
桑树APX基因的克隆及在拟南芥的遗传转化研究
2017-02-25 06:12:53林天宝陈金娥计东风吕志强
浙江农业科学 2017年1期
刘 岩,朱 燕,林天宝,陈金娥,计东风,吕志强
(浙江省农业科学院蚕桑研究所,浙江杭州 310021)
桑树APX基因的克隆及在拟南芥的遗传转化研究
刘 岩,朱 燕,林天宝,陈金娥,计东风,吕志强1*
(浙江省农业科学院蚕桑研究所,浙江杭州 310021)
利用同源克隆、RACE技术,克隆到桑树APX全长为1 207 bp,包含867 bp的开放阅读框,预测其编码蛋白分子量为31.83 ku,等电点8.08,与APX家族蛋白具有较高同源性;通过进一步构建含MaAPX基因的重组农杆菌,再转化拟南芥筛选后,获得了阳性转基因株,为后续探究桑树APX基因功能奠定了基础。
抗坏血酸过氧化物酶;RACE;桑树
逆境胁迫常常刺激植物产生活性氧物质(reactive oxygen species,ROS),包括超氧阴离子()、羟基自由基(OH-)、H2O2与单线态氧等。若未能及时清除,这些ROS便会与蛋白质、核酸、脂类等反应,引起氧化损伤,导致作物产量和品质降低,严重时甚至造成植物死亡[1]。
在长期的适应过程中,植物已形成了一套复杂的抗氧化防御系统,包括抗氧化酶,如超氧化物歧化酶(superoxide dismutase,SOD)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)、过氧化氢酶(catalase,CAT),以及非酶类复合物分子,如抗坏血酸(ascorbic acid,AsA)、甜菜碱、脯氨酸等[2]。其中,APX利用AsA为其电子供体,通过将H2O2催化生成H2O,从而实现对植物内源H2O2的清除[3-6]。目前,已有从不同物种中克隆到APX的研究报道,包括拟南芥[7]、大麦[8]、棉花[9]、甜菜[10]、橡胶树[11]等。不同的研究发现,在盐、低温等胁迫下,拟南芥AtAPX(又称为APX3)转录上调[12];过量表达拟南芥APX发现能增强转基因烟草的耐盐性[13]。可见APX基因的鉴定和功能研究对植物品种选育及遗传改良等方面具有重要意义。
桑树作为我国的一种古老树种,一直以来都是家蚕的饲料,是蚕丝产业的重要物质基础。传统的桑树研究主要围绕蚕丝产业生产而展开。然而近年来的调查发现,桑树对盐碱、干旱和贫瘠等逆境的适应性也较强,可以作为盐碱地绿化及防沙、治沙等生态治理方面的主要经济树种,在陕西黄土高原水土保持、新疆沙漠治理、重庆市石漠化治理、北京沙地治理、广西石漠化治理等生态建设中都发挥重要功效[14]。为此,本研究从桑树中克隆APX家族基因,与其他来源的APX进行生物信息学比对,再将桑树APX基因借助农杆菌系统转化到拟南芥中,筛选阳性转基因株。这深化了人们对APX家族基因的了解,为桑树抗逆品种的选育提供了候选参考基因。
1 材料与方法
1.1 植物材料
桑树品种强桑1号具有高产、广适、耐瘠的优良生产性能[15],由浙江省农业科学院蚕桑研究所提供。拟南芥(Col-0)由课题组保存。
1.2 M aAPX全长基因的克隆
1.2.1 RNA抽提和cDNA合成
按照Tranzol Plant(北京全式金生物公司)试剂盒方法提取桑树叶片总RNA,经逆转录合成cDNA;3’RACE,5’RACE逆转录反应分别参照Clontech Race kit操作说明进行。
1.2.2 引物设计
根据NCBI已经登录的拟南芥(AIR登录号:AT4G35000.1)、烟草(GenBank登录号:XM-0009806131.1)、水稻(GenBank登录号:AY382617.1)、番茄(GenBank登录号:NM_ 001308331.1)、葡萄(GenBank登录号:EU280159.1)、杨树(GenBank登录号:KF309667.1)APX基因的保守区,设计引物F1、R1。再根据扩增获得的保守区cDNA序列设计3’RACE(F2)和5’RACE(R2)引物,最后设计引物扩增全长ORF(F3,R3)序列。引物由上海生工生物工程技术服务公司合成。
