
低钾胁迫对甘薯块根淀粉理化特性的影响及其基因型差异
2017-02-16 03:09:01唐忠厚张爱君陈晓光靳容刘明李洪民丁艳锋
中国农业科学 2017年3期
唐忠厚,张爱君,陈晓光,靳容,刘明,李洪民,丁艳锋
(1江苏徐州甘薯研究中心/农业部甘薯生物学与遗传育种重点实验室,江苏徐州 221131;2南京农业大学农学院,南京 210095)
低钾胁迫对甘薯块根淀粉理化特性的影响及其基因型差异
唐忠厚1,2,张爱君1,陈晓光1,靳容1,刘明1,李洪民1,丁艳锋2
(1江苏徐州甘薯研究中心/农业部甘薯生物学与遗传育种重点实验室,江苏徐州 221131;2南京农业大学农学院,南京 210095)
【目的】甘薯[Ipomoea batatas(L.) Lam]属块根类淀粉作物,同时也是较典型“喜钾”作物,而甘薯种植区土壤缺钾十分普遍。同时,甘薯块根淀粉用途与其理化特性密切相关,因此,必须加强研究钾缺乏对甘薯块根淀粉理化特性的影响。【方法】选用3个不同钾效率型甘薯徐薯32(耐低钾与钾高效型)、徐薯18(中间型)与宁紫薯1号(不耐低钾与钾低效型)为材料,利用1980年设置的长期肥料定位试验田低钾(LK,氮磷处理)与正常钾水平(CK,氮磷钾处理)两处理,分析甘薯块根淀粉粒径大小与分布、热焓特性、糊化特性、吸湿性及膨胀力等特征指标差异。【结果】与正常钾相比,低钾胁迫改变甘薯块根中直链淀粉、支链淀粉、总磷、总脂肪和氮等主要组分,不同钾效率型甘薯间变化存在差异。甘薯淀粉粒径存在≤2.50 μm、2.50—5.00 μm和5.00—30.00 μm等明显的3峰分布,低钾胁迫下淀粉粒径及其分布均发生变化,淀粉平均粒径降低,其中在体积分布中,徐薯32平均粒径减幅最小,宁紫薯1号降幅达显著水平(P<0.05),表面积与数目分布中差异不显著,而不同粒径的百分比、平均粒径与峰值(数目分布中无峰值)均有不同的变化,且这些变化在不同品种间也存在差异。淀粉糊化过程中的起始温度(T0)、峰值温度(Tp)和热焓值(ΔH)普遍低于正常钾供应处理的相应指标,尤其宁紫薯1号在3个热特征值上的差异均达显著水平,其差异幅度大于徐薯32,3个甘薯品种最高粘度值、崩解值均明显下降,徐薯18与宁紫薯1号的最低粘度值、最终粘度值受低钾影响显著上升,而徐薯32的表现则相反。块根淀粉吸湿性均不同程度降低,徐薯18与宁紫薯1号降低达显著水平(P<0.05),徐薯32淀粉吸湿性变化较小,但其吸湿性值最高。淀粉膨胀力不同程度上升,宁紫薯1号增幅达显著水平(P<0.05),徐薯32淀粉颗粒膨胀力较高,受低钾胁迫影响效应小。