
银杏内酯B对阿尔茨海默病大鼠海马Synapsin-1、Beclin1 和LC3表达的影响❋
2017-03-08 11:18田新红游言文徐玉英于世奇
中国中医基础医学杂志 2017年12期
田新红,郝 莉,游言文,徐玉英,于世奇
(河南中医药大学基础医学院,郑州 450008)
学习记忆障碍是阿尔茨海默病(alzheimer’s disease, AD)的主要临床表现,β淀粉样蛋白(amyliod-βprotein, Aβ)沉积形成老年斑、神经纤维缠结、突触丢失是AD的主要病理学改变,其中Aβ引起的突触丢失是AD学习记忆障碍的主要原因[1-2]。突触蛋白1(Synapsin-1)是一种与突触结构、功能密切相关的膜蛋白。自噬又称为Ⅱ型细胞死亡,是细胞在自噬相关基因(autophagyrelated gene,Atg)调控下利用溶酶体降解自身受损细胞器和大分子物质的过程。自噬相关基因1(Beclin1)和微管相关蛋白轻链3 (microtubule-associated protein 1 light chain 3, LC3) 是哺乳动物细胞自噬的主要相关基因。已有研究证实,AD患者与动物模型大脑自噬水平上调类同[3],自噬是突触丢失的主要方式[4-5]。
中药银杏叶提取物可改善AD相关学习记忆力下降,而银杏内酯B(Ginkgobalide B,GB)是银杏叶提取物的主要活性成分,具有保护神经元、促进学习和改善记忆的作用[6]。实验证实,GB可改善AD相关的学习记忆缺陷[7],然而其具体作用机制尚未阐明。 本研究建立Aβ1-42诱导的AD大鼠模型,腹腔注射中药单体GB,从行为学和分子生物学角度探讨GB改善AD相关学习记忆能力下降及突触相关蛋白Synapsin-1、自噬相关基因Beclin1和LC3在其中的作用,为中药单体防治AD提供理论和实验依据。
1 材料与方法
1.1 动物与试剂仪器
雄性清洁级 Sprague-Dawley(SD)大鼠,体质量180~220 g,由河南实验动物中心提供(合格证号SCXK (豫)2010-0002)。GB购自中国食品药品检定研究院;Aβ1-42购自Sigma公司;兔单克隆一抗Synapsin-1、Beclin1和LC3购自Cell Signaling公司;辣根过氧化物酶标记羊抗兔二抗购自北京中杉金桥公司;Tecnai-12透射电镜购自飞利浦公司;莱卡显微成像系统购自莱尔公司;全自动凝胶成像系统购自天能公司;VCX130 超声破碎仪购自美国Sonics 公司;Stepone 实时荧光定量PCR仪购自美国Applied Biosystems公司。