F1:5′-ATGCTYCGHYTVGCRTGGCA-3′,R1:5′-AABGCDTCYTCATCCTTWGCA-3′;F2:5′-TGCT AAACAAGGTGCACCACATC-3′,R2:5′-GCACCTT GTTTAGCATCTGGTAGTCGCC-3′;F3:5′-ATGGCT TTTCCGGTGGTGGACGCTG-3′,R3:5′-CTATTTCT TTCTTTTGCGAACTTCGTAG-3′。其中Y=C/T,H= A/T/C,V=G/A/C,R=A/G,B=G/T/C,D= G/A/T,W=A/T。
1.2.3 扩增反应条件
保守区段的PCR程序为:94℃变性4 min;94℃变性30 s,55℃退火30 s,72℃延伸1 min,35个循环;72℃延伸10 m in。3′RACE和5′RACE参照Clontech公司试剂盒说明进行。
1.2.4 序列测定、分析
琼脂糖凝胶电泳分离PCR扩增产物,割胶回收,与pEASY-T1载体链接,转化DH5α感受态细胞,挑选克隆,送由上海生工生物工程技术公司进行测序。
获得序列用Blast比较,分析MaAPX的基因结构信息。再用ORF finder(http://www.ncbi.nlm. nih.gov/gorf/orFig.cgi)预测基因编码框,以及在线软件(http://isoelectric.ovh.org/)分析蛋白分子量和等电点,并通过TMHMM server v.2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)预测分析MaAPX蛋白的二级结构。利用软件DNAMAN 5.2来分析APX蛋白间的进化同源性。
1.3 定量PCR
200 mmol·L-1NaCl每日浇灌,胁迫处理盆栽的强桑1号苗1周后,分别采集胁迫处理组、正常对照组叶片,同上方法抽提RNA进行反转录后,设计引物F4(5′ACTCTCACGGCTCCAACAATG 3′)、R4(5′AGATGTCCCTAAGATGTGGTGC 3′)用于定量扩增MaAPX(产物237 bp)、引物F5(5′TTCCTATCTACGAGGGTTATGC 3′)、R5(5′GT CAAGAGCAATGTAAGCCAAT 3′)和F6(5′CCCTG CTATGTATGTGGCTAT 3′)、R6(5′GCTGTGGTGG TGAAAGAGTAA 3′)分别扩增Maactin基因179 bp和Atactin基因225 bp片段作为内参。扩增结果按2-ΔΔCt方法进行统计分析[16]。
1.4 重组载体的构建
在引物F3、R3的5′端分别添加XhoⅠ和Bam HⅠ位点,经扩增反应后,胶回收产物经XhoⅠ/Bam HⅠ双酶切后,亚克隆至FGC-5941载体中,转化农杆菌GV3101,涂布含卡那霉素和链霉素的双抗YEP培养基,28℃培养48 h后,挑取菌斑进行PCR鉴定。鉴定引物选用特异性的F3、R3引物对,针对菌斑进行菌落PCR鉴定,阳性菌落送测序做进一步的鉴定。
1.5 转基因拟南芥的转化及鉴定
挑取经过菌落PCR验证、含有重组质粒的阳性GV3101克隆,加到含有抗生素的LB液体培养基中,待D600为0.6时,收集菌体;用5%蔗糖溶液、稀释重悬菌体,浸花法转化拟南芥,收取转化植株的种子,在含10μg·mL-1BASTA除草剂的MS培养基上筛选阳性苗,并利用定量PCR方法检测转基因株的MaAPX转录表达水平。
1.6 统计分析
利用Excel和SPSS19 version进行数据的统计分析,试验数据为3次重复的平均值±标准误,采用单因素方差分析和最小显著差异法分析不同数据组间的差异。
2 结果与分析
2.1 全长M aAPX cDNA的获得
利用简并引物对桑叶cDNA样品扩增,克隆获得584 bp长度产物(图1中A),测序结果证实该段序列与NCBI登录的APX家族基因具有高度同源性。据此序列进行3′和5′RACE扩增,进一步获得822 bp和429 bp片段。经拼接得到全长的MaAPX cDNA序列(图1中B),序列全长1 207 bp,含一个完整的开放阅读框867 bp,编码288个氨基酸,编码蛋白包含植物过氧化酶结构域(图1中C),其分子量为31.83 ku;等电点p I 8.08。C端包含一个“AVGVAVAA”典型的富含缬氨酸和丙氨酸的过氧化酶体导肽序列(图1中B),同时,TMHMM的分析显示其包含一个跨膜域(图1中D)。