相关分析表明,甘薯淀粉主要理化指标间密切相关,低钾胁迫改变甘薯块根淀粉组分及颗粒粒径大小,是导致不同程度影响甘薯块根淀粉的糊化特性、热特性等理化特性的主要因素。【结论】钾是影响甘薯块根淀粉理化特性的重要元素,低钾胁迫对甘薯块根淀粉组分及其品质理化特性产生较明显影响,不同钾效率型甘薯块根淀粉特性对低钾胁迫响应不同。
甘薯;低钾胁迫;块根;淀粉;理化特性
0 引言
【研究意义】甘薯(Ipomoea batatas(L.) Lam)属淀粉类块根作物,同时也是较典型“喜钾”作物,土壤钾供应状况与甘薯生长发育及产量品质形成等过程均存在密切联系[1]。在中国甘薯种植区,尤其长江中下游薯区与南方薯区,土壤钾缺乏(低钾)现象较普遍,已成为制约甘薯产量提高和品质改善的主要因素之一。加之中国钾肥资源较短缺,农作物上钾素的大量使用也是一种普遍现象,因此,利用不同钾效率型甘薯材料,探讨低钾胁迫下甘薯块根的淀粉理化特性变异,可为甘薯块根淀粉品质改良及钾营养利用研究提供理论依据。【前人研究进展】甘薯块根品质性状本身存在基因型差异[2-3],甘薯块根品质的基因型效应、钾肥水平效应以及基因型与钾肥水平互作效应均达到极显著水平[2]。钾营养影响甘薯生长的生理基础,对甘薯生长发育与品质形成至关重要[4];施钾降低甘薯块根干率、淀粉率[5-6]和蛋白质含量[7],显著降低甘薯块根食味总评、淀粉最高粘度、崩解值和回复值[2],提高薯块鲜产、胡萝卜素与花青素[8]以及粗纤维含量[7]。淀粉是甘薯块根的主要组分,其理化特性直接决定主要用途[9],研究表明,甘薯淀粉膨润力、颗粒大小及支直比等理化特性对粉丝品质影响显著[10]。甘薯淀粉的理化特性主要受遗传因素控制[11],但栽培手段或外界环境同样影响淀粉的理化特性,如土温影响甘薯直链淀粉含量、糊化特性、粒径大小、支链淀粉链长分布及DSC特性等理化特性的形成[12],施钾量与施肥时期也是影响甘薯淀粉产量与糊化特性的重要因素[13-14]。【本研究切入点】以往研究多从基因型差异[15]、环境效应[2]、栽培[16]以及生物技术手段[17]等方面探讨甘薯块根淀粉理化特性和功能特征;钾缺乏影响甘薯前期植株的生长和养分吸收[18-19],然而低钾胁迫对甘薯块根淀粉理化特性的影响,以及不同钾效率型甘薯材料淀粉特性对缺钾胁迫的表现差异,迄今尚未见相关报道。【拟解决的关键问题】本研究以 3个不同钾效率型甘薯徐薯 18、徐薯 32与宁紫薯1号为材料,设置长期肥料定位试验田低钾(施氮磷肥)与正常钾水平(施氮磷钾肥)两处理,旨在从甘薯块根淀粉粒径大小与分布、热焓特性、糊化特性、吸湿性及与膨胀力等方面探讨低钾胁迫下不同钾利用效率基因型甘薯块根淀粉的主要理化特性差异,以期为甘薯淀粉品质改良及钾营养利用提供理论依据。