1.2 方法
1.2.1 AD动物模型建立 SD大鼠使用10%水合氯醛腹腔注射麻醉,头固定于脑立体定位仪上,参考大鼠脑立体定位图谱:正中做一切口暴露出前囟,以前囟为原点向后4 mm,旁开2.5 mm 左右各钻一孔,硬脑膜下3 mm至两侧海马。由微量注射器将2 μL(10μg)Aβ1-42在10 min内注射完毕,留针10 min,使得Aβ1-42充分浸润局部组织,术后均注射抗生素防止感染。假手术组大鼠注射等量生理盐水。
1.2.2 分组及治疗 将SD大鼠按随机数字表法分为假手术组(8只)、模型组(8只)、GB低剂量组(15只)、中剂量组(15只)5组和高剂量组(15只)。模型造成3 d后,GB低、中、高剂量组腹腔注射GB,剂量分别为每天2.5、5和7.5 mg/kg,假手术组和模型组给予等量生理盐水,均为15 d。
1.2.3 Morris水迷宫检测学习记忆能力 测试期间室内保持安静,水池内注入清水并加入墨汁至水不透明,水面高出平台2 cm。第1天进行预实验让大鼠适应环境,同时将其引导到平台。其后4 d,大鼠每日从不同入水点放入水中, 找到平台时间即逃避潜伏期,逃避潜伏期记录为60 s。第6天撤除平台,将大鼠从任一处放入水池,测试60 s内穿越原平台位置的次数。行为学检测期间仍对大鼠进行GB治疗。
1.2.4 透射电镜检测自噬 水迷宫实验结束后1 h,大鼠用10%水合氯醛腹腔注射麻醉,打开胸腔暴露心脏,灌注生理盐水冲洗血管,灌注2.5%戊二醛固定。剥离鼠脑,取约1 cm3海马CA1区组织,置于戊二醛中固定并用磷酸漂洗,乙醇脱水、丙酮包埋,固化、切片、染色,透射电镜观察、拍片。
1.2.5 免疫组织化学检测Synapsin-1、Beclin1和LC3表达 灌注预冷的多聚甲醛进行固定。快速断头取脑,多聚甲醛固定24 h,乙醇梯度脱水、二甲苯透明、石蜡包埋切片。脱蜡、修复、PBS清洗,血清封闭,37 ℃孵育20 min;滴加一抗(兔抗Synapsin-1、Beclin1和LC3,稀释1∶200),37 ℃孵育过夜;加HRP标记二抗,孵育、DAB显色,中性树胶封片。选取海马CA1区不重叠的5个高倍视野,计算其阳性光密度值。