2.2 桑树APX的进化比较分析
桑树MaAPX蛋白与同类蛋白同源性较高,在分析序列中平均高达75.92%。其中,与陆地棉(Gossypium hirsute,GenBank:NP_001314208.1)来源的APX蛋白同源性最高,达86.8%;其次是烟草(GenBank:XP_009804433.1)、番茄(GenBank:NP_001295260.1),分别高达86.4%和85.4%。相比之下,MaAPX与葡萄(GenBank:EU280159.1)、橡胶树(GenBank:AAO14118.1)、杨树(GenBank:KF309667.1)同源性相对较低,只有64.3%~65.9%。根据APX蛋白在不同物种间的保守性差异,绘制出进化树(图2)。
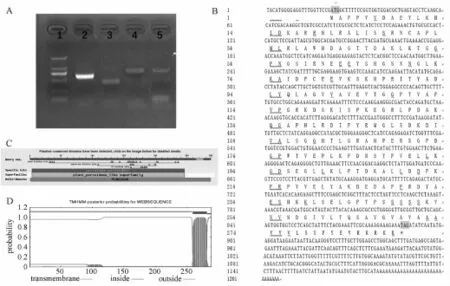
图1 MaAPX的扩增及序列特点
2.3 盐胁迫下桑树MaAPX的转录表达分析
200 mmol·L-1盐胁迫处理后,检测MaAPX在盆栽桑树苗叶片中的转录表达水平发现,与正常未胁迫的对照组桑苗相比,胁迫后叶片中MaAPX的转录表达水平明显上调(P<0.05,图3),证实MaAPX响应盐胁迫的刺激,参与胁迫条件下的植物适应性反应。
2.4 重组载体的构建及农杆菌转化
利用PCR和基因重组技术,将MaAPX片段构建入pFGC5941的Xho I和Bam H I限制性内切酶位点之间,经酶切和测序鉴定后,成功构建获得重组表达载体pFGC-MaAPX(图4中A)。转化GV3101农杆菌,涂布含卡那霉素(50μg·m L-1)和链霉素(50μg·m L-1)的抗性平板筛选,利用引物F3、R3,针对生长的农杆菌菌落进行PCR检测,结果挑选克隆均扩增到特异的MaAPX产物条带(图4中B)。
2.5 不同转基因拟南芥株系间的表达丰度检测
在Basta(10μg·m L-1)抗性的MS培养平板上,转基因植株能正常生长,而未转入基因的植株不能正常萌发。选择抗性筛选下长势良好的幼苗经PCR检测确定阳性后,将获得株系MaAPX移栽入营养土盆钵中。分别单株收转基因株系种子,重复以上抗性筛选过程,获得纯化转基因株系。利用荧光定量PCR检测了MaAPX在转基因株系中的转录表达量(图5)。
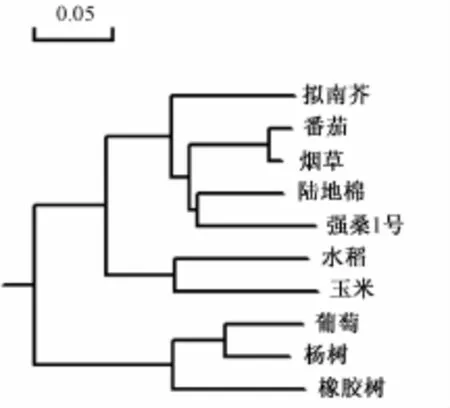
图2 桑树MaAPX与几种已报道APX蛋白的同源性及进化比较
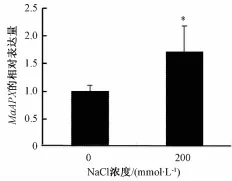
图3 胁迫后MaAPX在桑叶中的转录表达
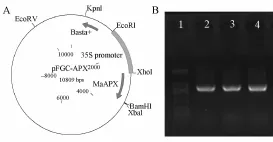
图4 重组pFGCc-MaAPX的构建及在转化农杆菌中的PCR检测
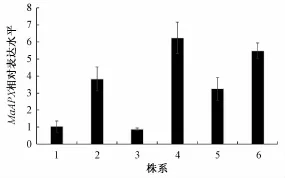
图5 不同拟南芥转基因株系中的MaAPX表达丰度
3 讨论
本研究从目前生产广泛应用的强桑品种中克隆到MaAPX基因,对其进行了一系列的生物信息学分析,发现MaAPX具有保守的植物过氧化物酶体结构域(图1),氨基酸组成上与APX家族蛋白同源性较高(图2)。MaAPX的C段包含序列“AVGVAVAA”即为典型的富含缬氨酸和丙氨酸的过氧化酶体导肽,Mullen等[17]报道含有这种过氧化物酶体定位导肽的酶锚定在过氧化酶体,蛋白分子向外暴露于胞质。推测MaAPX可能也是定位于过氧化酶体的同工酶,与毛白杨相似[18]。虽然同为过氧化酶体定位的APX,桑树APX与陆地棉、烟草等同源性却远高于与毛白杨APX(图2),由于植物体内执行过氧化物酶功能的有一大类家族蛋白,不同APX基因在不同物种间可能表现出不同的作用模式。