1 材料与方法
1.1 材料
试验材料选用徐薯32(耐低钾与钾高效品种,黄肉色,短蔓、深绿叶色,且熟食口味佳)、宁紫薯1号(不耐低钾与钾低效品种,紫肉色,熟食口味佳)和徐薯18(中间类型品种,白肉色,国内主栽品种)[1]。
1.2 试验设计与样品处理
1.2.1 试验设计 2011—2012年试验在江苏徐州甘薯研究中心 1980年秋播开始设置的长期定位试验田(北纬34°27′,东经117°29′)中进行,土壤为黄潮土,质地砂壤。试验前2处理土壤的主要养分含量如下,施氮磷钾肥处理(简称施钾处理):有效钾 105.10 mg·kg-1,有机质含量10.89 g·kg-1,全氮1.12 g·kg-1,碱解氮81.00 mg·kg-1,全磷0.89 g·kg-1,有效磷15.00 mg·kg-1;仅施氮磷肥处理(简称不施钾处理):有效钾46.10 mg·kg-1,有机质含量11.38 g·kg-1,全氮1.10 g·kg-1,碱解氮78.50 mg·kg-1,全磷0.74 g·kg-1,有效磷15.50 mg·kg-1。
本试验共2个处理:(1)施钾处理(对照CK);(2)不施钾处理(低钾处理,Low potassium,LK),每处理4次重复。每季肥料种类与用量:氮肥为尿素,用量150 kg N·hm-2;磷肥为过磷酸钙,用量75 kg P2O5·hm-2;钾肥为硫酸钾,用量为112.5 kg K2O·hm-2,起垄时肥料一次施入。小麦-甘薯轮作,平茬收割,人工翻地(20 cm深),小区面积33.3 m2,每小区6行,垄宽85 cm,高25 cm。6月中旬扦插,苗长25—30 cm,每小区155株,10月下旬收获;田间管理同一般甘薯高产栽培。
1.2.2 取样与样品处理 取样:120 d挖根,选择健壮、无病害、重约100—300 g鲜甘薯6块,洗净。淀粉制备:去皮,切成细块状,放入高速打浆机,加适量水,粉碎30 s,倒入100目纱袋,加0.5 L水洗提,另0.5 L水再洗提一次,将洗提液合并,过100目筛,静置12 h,置50℃烘箱24 h,研磨,过100目筛,封存。测定前用无水乙醇洗涤,离心,0.5 mol·L-1NaOH振荡,离心,除去可溶性糖、蛋白等,45℃烘干至衡重,沉淀物即为纯淀粉,用于淀粉化学组分及特性分析。
