
21世纪桃树根的解剖学特征
2017-03-08 09:28:46刘建珍郭学民张立彬
河北科技师范学院学报 2017年4期
刘建珍,郭学民,肖 啸,张立彬
(河北科技师范学院1 园艺科技学院,2 生命科技学院;河北 秦皇岛,066600)
根是植物重要的地下营养器官,其主要功能是从土壤吸收水分和无机盐。根的解剖结构在一定程度上决定了它的吸收与输导能力,从而影响地上部分的生长及其对环境的适应性。21世纪桃树(河北科技师范学院园艺科技学院培育)是以毛桃(Prunuspersica)为砧木、21世纪桃树的芽为接穗嫁接而成的优质果树,和其他晚熟桃一样,在栽培过程中常发生冬害。郭学民等[1]报道,在风力强劲的冬季,21世纪桃树茎导管分子的巨型性容易使导管形成空穴和栓塞,水力结构破坏,致使输水系统受限,可能造成冬害。而茎输导系统的水分直接来源于根。因此,研究21世纪桃树根的解剖结构,对于进一步了解果树发生冬害的深层结构机制具有重要意义。
有关根的解剖结构(横切结构)及其吸收功能的研究主要集中在砧木矮化的预选指标上,一般认为,根皮率、皮率、木质率、根导管密度、根导管总面积等都与果树植株生长势呈负相关[2,3];对导管分子的研究往往集中在观赏植物[4,5]、分类上存在争议的植物[6]和一些果树[7~11]的茎上;刘霞等[12]比较柑橘实生砧木和2种体细胞杂种根系导管分子的结构。而在根导管与植物抗寒性关系方面,迄今仅见到李荣富等[3]对苹果砧木组织结构特性与抗寒矮化效应关系的报道,他们认为抗寒矮砧根系木质部的导管密度小且导管面积小,而乔砧木质部导管多而大。
笔者采用石蜡切片法和组织离析法,以晚熟品种21世纪桃为试材、典型耐寒品种珲春桃为对照,对2个材料根木质部和导管分子进行了观察,以期为21世纪桃冬害的深层解剖学机制研究、果树的遗传育种和生产实际提供解剖学参考资料。
1 材料和方法
1.1 材料
试验材料于2008年10月取自河北科技师范学院园艺科技学院试验基地。21世纪桃为5年生嫁接苗,珲春桃为5年生实生苗。
取样后洗去泥土,取根尖直径在1 mm左右且成熟度相对一致的部位,然后分为两部分固定,一部分用于石蜡切片,一部分用于导管离析。
1.2 方法
石蜡切片参照李正理[13]的方法略有改动。将根切成5 mm 长小段,FAA固定24 h,体积分数为150 mL/L氢氟酸30 ℃下软化12 d。将根漂洗后进行乙醇梯度脱水、包埋、切片,切片厚度10~14 μm,用番红- 固绿双重染色,中性树胶封片。Olympus BX51显微镜下观察,DP70数码相机拍照。2种材料根横切解剖指标测定时分别选择20张切片进行测定。
导管离析参照郭学民等[1]的方法进行。以Olympus BX51显微镜观察,DP70数码相机拍照,每种材料导管分子测定100个,依据Tippo的描述方法对导管进行描述[14]。
1.3 数据统计
利用DPS v3.01专业版数据处理系统,对2个品种各项数据进行Duncan新复极差法比较。
