28德国2无未知1999O26∶H11德国33未知2000O103美国18有冲压机2000O26∶H11德国11无牛肉2001O145∶H28德国61未知2001O111美国3无日托2001O26美国4无湖水2002O145∶H28德国2无不明2004O111美国212无苹果酒2005O45美国52不详食品处理2006O45美国不详不详日托2006O121美国不详不详日托2006O121∶H19美国73无卷心菜2006O103∶H25挪威1711羊肉香肠2007O145,O26比利时125冰淇淋2007O26丹麦20不详牛肉香肠2008O111美国341有餐厅2011O104∶H4德国3814845有机芽苗菜2012O26日本417不详幼儿园2013O111日本173不详幼儿园2013O103,O121,O145日本64不详幼儿园2013O121美国352冷冻食品2014O121美国19无生苜蓿芽2015O26美国5521餐厅2016O121美国38无面粉
5 实验室诊断
5.1 非O157 STEC分离培养 目前国内外均建立了针对STEC O157∶H7的分离培养方法,特别是O157免疫磁珠的使用,极大地提高了病原菌的分离率。对于非O157 STEC,目前国际上尚无统一有效的方法,其分离培养方法的优化,主要集中在选择性增菌和选择性分离两个环节。常用的增菌液主要有BPW,TSB,ECB等,为了提高选择性增菌效果,可加入选择性抑菌成分,如亚碲酸钾、新生霉素、头孢菌素、吖啶黄等以抑制其它菌群的生长。由于非O157 STEC菌株的多样性,选择性抑菌成分的种类及使用浓度也会对目标菌本身产生不同程度的影响,进而影响增菌效果。麦康凯琼脂(MacConkey agar)、山梨醇-麦康凯琼脂以及亚碲酸钾-头孢菌素山梨醇麦康凯琼脂广泛用于STEC O157∶H7的分离,也可以用于非O157 STEC的分离,但对其它肠道菌选择性低,使用具有一定的局限性。商品化的选择性培养基如科玛嘉O157、科玛嘉STEC等因其对STEC的选择特异性高,在STEC的筛选中得到广泛应用,但对不同非O157 STEC菌株的分离效果仍有差异[25]。因此,为了兼顾分离率和降低工作量,建议在对非O157 STEC分离时同时使用高选择性和低选择性的两种培养基。
5.2 非O157 STEC志贺毒素基因检测 无论是O157 STEC或是非O157 STEC,志贺毒素基因是STEC的共有特征,因此可以通过1)检测样品对Vero细胞的毒性;2)通过抗原免疫测定志贺毒素;3)核酸扩增试验(PCR)检测stx1/stx2基因等方法对STEC感染进行诊断。
6 治疗与预防
6.1 治疗 多数患者采取一般腹泻病的支持疗法,抗肠蠕动药物可能延长感染源的接触时间,具有一定风险,应尽量避免服用。对于HUS病人,尽可能通过补充液体、保持电解质平衡、透析等方法防止肾衰竭以及其他并发症,如中枢神经系统症状。由于抗生素的服用,可能导致HUS的发病率增加,且抗生素的使用并不能缩短病程,因此应尽量避免使用或在感染早期使用抗生素。 最近有研究表明,阿奇毒素可用于STEC O104∶H4的治疗[26]。
6.2 预防 发展中国家儿童STEC感染免疫的研究表明,早期感染STEC的患者,随着年龄的增长,继发感染STEC症状减轻或者无症状;非O157 STEC感染会引起HUS患者产生免疫反应。大量的体内体外实验表明,由志贺毒素刺激产生的抗体可以起到免疫保护作用。早期接种疫苗可能是预防STEC感染的一种有效策略[27]。然而,对于人类STEC的免疫接种,目前尚没有太大进展。
STEC主要经粪口途径传播,保证食品和饮用水的安全,控制传染源是主要的预防手段。STEC的感染和暴发流行与携带病菌的家畜家禽密切相关。因此,养殖场科学管理动物粪便,减少动物粪便的暴露;通过微生态试剂、抗生素、疫苗等手段提高家畜家禽对STEC感染的抵抗力,降低排菌量;加强对养殖、肉类屠宰加工等食品生产加工过程的检测和检测;提高对非O157 STEC的检测和监测能力;同时养成良好的个人卫生和生活习惯对于降低人群STEC的感染率具有重要意义。
7 结 语
STEC O157∶H7由于是导致出血性肠炎和HUS的重要病原而广受重视,而非O157 STEC感染因为诊断的局限性以及监测的不足往往被忽略。目前非O157 STEC已经在世界范围内引起散发感染和暴发流行,其感染率可以达到总STEC感染率的30%~50%,甚至在某些国家已经超过了O157∶H7。随着2011年德国O104∶H4疫情暴发,提示人们应加强对非O157 STEC认识,提高对这类感染的检测、监测、治疗以及预防控制能力。
[1] Bryan A, Youngster I, McAdam AJ. Shiga toxin producingEscherichiacoli[J]. Clin Lab Med, 2015, 35(2): 247-272. DOI: 10.1016/j.cll.2015.02.004
[2] Frank C, Werber D, Cramer JP, et al. Epidemic profile of Shiga-toxin-producingEscherichiacoliO104:H4 outbreak in Germany[J]. N Engl J Med, 2011, 365(19): 1771-1780. DOI: 10.1056/NEJMoa1106483
[3] Bielaszewska M, Mellmann A, Zhang W, et al. Characterisation of theEscherichiacolistrain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: a microbiological study[J]. Lancet Infect Dis, 2011, 11(9): 671-676. DOI: 10.1016/S1473-3099(11)70165-7
[4] Blanco M, Blanco JE, Mora A, et al. Serotypes, virulence genes, and intimin types of Shiga toxin (verotoxin)-producingEscherichiacoliisolates from cattle in Spain and identification of a new intimin variant gene (eae-xi)[J]. J Clin Microbiol, 2004, 42(2): 645-651. DOI: 10.1128/JCM.42.2.645-651.2004