1.3 测定内容与方法
1.3.1 淀粉主要化学组分 直链淀粉含量:参照唐忠厚等[20]方法;总磷测定:参照GB/T 5009.87—2003食品中磷的测定;总脂肪测定:参照GB/T 5512—85,索氏提取法测定;氮测定:参照GB/T 5511—85,采用半微量凯氏定氮法测定。
1.3.2 淀粉粒径大小与分布 采用 Saturn Digisizer 5200激光衍射粒度分析仪(Micromeritics公司,美国)测定淀粉粒粒级分布及粒径大小。称取50 mg淀粉样品,加入5 mL去离子水,置于超声波中水浴振荡2—3 min至充分分散,得待测水悬液。仪器测定参数如下:流速为12 mL·min-1,运行时间为60 s;待仪器预热(光路系统温度稳定)后,运行背景测定操作(仪器调零),然后注入一定体积待测液,遮光率(20±1)%且稳定后开始分析。每个样品重复测定2次,取其平均值作为最终测定值。
1.3.3 淀粉热焓分析 采用DSC1差示扫描量热分析仪(Mettler Toledo公司)测定。具体步骤:精确称取2.5 mg左右已知水分的淀粉样品,加入7.5 mL的去离子水于铝质样品盘中,摇均,立即将样品池压紧密封,放入仪器内的样品座,用密封空的铝盒参比盘作参比物,通氮气;测定结束,曲线自动储存到数据库中,采用STARe Evaluation 13.0(Mettler)软件处理,获得吸热曲线上的起始温度(onset temperature,T0)、峰值温度(peak temperature,TP)、终止温度(conclusion temperature,Tc)和热焓变化(enthalpy of transition,ΔH),每个样品重复3次。条件:升温范围50—120℃,升温速率10℃·min-1,通氮气速率30 mL·min-1。
1.3.4 RVA特性测定 利用 Tech-master RVA仪(Perten公司)测定,参考唐忠厚[13]等方法。测定指标包括最高粘度(PKV)、热浆粘度(HPV)、最终粘度(CPV)、崩解值 (BDV) 、回复值(CSV)和糊化温度(PT)等。
1.3.5 吸湿性测定 准确称取经烘干、研磨、过筛(100目)后50 mg淀粉试样进行吸湿性测定,加10 mL纯水,摇均,密封,静置12 h,离心(6 000 g/min,15 min),弃去上清液,取出沉淀物G1,精确称量,然后 45℃烘干,达恒重后,读取干物质的重量 G2;按下式计算吸湿能力:吸湿能力=(烘干前重量G1-烘干后重量G2)/烘干后重量G2。