2 结果与分析
2.1 根横切结构特征
2个品种桃树根横切面的主要结构从外向内均为周皮、次生韧皮部和次生木质部,其中周皮和次生韧皮部共同构成根皮。21世纪桃根皮率比珲春桃低11.2%,21世纪桃维管射线宽、数量多,而珲春则相反(图1),说明21世纪桃维管射线发达。加之,21世纪桃的导管密度和木质部面积/横断面积均大于珲春桃的(表1),这为生长季节旺盛的生长势奠定了良好的结构基础。但是21世纪桃的导管总面积和导管总面积/木质部面积却小于珲春桃的(表1)。

表1 供试材料横切解剖构造比较
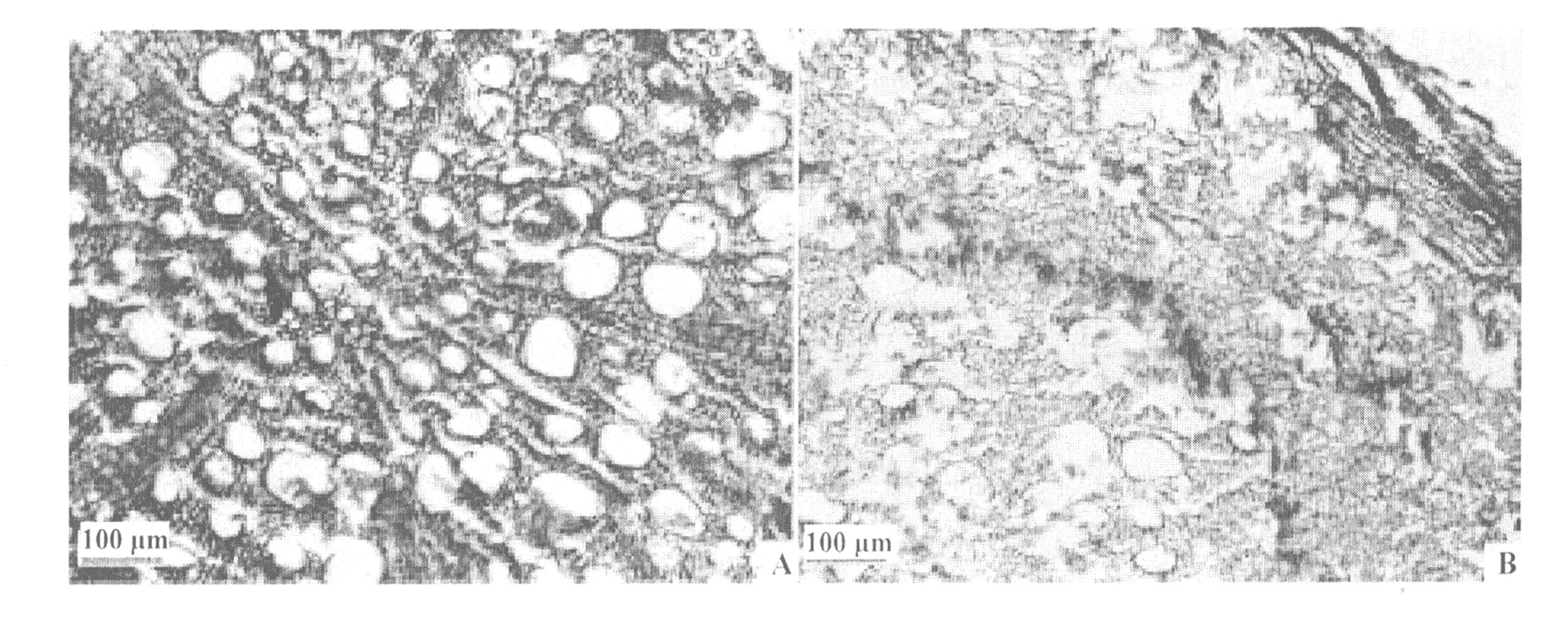
图1 21世纪桃和珲春桃树根横切结构A:21世纪桃;B:珲春桃
2.2 导管分子特征
对2个品种桃树根次生木质部进行离析处理,分离出不同式样的导管分子(图2和图3)。从导管分子侧壁次生增厚和木质化的方式看,都属于孔纹导管。导管分子管间纹孔均为互列纹孔式。从导管分子的长度和宽度(直径)看,21世纪桃的导管长度比珲春桃的长13.96%(p﹤0.01),而导管宽度却比珲春桃的小40.08%(p﹤0.01)(图4)。从导管分子长度分布情况看,2种材料导管分子长度主要分布于151~350 μm,但是21世纪桃长度大于250 μm的导管分子数目比珲春桃的多(图5);从导管分子宽度分布情况看,21世纪桃的导管分子分布在15~75 μm,但是珲春桃的宽度大于76 μm的导管分子数远远多于21世纪桃的(图6)。
从穿孔板的式样看,在所观察的材料中,导管分子均为单穿孔(图2,图3)。
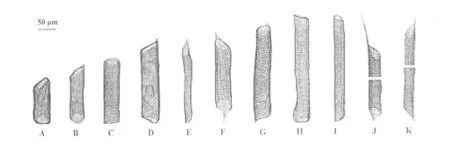
图2 21世纪桃根导管分子形态A,I:一端壁水平,一端壁倾斜;B,D,E:两端倾斜,一端具尾;C,H:两端近水平,无尾;F,G,J,K:两端倾斜且具尾
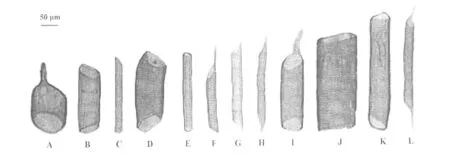
图3 珲春桃根导管分子形态A,F,I:两端倾斜,一端具尾;B,E,J:两端近水平,无尾;C:一端倾斜,无尾;D:两端倾斜,无尾;G,H,L:两端倾斜,两端具尾;K:一端近水平,无尾

图4 导管分子平均长度与平均宽度 图5 导管分子长度分布情况
从导管分子尾的有无来看,2个品种桃树根导管分子有的两端具尾,有的一端具尾一端无尾,有的两端无尾,但2个品种间导管分子具尾的类型所占的比例不同。在无尾的导管分子中,珲春桃无尾导管分子所占比例显著大于21世纪桃的,而两端具尾和一端具尾的导管分子所占的比例与无尾的导管呈相反趋势(图7)。
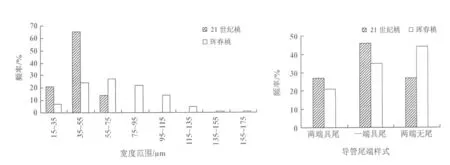
图6 导管分子宽度分布情况 图7 导管分子不同尾的类型出现的频率
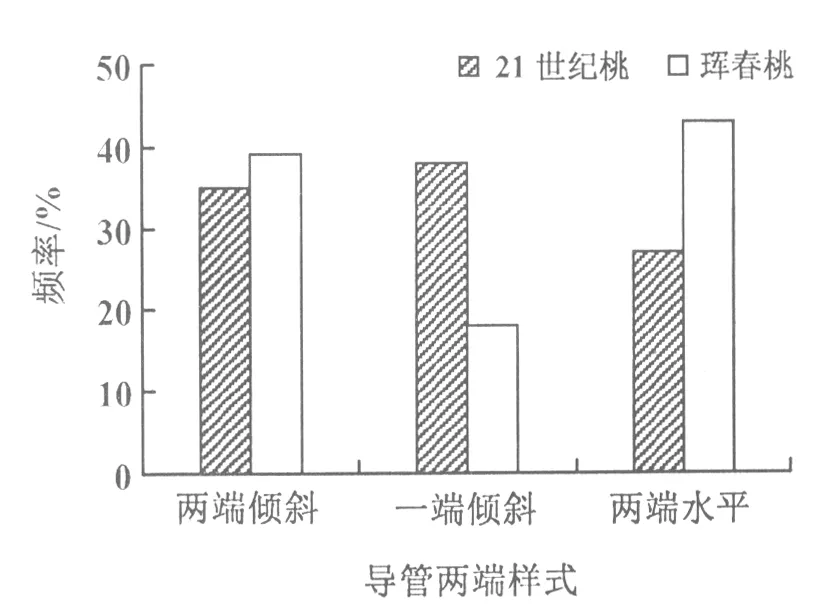
图8 导管分子端壁倾斜度的情况