1.2.6 荧光定量PCR检测海马组织Synapsin-1、Beclin1和LC3-Ⅱ mRNA表达 按照Trizol试剂盒说明书从冰冻海马组织中快速提取总RNA并逆转录成 cDNA。Synapsin-1上游引物:5′-CTGGACTGCGGTATTGAGAC-3′,下游引物:5′-CCGGGTGCGGTAGAGTAAGC-3′;Beclin1上游引物:5′-GTTCTCTCAGTTGCCTTTC-3′,下游引物:5′-AGCTGTAACCTGTCGCCGAGTCCC-3′;LC3-Ⅱ上游引物:5′-CGGGTTGAGGAGACACACAA-3′,下游引物:5′-ATGAGCCGGACATCTTCCAC-3′;GAPDH上游引物:5′-CTACCCACGGCAAGTTCAAT-3′,下游引物:5′-GGATGCAGGGATGATGTTCT-3′,均由上海生工生物有限公司合成。按照试剂盒说明书进行扩增,PCR反应总体积为25 μL,反应条件:95 ℃ 预变性5 min,94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸40 s,35个循环;72 ℃延伸10 min。由系统软件自动生成循环域值(Ct)。每个样品均做3个平行孔,取平均值作为该样品的Ct值,并计算出Synapsin-1、Beclin1和LC3-Ⅱ相对表达量。
1.3 统计学方法

2 结果
2.1 各组大鼠 Morris 水迷宫结果
表1显示,与假手术组比较,模型组大鼠的平均逃避潜伏期延长(P<0.05),去掉平台后模型组穿越原平台次数明显减少(P<0.01);腹腔注射银杏内酯B 15 d后,中、高剂量组平均逃避潜伏期时间缩短(P<0.05),穿越原平台次数增加(P<0.05),行为学结果提示GB具有改善 AD 大鼠学习记忆下降的能力。
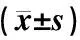
表1 各组大鼠 Morris水迷宫实验结果比较
注:与假手术组比较:1)P<0.05,2)P<0.01;与模型组比较:3)P<0.05,4)P<0.01
2.2 各组大鼠海马CA1区神经元电镜观察自噬结果
图1显示,与假手术组比较,模型组大鼠电镜自噬体结构较明显。随着GB治疗剂量的增加 ,可观察到自噬体结构逐渐不明显。
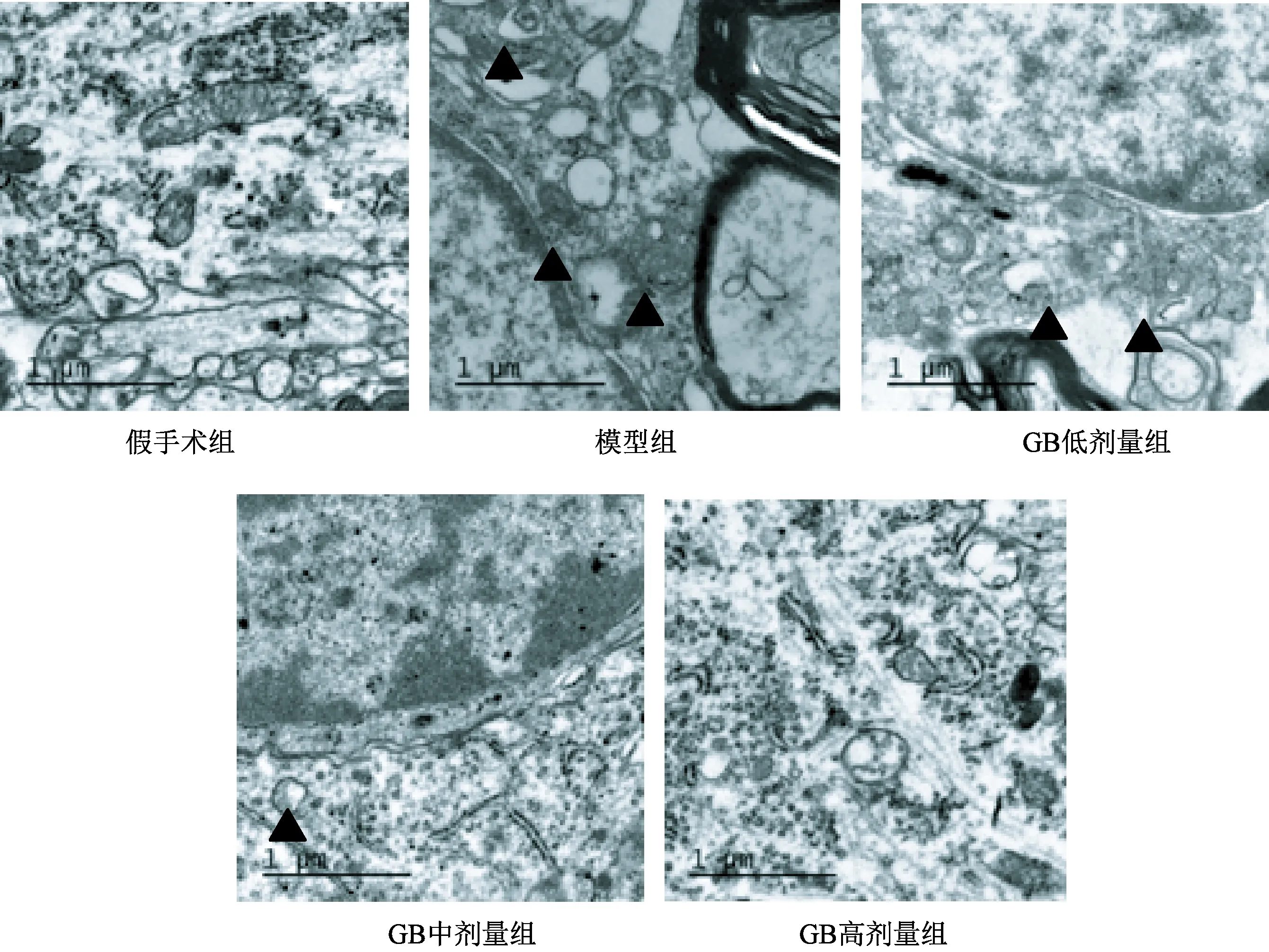
图1 各组大鼠海马CA1区神经元自噬电镜观察结果(20000×)注:▲代表自噬体
2.3 各组大鼠海马CA1区Synapsin-1、Beclin1和LC3蛋白和mRNA表达