1.3.6 膨胀力测定 参照RASPER[21]方法,称取0.5 g淀粉(干基)加入50 mL去离子水,振荡均匀后,在85℃的水浴中振荡糊化30 min,倒入100 mL量筒中,静置24 h,测定沉淀固体物所占体积,以每g淀粉的体积表示淀粉的膨胀力。
1.4 数据统计与分析
试验数据为2年均值,采用 Sigmaplot13作图和SPSS13.0 统计分析与处理数据。
2 结果
2.1 低钾胁迫对甘薯块根淀粉主要化学组分的影响及其差异
甘薯淀粉颗粒主要由直链淀粉和支链淀粉组成,还含少量脂肪、磷、蛋白、灰分和水等[11]。由表 1可知,3个甘薯材料间表观直链淀粉与支链淀粉含量差异显著(P<0.05);低钾下徐薯18、宁紫薯1号直链淀粉含量降低,徐薯32直链淀粉含量增加,支链淀粉含量变化正相反,但处理间变幅差异均未达显著水平。低钾胁迫显著增加淀粉中磷含量(P<0.05),两处理间宁紫薯1号块根淀粉磷含量均最高,与徐薯18、徐薯32含量差异达显著水平(P<0.05)。低钾胁迫抑制脂肪形成,但降幅均不显著,低钾胁迫下材料间脂肪含量差异达显著水平(P<0.05)。材料间块根淀粉中氮含量差异不显著,低钾胁迫降低块根淀粉中氮含量,徐薯32降低幅度最大,但差异未达显著水平。
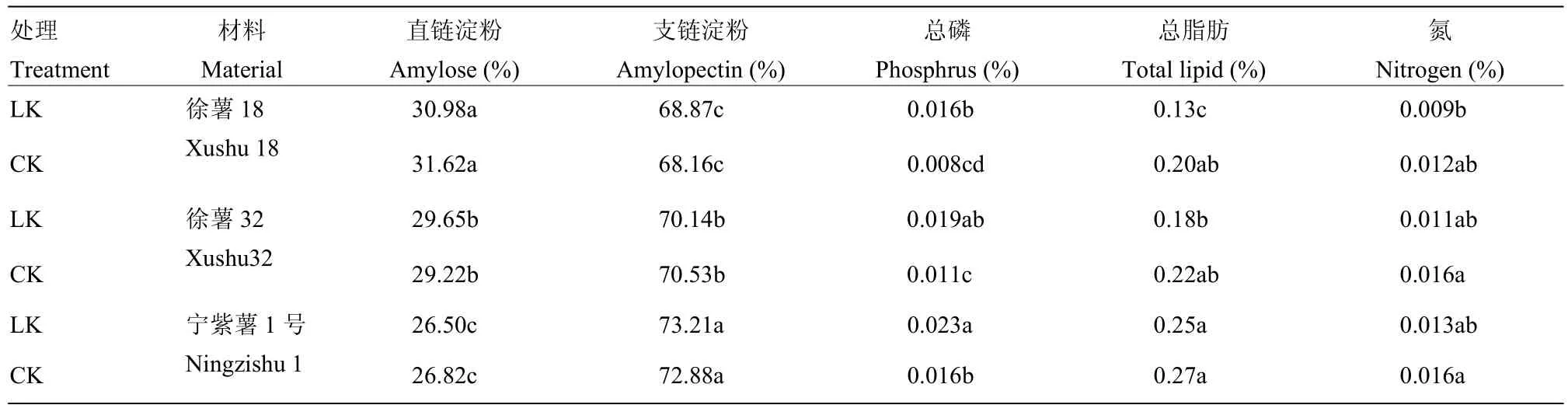
表1 低钾胁迫对甘薯材料块根淀粉主要组分差异的影响Table 1 Influence of LK stress on the difference of major compositions of three kinds of sweetpotato storage root starch
2.2 低钾胁迫对甘薯块根淀粉粒径大小与分布的影响及其差异
2.2.1 甘薯块根淀粉粒径与体积分布比较分析 在淀粉类作物的贮藏器官中,淀粉粒大小、粒度分布等是影响淀粉功能特性的重要因素之一[21]。徐薯 18、徐薯32与宁紫薯1号等3个材料在低钾胁迫与CK处理下淀粉粒的体积分布如图1所示,3个材料的淀粉粒径范围分别为0.47—29.72、0.47—28.06和0.47—30.32 μm;体积分布均呈现为三峰,第3峰高于前2个峰;根据峰谷值界限,将淀粉粒径大小划分 3个级别,分别为≤2.50、2.50—5.00和5.00—30.00 μm,其中80%以上粒径在5.00—30.00 μm,2.50—5.00 μm的比例最小,表明大淀粉颗粒占淀粉主要体积。