H2O2是一种重要的活性氧基团,在植物体内可由多种刺激(如胁迫、激素等)诱导产生。高浓度的H2O2对细胞具有较强的损伤能力,能够氧化细胞内多种重要的酶分子,甚至损伤DNA,对植物细胞造成不可逆的伤害。鉴于APX可以清除逆境产生的自由基,近年来植物耐逆性研究中的一个热点就是向植物中导入APX基因,提高植物体内的APX蛋白活性[19]。Wang等[20]在番茄中过量表达APX证实转基因番茄的抗寒和抗盐能力得到显著提高;Sarowar等[21]将一个辣椒APX基因转入烟草,同样提高了转基因烟草的抗氧化胁迫与抗真菌能力,而许传俊等[22]从蝴蝶兰中克隆了APX基因,研究其在蝴蝶兰体内的时空表达情况及对机械伤害和高盐胁迫的响应。本研究为后续开展桑树APX生理功能及其相关的抗氧化机理研究提供了基础。
[1] BADAW IG H,KAWANO N,YAMAUCH I Y,et al.Overexpression of ascorbate peroxidase in tobacco chloroplasts enhances the tolerance to salt stress and water deficit[J]. Physiologia Plantarum,2004,121(2):231-238.
[2] ASADA K.The water-water cycle in chloroplasts:Scavenging of active oxygens and dissipation of excess photons[J].Annual Review of Plant Physiology and Plant Molecular Biology,1999,50:601-639.
[3] ASADA K.Production and scavenging of reactive oxygen species in chloroplasts and their functions[J].Plant Physiology,2006,141(2):391-396.
[4] LU Z,LIU D,LIU S.Two rice cytosolic ascorbate peroxidases differentially improve salt tolerance in transgenic Arabidopsis[J].Plant Cell Reports,2007,26(10):1909-1917.
[5] GILL S S,TUTEJA N.Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in cropplants[J].Plant Physiology&Biochem istry,2010,48(12):909-930.
[6] SH IGEOKA S,ISHIKAWA T,TAMOI M,et al.Regu lation and function of ascorbate peroxidase isoenzymes[J].Journal of Experimental Botany,2002,53(372):1305-1319.
[7] ZHANG H,WANG J,NICKEL U,et al.Cloning and expression of an Arabidopsis gene encoding a putative peroxisomal ascorbate peroxidase[J].Plant Molecular Biology,1997,34(6):967-971.
[8] SH IW M,MURAMOTO Y,UEDA A,et al.Cloning of peroxisomal ascorbate peroxidase gene from barley and enhanced thermos tolerance by overexpressing in Arabidopsis thaliana[J].Gene,2001,273(1):23-27.
[9] SHEN G,KUPPU S,VENKATARAMANI S,et al.Ankyrin repeat-containing protein 2A is an essentialmolecular chaperone for peroxisomal membrane-bound ascorbate peroxidase3 in Arabidopsis[J].Plan t Cell,2010,22(3):811-831.