图2表2显示, Synapsin-1的阳性表达部位主要在细胞核周围的细胞质中,呈深黄色或褐色,细胞核呈蓝色。与假手术组比较,模型组大鼠海马CA1区Synapsin-1蛋白表达降低(P<0.05)呈弱阳性。随着腹腔注射GB剂量的增加,Synapsin-1蛋白表达逐渐上升呈强阳性,其中GB高剂量组作用最强(P<0.05)。 LC3和Beclin-1主要定位于海马神经元细胞质中,阳性表达呈棕色。假手术组仅有少量阳性细胞,且着色浅淡;模型组中LC3和Beclin-1胞质的染色程度加深,阳性细胞数明显增多(P<0.05)。与模型组比较,随着腹腔注射GB剂量的增加,Beclin-1的表达程度及阳性细胞数逐渐减少(P<0.05),而LC3有降低的趋势,但差异无统计学意义(P>0.05),提示GB具有提高 AD大鼠海马CA1区Synapsin-1,降低Beclin-1蛋白表达的能力(图中箭头分别表示阳性细胞)。
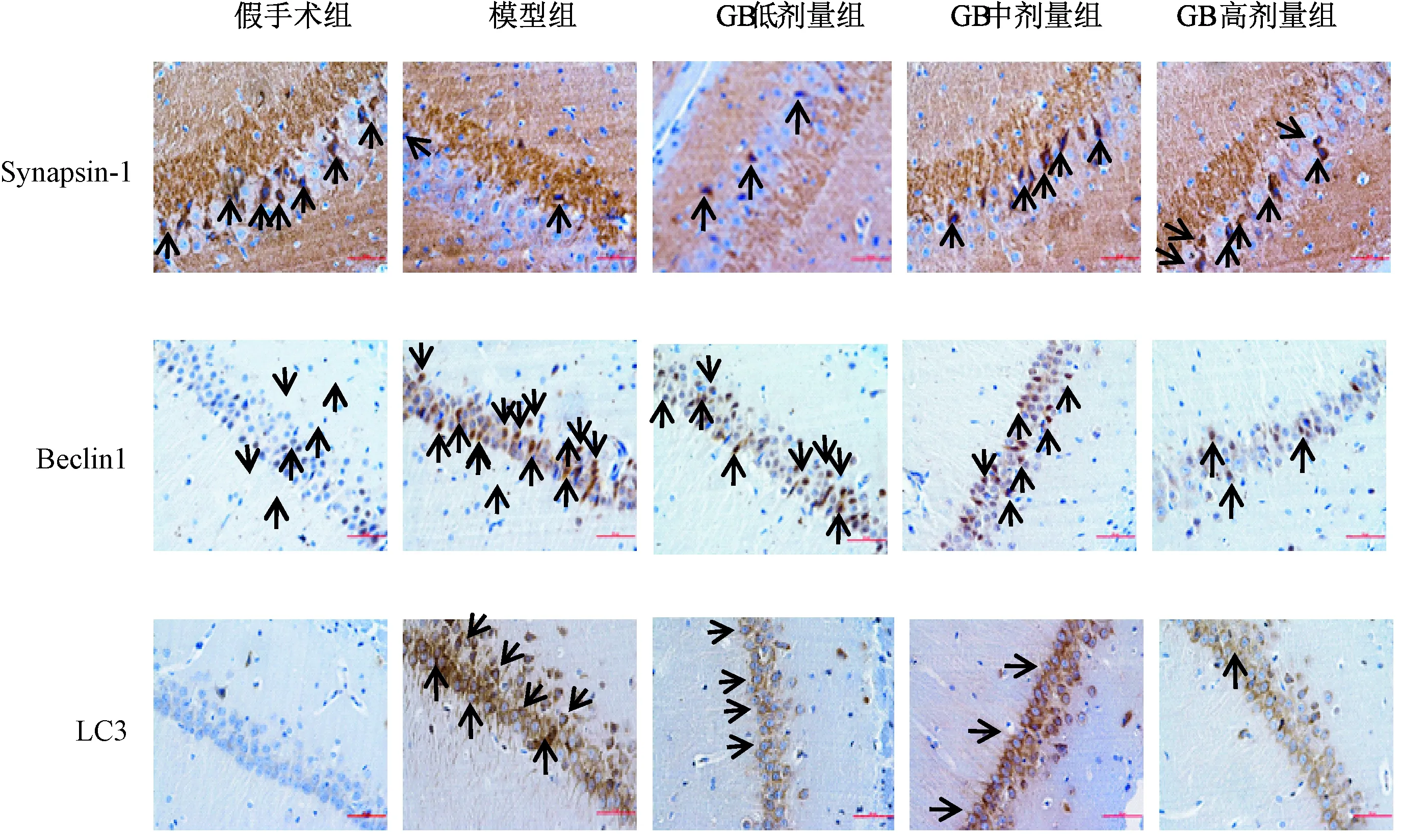
图2 各组大鼠海马CA1区免疫组化Synapsin-1、Beclin1和LC3蛋白表达(400×)
表2显示,与假手术组比较,模型组大鼠海马组织Synapsin-1 mRNA水平下降(P<0.01),Beclin1和LC3-Ⅱ mRNA水平表达升高(P<0.05);与模型组比较,GB治疗中、高剂量组Synapsin-1和Beclin1 mRNA相对表达量分别增加和降低(P<0.05);与模型组比较,GB治疗中剂量组LC3-ⅡmRNA表达量有降低趋势(P>0.05),高剂量组表达量明显降低(P<0.05)。
表2 各组大鼠海马CA1区Synapsin-1、Beclin1和LC3蛋白和mRNA表达
注:与假手术组比较:1)P<0.05,2)P<0.01;与模型组比较:3)P<0.05,4)P<0.01
3 讨论
本研究选择使用Aβ1-42硬脑膜下注射大鼠两侧海马建立AD动物模型。学习记忆障碍是AD的主要临床表现,Morris水迷宫是检测学习记忆能力的主要行为学方法。它通过让受试动物在水环境中寻找隐匿平台,并分析其找到平台所需时间(逃避潜伏期)及动物穿越隐匿平台的次数来判断空间学习记忆能力。本研究通过Morris水迷宫检测AD动物模型的学习记忆能力,结果与正常组比较,AD模型组大鼠平均逃避潜伏期明显延长,去除平台后模型组穿越原平台的次数减少,说明Aβ1-42建立的大鼠模型学习记忆能力下降,提示AD动物造模成功,可被用于后续的实验。
Morris水迷宫中逃避潜伏期越短、穿越隐匿平台的次数越多,表明受试动物的空间学习记忆能力越好。本研究在Morris水迷宫行为学检测时观察到,与AD模型组比较,随着腹腔注射GB剂量的增加,治疗组大鼠逃避潜伏期时间缩短,穿越原平台次数明显增加,提示中、高剂量即5和7.5 mg/kg的中药单体GB具有改善AD大鼠学习记忆下降的能力。