由表2可知,3个材料不同处理下全部平均粒径范围12.80—16.25 μm,徐薯32全部平均粒径最小,与徐薯18、宁紫薯1号差异显著(P<0.05);低钾胁迫下淀粉全部平均粒径均减少,受粒径≤2.50 μm的淀粉粒平均粒径上升影响,徐薯32减幅最小,而宁紫薯1号降幅达显著水平(P<0.05)。对比CK处理,低钾胁迫下3个材料淀粉的不同粒径颗粒体积分布发生变化。粒径≤2.50 μm的淀粉粒比例均有所增加,徐薯32增幅最大,达16.91%;平均粒径变化不一,徐薯18与宁紫薯1号降低,徐薯32增加;百分比峰值均有增加。粒径2.50—5.00 μm的淀粉粒百分比占整个比例最小,徐薯 32的比例最高,与徐薯18、宁紫薯1号间差异显著(P<0.05);低钾胁迫下徐薯 32与宁紫薯 1号的粒径 2.50—5.00 μm百分比增加,徐薯32增幅达32.03%;平均粒径变化不一,徐薯18与宁紫薯1号增加,徐薯32降低;百分比峰值均有增加。粒径5.00—30.00 μm的淀粉粒百分比占整个比例最大,徐薯32百分比与平均粒径最低,与徐薯18、宁紫薯1号间差异显著(P<0.05);而峰值差异不显著;低钾胁迫下,徐薯32的粒径5.00—30.00 μm百分比比例降低较大,但未达显著水平,徐薯18与宁紫薯1号变化较小。
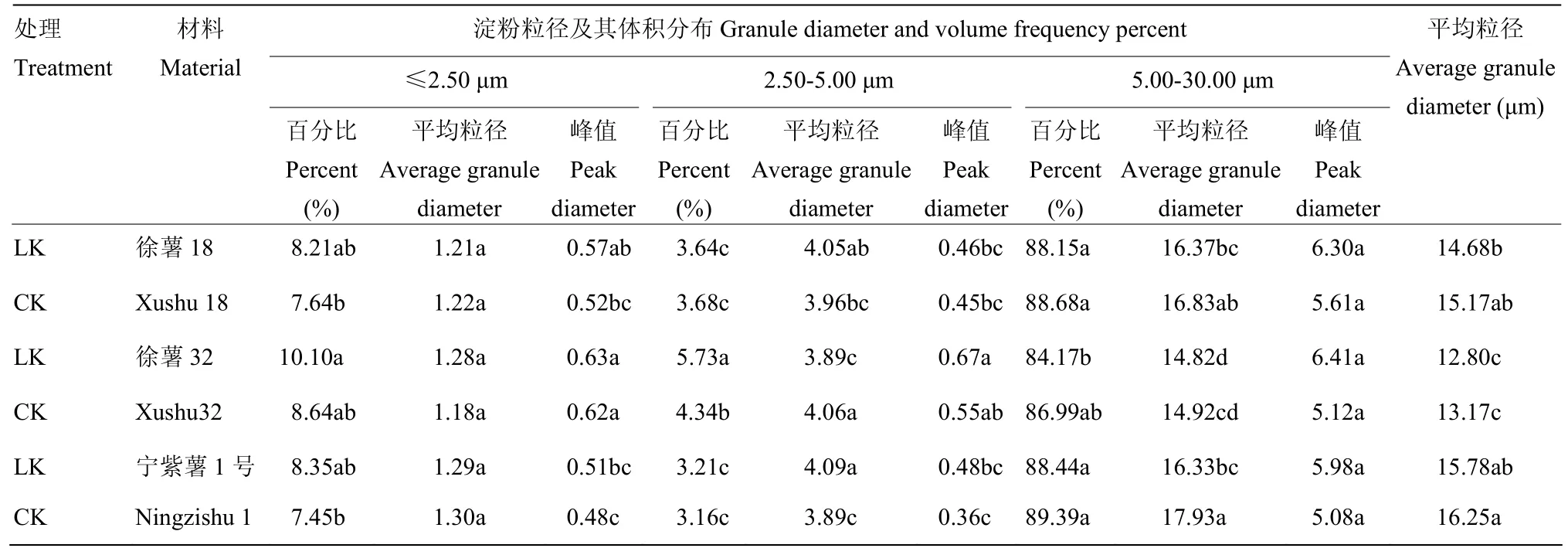
表2 低钾胁迫对甘薯材料块根淀粉粒径及其体积分布的影响Table 2 Granule diameter and volume frequency percent of three kinds of sweetpotato storage root starch under low K stress
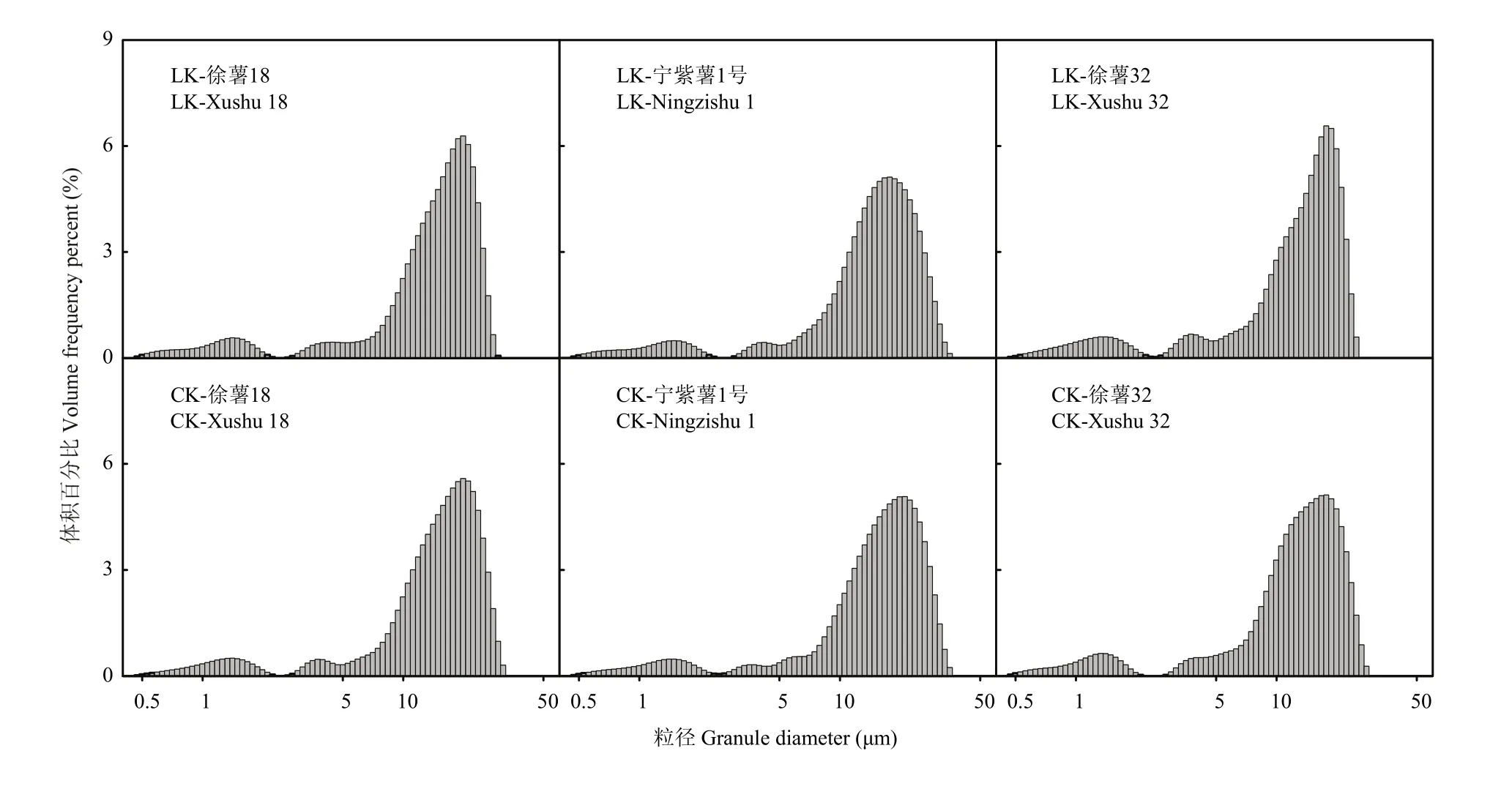
图1 低钾胁迫下甘薯材料块根淀粉粒径体积分布图Fig. 1 Starch granule distribution of three kinds of sweetpotato storage root starch under low K stress (by volume)
2.2.2 甘薯块根淀粉粒径与表面积分布比较分析徐薯18、徐薯32与宁紫薯1号等3个材料在低钾胁迫与CK处理下淀粉粒的表面积分布如图2所示,甘薯淀粉表面积分布主要呈三峰,以2.50 μm和5.00 μm为三峰分界,比较发现,第1峰和第2峰间区分明显,淀粉中第1峰和第2峰的比例与峰值均大于其第3峰;50%左右粒径小于2.50 μm,粒径2.50—5.00 μm比例最小。由表3可知,3个材料不同处理下全部平均粒径范围为5.86—7.53 μm,徐薯32全部平均粒径最小,与徐薯18、宁紫薯1号差异显著(P<0.05);低钾胁迫下淀粉全部平均粒径均减少,但未达显著水平。