从导管分子端壁倾斜性来看,珲春桃两端壁均为水平的所占比例显著高于21世纪桃的,一端壁水平、一端壁倾斜的导管分子所占的比例在2个材料中呈相反趋势,两端壁均倾斜的导管分子所占比例在2个品种中没有明显差异(图8)。
3 讨 论
3.1 根横切结构与21世纪桃生长的关系
维管射线是根的横向运输结构,次生木质部导管中的水分和无机盐可经维管射线运至形成层和次生韧皮部,而次生韧皮部中的有机养料可通过维管射线运至形成层和次生木质部。根皮率与果树长势呈负相关[2]。本次研究发现,21世纪桃根的维管射线发达,木质部面积/横断面积高,即根皮率低,导管密度高(图1,表1),说明21世纪桃木质部输导能力较强,这正是其生长势旺盛的解剖学原因,也与生产实际相一致。
3.2 根横切结构与21世纪桃冬害的关系
本次研究发现,21世纪桃根皮率低,所以在寒冷的冬天,其根皮不可能有效地保护木质部的输导功能。加之,21世纪桃根导管总面积和导管总面积/木质部面积低,在没有根皮有效的保护作用下,输导能力可能降低。然而,地上部分在强劲的寒风袭击下,蒸腾加剧,而根部的输导能力不能满足地上部分对于水分的需要,即可能产生冬害。
3.3 导管分子的式样与功能
在实际生产中,21世纪桃树是比较容易遭受冬害的果树。从根导管分子的性状看,21世纪桃导管长,导管宽度(直径)小(图4),具尾和端壁倾斜比率高(图7,图8)。在严冬里,这些特点强烈地影响了21世纪桃根的输导能力。由于植物地下部分与地上部分的维管组织是一脉相通的,联系地上部分茎的导管性状看,21世纪桃茎的迎风面和背风面导管分子平均宽度为53.06 μm[1],而根的则为43.8 μm(图4),根中宽度(直径)较小的导管分子的输导能力,在冬天不能满足茎中宽度(直径)较大的导管分子对于水分的需求,即根茎导管分子直径不匹配,容易造成冬害。
相反,珲春桃是抗寒性能较好的品种。从根导管分子性状看,珲春桃导管短,导管宽度(直径)大(图4),两端无尾和两端壁水平的比率高(图7,图8),在冬季,根导管输导能力强。珲春桃茎的迎风面和背风面导管分子平均宽度为52.07 μm[1],而根的则为73.1 μm(图4),很显然,其根的导管分子宽度(直径)远远大于茎的,即使在严寒胁迫下,根的导管分子输导能力也可满足茎的宽度(直径)较小的导管分子对水分的需求,即根茎导管分子匹配,果树抗冬害能力可能较强。
穿孔板上的穿孔是相连导管分子输导水分和无机盐的通道,被子植物导管分子的单穿孔板是由梯状穿孔板的横闩消失演化而成的,因此单穿孔板较为进化[15]。2个供试材料根导管分子均为单穿孔,管间纹孔均为互列纹孔式,部分导管无尾且端壁水平,这些均为导管分子系统演化过程中较为进化的特征,但在不同类型的导管分子上仍存在着具有尾以及比较倾斜的端壁等原始的形状。
[1] 郭学民,肖啸,徐兴友,等.21世纪桃树次生木质部导管分子特征的观察[J].果树学报,2008,25(1):22- 26.