已有研究证实,AD病人和动物模型脑中均存在突触丢失[8-9],且突触丢失较神经元数量减少更显著。突触丢失是AD的一个主要病理学特征,是学习记忆障碍的结构基础[10-11]。促进突触丢失的主要原因,是病理情况下β淀粉样前体蛋白(amyliod precursor protein,APP)被2种分泌酶切割产生Aβ,主要是由39~43个氨基酸组成的Aβ40和Aβ422种分子,其中Aβ42比Aβ40疏水性强,易聚集、 易导致AD老年斑块的形成[12]。Aβ是AD最主要的突触毒素,可破坏海马脑片或动物的长时程增强,降低树突棘密度影响学习记忆[13]。Synapsin-1是进化中高度保守的蛋白质,定位于突触囊泡膜上,是一种与突触结构和功能密切相关的特征性膜蛋白,其数量和分布密度可间接反映突触的密度。AD小鼠模型中,SynapsinⅠ表达水平明显降低。海马是参与学习记忆功能的重要结构,也是AD病人脑中最易受累的区域之一。本研究观察到,Aβ1-42建立的AD大鼠模型海马组织SynapsinⅠmRNA和蛋白表达下降与文献基本一致。使用中、高剂量的银杏内酯B可增加SynapsinⅠ水平。GB是银杏叶提取物的主要单体。本课题组前期实验发现,银杏叶提取物可改善衰老鼠学习记忆力下降[14-15],且与其提高海马神经元SynapsinⅠ表达相关。AD是与衰老密切相关的疾病,学习记忆力下降是其主要临床症状。结合本研究观察到的结果提示,银杏叶提取物主要可能是通过其中的单体GB改善衰老相关的学习记忆,而这种改善可能与大鼠海马突触蛋白SynapsinⅠ表达水平增加有关。
本研究通过透射电镜观察各组大鼠海马CA1区神经元超微结构发现,模型组出现的自噬体大多位于细胞核旁,GB可减少细胞内自噬体数量。自噬与突触丢失相关,AD动物模型SAMP8小鼠的海马神经元突触丢失伴随LC3阳性细胞数的增加[5]。LC3 是酵母 Atg8 的哺乳动物同源体,是公认的哺乳动物细胞自噬标志物。免疫组化方法检测到AD模型大鼠海马LC3蛋白表达增加,与文献基本一致;而GB对LC3影响不明显差异无统计学意义。自噬发生时,pro-LC3 被加工成胞浆型LC3(即LC3-Ⅰ)。经自噬相关基因-5(autophagy related gene-5, ATG5) 和自噬相关基因-3(autophagy related gene-5, Atg3)催化,LC3-Ⅰ酶解掉一小段多肽,转变为一种膜结合形式即 LC3-Ⅱ。