对比CK处理,低钾胁迫下3个材料淀粉的不同粒径颗粒表面积分布发生变化。粒径≤2.50 μm的淀粉粒百分比例均有增加,徐薯18增幅最大,达4.38%;平均粒径变化不一,徐薯18与宁紫薯1号略降,徐薯32略增;徐薯32百分比峰值下降明显,降幅达8.28%,徐薯18与宁紫薯1号百分比峰值增加,其中宁紫薯1号增幅16.72%。粒径2.50—5.00 μm的淀粉粒百分比占整个比例最小;低钾胁迫下3个材料间百分比差异明显,徐薯18与宁紫薯1号均减少,徐薯32比例增加,且增幅较大,达29.11%;徐薯18与宁紫薯1号平均粒径增加,徐薯32略减,百分比峰值均上升,且徐薯32的平均粒径和峰值与徐薯18、宁紫薯1号间差异达显著水平(P<0.05)。粒径5.00—30.00 μm的淀粉粒百分比占整个比例40%左右,徐薯32的百分比与平均粒径最低,其平均粒径与徐薯18、宁紫薯1号间差异显著(P<0.05);低钾胁迫下,粒径 5.00—30.00 μm百分比均降低,徐薯32降幅最大,达显著水平(P<0.05),徐薯18与宁紫薯1号差异不明显;徐薯18与宁紫薯1号平均粒径下降,宁紫薯1号达显著水平(P<0.05),而徐薯32小幅增加;钾对3个材料间峰值影响有限,变化差异均未显著。
2.2.3 甘薯块根淀粉粒径与数目分布比较分析 徐薯18、徐薯32与宁紫薯1号等3个材料在低钾胁迫与CK处理下淀粉粒的数目分布如图3所示,甘薯淀粉数目分布呈单峰,98%以上为粒径≤2.50 μm,粒径5.00—30.00 μm比例最小,表明小淀粉颗粒所占体积小,但数目最大。由表4可知,3个材料不同处理下全部平均粒径范围0.86—0.92 μm,徐薯32全部平均粒径最高,但材料间与处理间差异均未达显著水平。
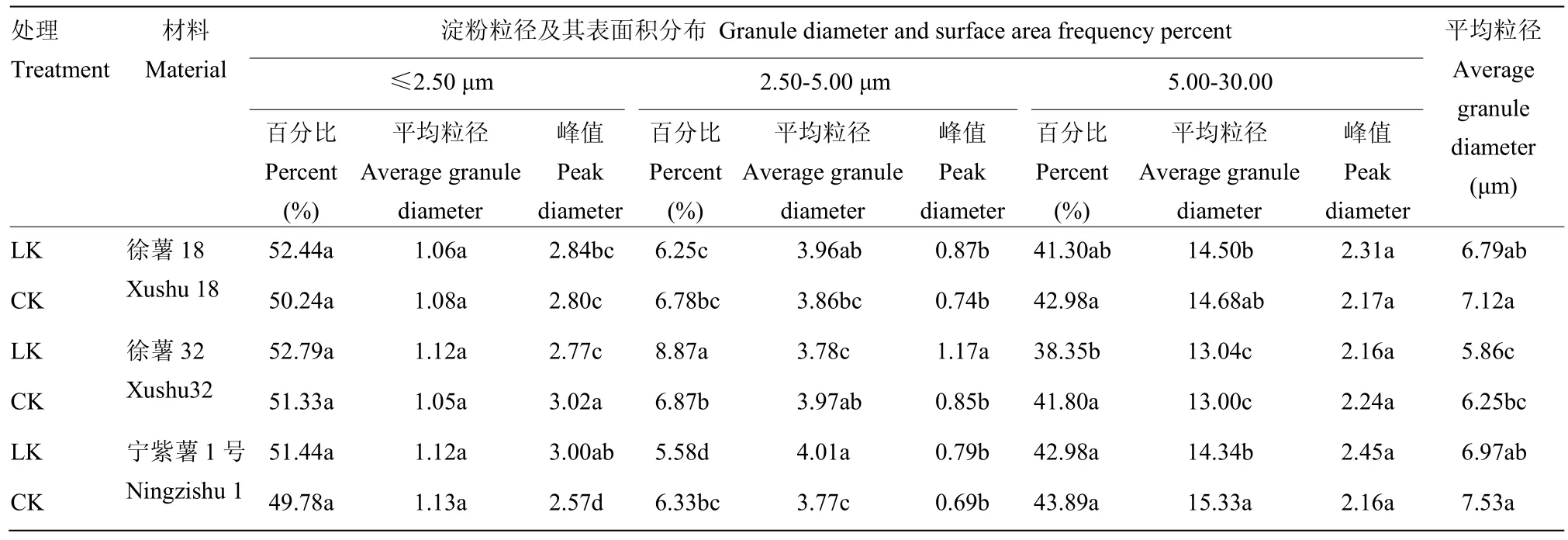
表3 不同处理下3个甘薯材料块根淀粉粒径及其表面积分布Table 3 Granule diameter and surface area frequency percent of three kinds of sweetpotato storage root starch under low K and CK treatment (by surface area)
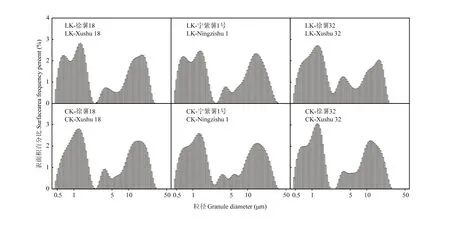