[10] DUNAJSKA-ORDAK K,SKORUPA-KŁAPUTM,KURNIK K,et al.Cloning and expression analysis of a gene encoding for ascorbate peroxidase and responsive to salt stress in beet(Beta υulgaris)[J].Plant Molecular Biology Reporter,2014,32(1):162-175.
[11] CHAO J,ZHANG S,CHEN Y,et al.Cloning,heterologous exp ression and characterization of ascorbate peroxidase(APX)gene in laticifer cells of rubber tree(Heυea brasiliensis Muell. Arg.)[J].Plant Physiology&Biochemistry,2015,97:331-338.
[12] ZHANG H,WANG J,NICKEL U,et al.Cloning and expression of an Arabidopsis gene encoding a putative peroxisomal ascorbate peroxidase[J].Plant Molecular Biology,1997,34(6):967-971.
[13] BADAW I GH,KAWANO N,YAMAUCH I Y,et al.Overexpression of ascorbate peroxidase in tobacco chloroplasts enhances the tolerance to salt stress and water deficit[J]. Physiologia Plantarum,2004,121(2):231-238.
[14] VIJAYAN K,SRIVASTAVAA P P,RAGHUNATHB M K,et al.Enhancement of stress tolerance in mulberry[J].Scientia Horticulturae,2011,129:511-519.
[15] 吕志强,计东风,周勤,等.桑树新品种强桑1号的选育[J].蚕业科学,2011,37(1):9-12.
[16] LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-DeltaDeltaC(T))method[J].Methods,2001,25(4):402-408.
[17] MULLEN R T,TRELEASE R N.The sorting signals for peroxisomalmembrane-bound ascorbate peroxidase are within its C-terminal tail[J].Journal of Biological Chem istry,2000,275(21):16337-16344.
[18] 潘翔,李娜,李印军,等.毛白杨抗坏血酸过氧化物酶基因PtAPX2的克隆表达及分析[J].北京林业大学学报,2013,35(1):36-44.
[19] ISHIKAWA T,SAKAI K,YOSHIMURA K,et al.cDNAs encoding sp inach stromal and thylakoid-bound ascorbate peroxidase,differing in the presence or absence of their 3’-coding regions[J].FEBS Letters,1996,384(3):289-293.
[20] WANG Y J,W ISNIEWSKI M,MEILAN R,et al. Overexpression of cytosolic ascorbate peroxidase in tomato con fers tolerance to chilling and salt stress[J].Journal of American Society for Horticultural Science,2005,130(2):167-173.
[21] SAROWAR S,KIM E N,KIM Y J,et al.Overexpression of a pepper ascorbate p roxidase-like 1 gene in tobacco plants enhances tolerance to oxidative stress and pathogens[J].Plant Science,2005,169(1):55-63.
[22] 许传俊,孙叙卓,李玲,等.蝴蝶兰抗坏血酸过氧化物酶基因克隆及其表达研究[J].园艺学报,2012,39(4):769-776.
(责任编辑:张 韵)
S888
A
0528-9017(2017)01-0155-05
文献著录格式:刘岩,朱燕,林天宝,等.桑树APX基因的克隆及在拟南芥的遗传转化研究[J].浙江农业科学,2017,58(1):155-159.
10.16178/j.issn.0528-9017.20170149
2016-08-23
现代农业产业技术体系建设专项(蚕桑)(CARS-22-ZJ0105);浙江省农业新品种选育重大科技专项
刘 岩(1976-),女,河南焦作人,副研究员,从事桑蚕分子生物学研究工作,E-mail:lymorus@gmail.com。
吕志强(1965-),研究员,从事桑树育种学研究工作,E-mail:1398131715@139.com。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
学与玩(2022年10期)2022-11-23 08:32:00
今日农业(2022年3期)2022-06-05 07:12:08
音乐教育与创作(2022年1期)2022-04-26 02:21:20
今日农业(2020年16期)2020-12-14 15:04:59
生命科学研究(2018年1期)2018-05-29 01:12:47
小学生作文(中高年级适用)(2018年3期)2018-04-18 01:24:40
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
学生天地(2016年16期)2016-05-17 05:46:06