[5] Bettelheim KA. The non-O157 Shiga-toxigenic (Verocytotoxigenic)Escherichiacoli; under-rated pathogens[J]. Crit Rev Microbiol, 2007, 33(1): 67-87. DOI: 10.1080/10408410601172172[6] Karmali MA, Mascarenhas M, Shen S, et al. Association of genomic O island 122 ofEscherichiacoliEDL 933 with verocytotoxin-producingEscherichiacoliseropathotypes that are linked to epidemic and/or serious disease[J]. J Clin Microbiol, 2003, 41(11): 4930-4940. DOI: 10.1128/JCM.41.11.4930-4940.2003
[7] Vidovic S, Korber DR.EscherichiacoliO157: Insights into the adaptive stress physiology and the influence of stressors on epidemiology and ecology of this human pathogen[J]. Crit Rev Microbiol, 2016, 42(1): 83-93. DOI: 10.3109/1040841X.2014.889654
[8] Croxen MA, Finlay BB. Molecular mechanisms ofEscherichiacolipathogenicity[J]. Nat Rev Microbiol, 2010, 8(1): 26-38. DOI: 10.1038/nrmicro2265
[9] Bergan J, Dyve Lingelem AB, Simm R, et al. Shiga toxins[J]. Toxicon, 2012, 60(6): 1085-1107. DOI: 10.1016/j.toxicon.2012.07.016
[10] Scheutz F, Teel LD, Beutin L, et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature[J]. J Clin Microbiol, 2012, 50(9): 2951-2963. DOI: 10.1128/JCM.00860-12
[11] Lim JY, Yoon J, Hovde CJ. A brief overview ofEscherichiacoliO157∶H7 and its plasmid O157[J]. J Microbiol Biotechnol, 2010, 20(1): 5-14. DOI: 10.4014/jmb.0908.08007
[12] Brooks JT, Sowers EG, Wells JG, et al. Non-O157 Shiga toxin-producingEscherichiacoliinfections in the United States, 1983-2002[J]. J Infect Dis, 2005, 192(8): 1422-1429. DOI: 10.1086/466536
[13] Bettelheim KA. The non-O157 shiga-toxigenic (verocytotoxigenic)Escherichiacoli; under-rated pathogens[J]. Crit Rev Microbiol, 2007, 33(1): 67-87. DOI: 10.1080/10408410601172172[14] Smith JL, Fratamico PM, Gunther NW 4th. Shiga toxin-producingEscherichiacoli[J]. Adv Appl Microbiol, 2014, 86: 145-197. DOI: 10.1016/B978-0-12-800262-9.00003-2
[15] Eggert M, Stuber E, Heurich M, et al. Detection and characterization of Shiga toxin-producingEscherichiacoliin faeces and lymphatic tissue of free-ranging deer[J]. Epidemiol Infect, 2012, 141(02): 251-259. DOI: 10.1017/S0950268812000246
[16] Ghanbarpour R, Daneshdoost S. Identification of shiga toxin and intimin coding genes inEscherichiacoliisolates from pigeons (Columba livia) in relation to phylotypes and antibiotic resistance patterns[J]. Trop Anim Health Prod, 2012, 44(2): 307-312. DOI: 10.1007/s11250-011-0021-0
[17] Tseng M, Fratamico PM, Manning SD, et al. Shiga toxin-producingEscherichiacoliin swine: the public health perspective[J]. Anim Health Res Rev, 2014, 15(1): 63-75. DOI: 10.1017/S1466252313000170
[18] Meng Q, Bai X, Zhao A, at al. Characterization of Shiga toxin-producingEscherichiacoliisolated from healthy pigs in China[J]. BMC Microbiol, 2014, 14: 5. DOI: 10.1186/1471-2180-14-5
[19] Bai X, Zhao A, Lan R, et al. Shiga toxin-producingEscherichiacoliin yaks (Bosgrunniens) from the Qinghai-Tibetan Plateau, China[J]. PLoS One, 2013, 8(6): e65537. DOI: 10.1371/journal.pone.0065537
[20] Bai X, Wang H, Xin Y, et al. Prevalence and characteristics of Shiga toxin-producingEscherichiacoliisolated from retail raw meats in China[J]. Int J Food Microbiol, 2015, 200: 31-38. DOI: 10.1016/j.ijfoodmicro.2015.01.018