LC3-Ⅱ是由LC3-Ⅰ转化而来,LC3-Ⅱ的含量已被证实与自噬体形成有关。由于哺乳动物细胞内LC3-Ⅱ蛋白能够调控自噬体形成,其表达水平的高低在某种程度上可反映细胞自噬活性[16]。为进一步探讨银杏内酯B对LC3的影响,又检测了LC3- ⅡmRNA,结果显示中、高剂量GB可降低AD模型大鼠海马LC3-ⅡmRNA表达水平。Beclin1基因是酵母ATG6的同系物,调节启动自噬体的形成,也是哺乳动物细胞自噬的特异性基因。有学者发现,Beclin1在AD脑中的表达增加。本研究AD模型大鼠海马Beclin1 mRNA和蛋白表达升高,腹腔注射不同剂量的GB后,Beclin1表达水平下降。
临床上银杏叶提取物已被应用于缺血性心脑血管疾病的防治,GB是银杏叶提取物的主要有效成分之一。本研究观察到,GB可改善AD模型大鼠学习记忆力下降,提高突触相关蛋白SynapsinⅠ表达,这些作用可能与GB降低海马神经元自噬水平有
[20] 王永炎, 谢颖桢. 化痰通腑法治疗中风病痰热腑实证的源流及发展(二)——化痰通腑法治疗后的不同证候演变及疾病转归与治疗[J]. 北京中医药大学学报:中医临床版, 2013,20(2):1-3.
[21] 王纶. 明医杂著[M].薛己注.王新华,点校. 南京: 江苏科学技术出版社,1995:130.
[22] 朱震亨.丹溪心法[M].赵建新, 点校. 北京: 人民军医出版社,2007:12.
[23] 涂佳玉, 阿基业, 王广基, 等. 通塞脉微丸干预缺血性中风模型大鼠的血浆代谢组学研究[J]. 中国中药杂志, 2012,37(7):1028-1033.
[24] 张允岭, 张锦, 扈新刚, 等. 制作缺血性中风气虚血瘀证大鼠模型的实验研究[J]. 中国中西医结合杂志, 2009,29(4):343-346.
[25] 朱震亨.丹溪心法[M].赵建新,点校. 北京: 人民军医出版社,2007:325.
[26] 吴立本.女科切要[M].佘德友, 点校. 北京: 中医古籍出版社,1999:20.
[27] 杨正望, 周芳, 谈珍瑜, 等. 不同造模方法对大鼠多囊卵巢综合征模型的影响[J]. 中国实验动物学报, 2010,18(1):13-16.
[28] 王瑞杰, 李晖, 邱方, 等. 补肾活血化痰汤对多囊卵巢综合征模型大鼠卵巢血供及形态的影响[J]. 中国药房, 2017,28(4):479-482.
[29] 田硕, 白明, 苗明三. 基于围绝经期综合征临床病症特点的动物模型分析[J]. 中国实验方剂学杂志, 2015,21(19):217-221.