[2] 赵大中,罗先实,鲁俊良,等.砧木矮化预选指标的解剖学研究[J].果树科学,1995,12(4):219- 223.
[3] 李荣富,梁艳荣,梁莉,等.苹果砧木组织结构特征与抗寒矮化效应的关系[J].内蒙古农业大学学报,2003,24(3):49- 52.
[4] 陈树思,唐为萍.银桦次生木质部导管分子观察研究[J].广西植物,2004,24(4):380- 382.
[5] 陈树思,唐为萍.沉香(Aquilariaagallocha)次生木质部导管分子研究[J].华中师范大学学报(自然科学版),2004,38(4):486- 489.
[6] 林祁,邬志荣,钟学敏,等.东亚八角科比较木材解剖[J].林业科学,2010,46(1):91- 100,Ⅰ,Ⅱ.
[7] 杨佩芳,郝燕燕,田彩芳.苹果短枝型品种导管分子的解剖学研究[J].园艺学报,2000,27(1):52- 54.
[8] 王彦涵,高建平,乔春峰,等.国产五味子科植物导管分子的比较解剖[J].广西植物,2003,23(3):226- 230.
[9] 杨佩芳,刘和,高建平,等.桃、苹果枝条次生木质部细胞的解离研究[J].果树科学,1995,12(3):146- 150.
[10] 王成霞,李培环,王永章,等.桃树枝条的解剖结构与树体矮化的相关性研究[J].莱阳农学院学报(自然科学版),2006,23(3):185- 188.
[11] 刘和,杨佩芳,牛吉山,等.杏李次生木质部导管分子的解剖学研究[J].山西农业大学学报,1996,16(4):404- 407.
[12] 刘霞,彭抒昂,郭文武.三种柑橘实生砧木及其两种体细胞杂种根系解剖结构的比较[J].园艺学报,2008,35(9):1 249- 1 254.
[13] 李正理.植物组织制片学[M].北京:北京大学出版社,1996:91- 92.
[14] Tippo O.A list of diagnostic characteristics for description of dicotyledonous woods[J].Trans Illinois Acad Sci,1941,34:105- 106.
[15] Fahn A.植物解剖学[M].吴树明,刘德仪,译.天津:南开大学出版社,1990:497.
猜你喜欢
小主人报(2022年5期)2022-04-01 01:12:02
今日农业(2021年16期)2021-11-26 06:05:34
今日农业(2021年21期)2021-11-26 05:07:00
植物研究(2021年2期)2021-02-26 08:40:10
小学生导刊(2016年13期)2016-12-01 06:02:56
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
安徽农学通报(2014年7期)2014-04-29 03:00:24
中国边疆民族研究(2012年0期)2012-02-13 02:42:38