[21] Bai X, Zhang W, Tang X, et al. Shiga toxin-producingEscherichiacoliin Plateau Pika (Ochotona curzoniae) on the Qinghai-Tibetan Plateau, China[J]. Front Microbiol, 2016, 7: 375. DOI: 10.3389/fmicb.2016.00375
[22] Buchholz U, Bernard H, Werber D, et al. German outbreak ofEscherichiacoliO104:H4 associated with sprouts[J]. N Engl J Med, 2011, 365(19): 1763-1770. DOI: 10.1056/NEJMoa1106482
[23] King LA, Nogareda F, Weill FX, et al. Outbreak of Shiga toxin-producingEscherichiacoliO104:H4 associated with organic fenugreek sprouts, France, June 2011[J]. Clin Infect Dis, 2012, 54(11): 1588-1594. DOI: 10.1093/cid/cis255
[24] Terajima J, Iyoda S, Ohnishi M, et al. Shiga toxin (verotoxin)-producingEscherichiacoliin Japan[J]. Microbiol Spectr, 2014, 2(5).
[25] Hirvonen JJ, Siitonen A, Kaukoranta SS. Usability and performance of CHROMagar STEC medium in detection of Shiga toxin-producingEscherichiacolistrains[J]. J Clin Microbiol, 2012, 50(11): 3586-3590. DOI: 10.1128/JCM.01754-12
[26] Hauswaldt S, Nitschke M, Sayk F, et al. Lessons learned from outbreaks of Shiga toxin producingEscherichiacoli[J]. Curr Infect Dis Rep, 2013,15(1): 4-9. DOI: 10.1007/s11908-012-0302-4
[27] Girard MP, Steele D, Chaignat CL, et al. A review of vaccine research and development: human enteric infections[J]. Vaccine, 2006, 24(15): 2732-2750. DOI: 10.1016/j.vaccine.2005.10.014
Research progress on non-O157 Shiga toxin-producingEscherichiacoli
BA Peng-bin1,2, MENG Qiong1, BAI Xiang-ning1, AI Yong-xun2, XIONG Yan-wen1
(1.StateKeyLaboratoryforInfectiousDiseasePreventionandControl,NationalInstituteforCommunicableDiseaseControlandPrevention,ChineseCenterforDiseaseControlandPrevention,Beijing102206,China;
2.MedicalCollege,JianghanUniversity,Wuhan430056,China)
Shiga toxin-producingEscherichiacoli(STEC) is a subset ofE.coliwhich can produce one or more Shiga toxins. Until now, more than 400 STEC serotypes were identified, but O157:H7 serotype is still the most prevalent. In recent years, more and more sporadic infections and outbreaks caused by non-O157 STEC strains were reported worldwide. There is no an effective method to identify all of heterogeneous non-O157 STEC strains, so the situation of non-O157 STEC infection may be under-estimated. This review is aimed to address the etiology, pathogenesis, epidemiology, diagnosis, treatment and prevention of non-O157 STEC.
Shiga toxin-producingEscherichiacoli;EscherichiacoliO157:H7; Shiga toxin
Xiong Yan-wen, Email: xiongyanwen@icdc.cn;AI Yong-xun,Email: iyx548@aliyun.com
10.3969/j.issn.1002-2694.2017.02.012
国家自然科学基金(No.81371762)资助
熊衍文,Email: xiongyanwen@icdc.cn; 艾永循,Email: iyx548@aliyun.com
1.中国疾病预防控制中心传染病预防控制所,传染病预防控制国家重点实验室,北京 102206; 2. 江汉大学医学院,武汉 430056
R378
A
1002-2694(2017)02-0156-07
2016-07-21 编辑: 李友松
Supported by National Natural Science Foundation of China(No.81371762)
猜你喜欢
首都食品与医药(2022年21期)2022-12-06
健康之友(2022年12期)2022-06-17
当代医药论丛(2022年1期)2022-01-27
医药前沿(2021年25期)2021-10-16
意林·作文素材(2021年15期)2021-10-08
昆明医科大学学报(2021年8期)2021-08-13
养猪(2020年6期)2021-01-27
学生天地(2019年33期)2019-08-25
中国感染与化疗杂志(2019年5期)2019-01-06
小学生导刊(高年级)(2016年4期)2016-04-08