[30] 杂 证谟选读编写点校组.景岳全书·杂证谟选读[M]. 重庆: 重庆大学出版社,1988:141.
[31] 鞠大宏, 张春英, 王安民, 等. 左归丸对去卵巢所致大鼠骨质疏松症的治疗作用[J]. 中国中医基础医学杂志, 2001,7(3):17-20.
[32] 王景, 李小梅, 洪苑. 脂代谢及血清脂联素水平与绝经后骨质疏松症的相关性研究[J]. 实用妇产科杂志, 2014,30(11):828-830.
[33] 成龙, 申竹芳, 孙桂波, 等. 糖尿病动物模型研究进展及在中药研究中的应用[J]. 药学学报, 2015,50(8):951-958.
[34] 戴红, 邹小娟, 赵厚睿, 等. 不同STZ剂量造模对2型糖尿病动物模型证候属性影响的研究[J]. 时珍国医国药, 2015,26(3):751-755.
[35] 陈婧, 易玮, 许能贵, 等. OLETF大鼠自发性2型糖尿病动物模型研究概述[J]. 广州中医药大学学报, 2015,32(1):178-182.
[36] 熊兴江, 王阶. 从痰论治糖尿病[J]. 中国中医基础医学杂志, 2008,14(12):923-924.
[37] 田思胜, 等.朱丹溪医学全书[M]. 北京: 中国中医药出版社,2006:72.
[38] 石变, 袁秀霞, 秦志丰, 等. 论痰浊内阻与肿瘤发生[J]. 中华中医药杂志, 2012,27(9):2389-2393.
[39] 王文萍, 王垂杰. 肿瘤转移的“痰毒流注”理论形成基础及实践意义[J]. 中国中医基础医学杂志, 2002,8(5):4-6.
[40] 郭蕾, 王永炎, 张俊龙, 等. 浊邪在动脉粥样硬化疾病中的病机学意义[J]. 世界中西医结合杂志, 2012,7(2):163-165.
[41] 齐锡友, 董致郅, 谢春荣. 从病因病机谈颈动脉粥样硬化与痰浊的关系[J]. 北京中医药, 2012,31(6):480-481.
[42] 覃光辉, 王骁, 杨亚魁, 等. 从痰论治高黏滞血症[J]. 辽宁中医药大学学报, 2008,10(2):84-85.
[43] 周明学, 徐浩, 陈可冀, 等. 活血解毒中药有效部位对ApoE基因敲除小鼠血脂和动脉粥样硬化斑块炎症反应的影响[J]. 中国中西医结合杂志, 2008,28(2):126-130.
[44] 衡先培, 黄苏萍, 程心玲, 等. 丹栝方干预糖尿病动脉粥样硬化大鼠糖脂代谢及氧化应激研究[J]. 中国中西医结合杂志, 2013,33(2):244-251.
[45] 贾明贤, 余婕, 张媛, 等. 冠心病动物模型建立研究[J]. 世界科学技术:中医药现代化, 2013,15(8):1735-1740.
[46] 任毅, 陈可冀, 张敏州, 等. 405例冠心病患者冠状动脉造影结果与中医证型的相关性[J]. 中医杂志, 2010,51(8):725-728.
[47] 刘建勋, 林成仁, 任建勋, 等. 小型猪痰瘀互结证冠心病“痰、毒、瘀”病机演变规律的实验研究[J]. 中国中药杂志, 2013,38(23):4138-4143.
猜你喜欢
全科护理(2022年10期)2022-12-26
中国合理用药探索(2022年1期)2022-11-26
作文周刊·小学二年级版(2022年20期)2022-05-05
世界科学技术-中医药现代化(2021年5期)2021-11-05
世界科学技术-中医药现代化(2021年12期)2021-04-19
乡村科技(2021年33期)2021-03-16
国际放射医学核医学杂志(2021年10期)2021-02-28
世界科学技术-中医药现代化(2020年2期)2020-07-25
创新作文(小学版)(2019年10期)2019-09-25
中外玩具制造(2013年8期)2013-11-25