图2 低钾胁迫下甘薯材料块根淀粉粒径表面积分布图Fig. 2 Starch granule distribution of three kinds of sweetpotato storage root starch under low K treatment (by surface area)
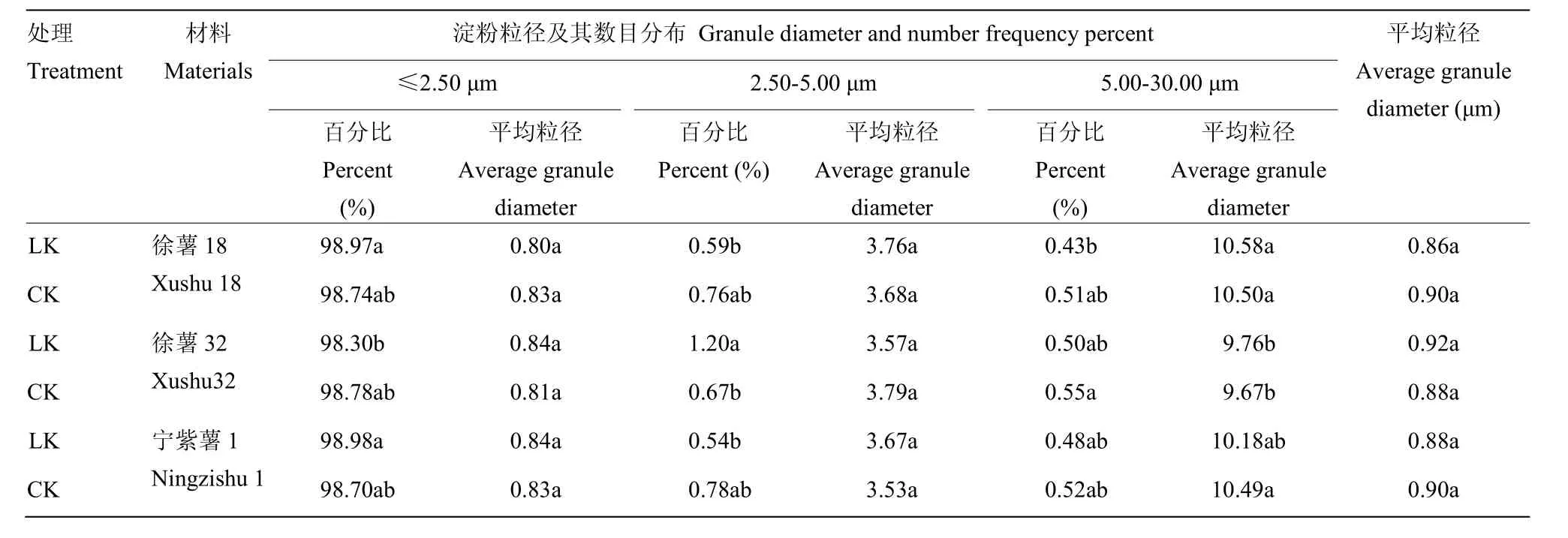
表4 低钾胁迫对甘薯材料块根淀粉粒径及其数目分布的影响Table 4 Granule diameter and number frequency percent of three kinds of sweetpotato storage root starch under low K stress (by number)
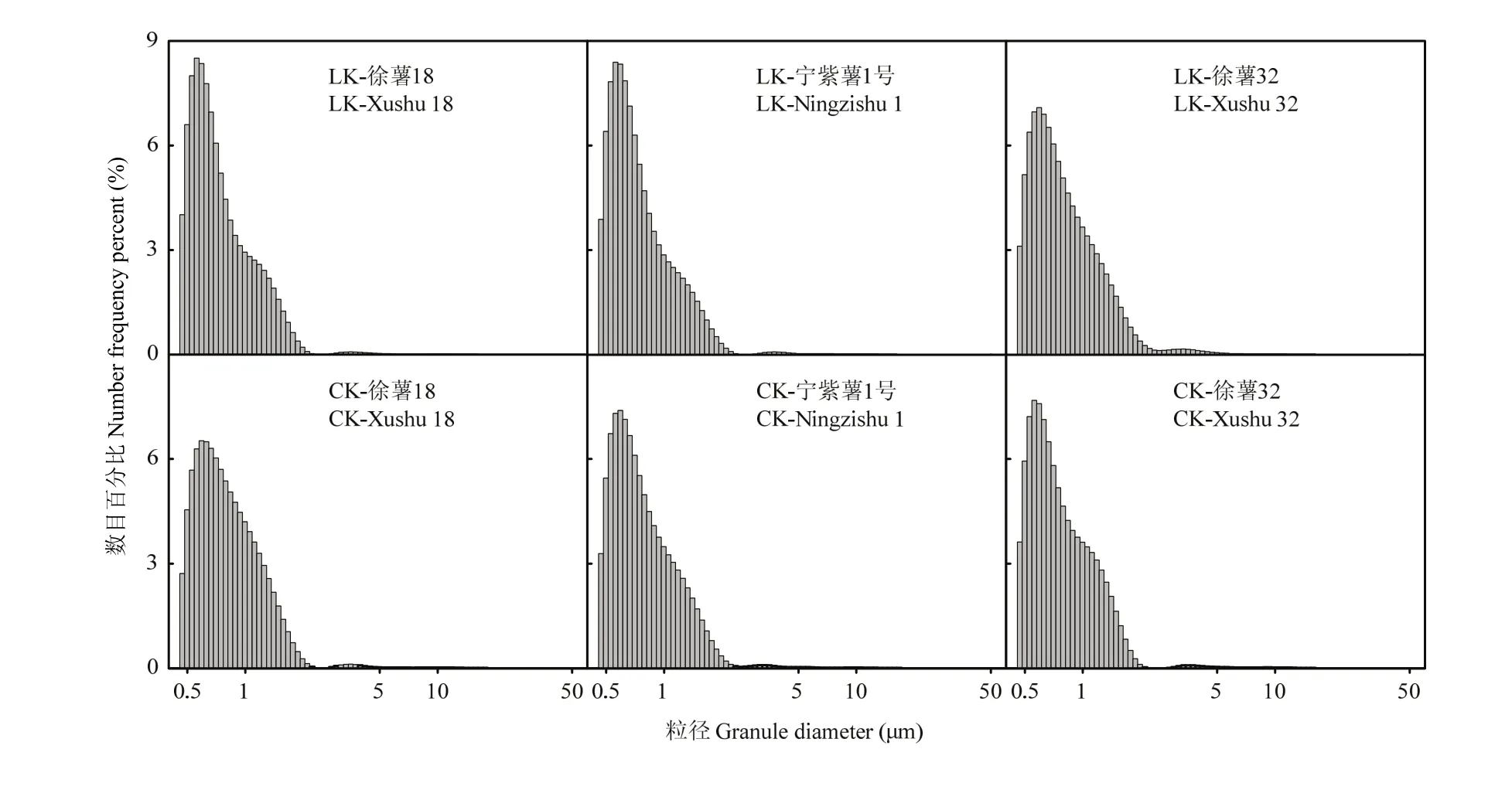
图3 低钾胁迫下甘薯材料块根淀粉粒径数目分布图Fig. 3 Starch granule distribution of three kinds of sweetpotato storage root starch under low K stress (by number)
对比CK处理,低钾胁迫下3个材料的淀粉不同粒径颗粒数目分布发生变化。徐薯32粒径≤2.50 μm的淀粉粒百分比减少,徐薯18与宁紫薯1号增加,但差异均未达显著;徐薯 18平均粒径降低,宁紫薯 1号与徐薯32略增。低钾胁迫下粒径2.50—5.00 μm的淀粉粒百分比变化不一,徐薯32增加,差异达显著水平(P<0.05),徐薯18与宁紫薯1号略降;徐薯32平均粒径减少,徐薯18与宁紫薯1号略增。粒径5.00—30.00 μm 的淀粉粒百分比材料间差异均不显著,徐薯32百分比最高,其平均粒径最低;低钾胁迫下,3个材料的百分比均下降,差异不显著;徐薯18与徐薯32平均粒径略升,宁紫薯1号下降,差异均未达显著。
2.3 低钾胁迫对甘薯块根淀粉热焓特性的影响及其差异
将低钾胁迫与CK处理下的3个不同类型材料块根淀粉DSC曲线相比较(图4),可以发现,不同材料间的差异相当明显,在淀粉吸热过程中,低钾胁迫下宁紫薯1号淀粉糊化起始时间早于其他两个材料,且其吸热的峰值差异较大;曲线所表现出的基本规律是一致的,在 50—120℃范围内均呈单峰趋势变化,单峰均出现在2—6 min时间段内。钾对甘薯淀粉DSC曲线的特征值有较显著的影响,其影响效应与品种有关,这与前人的研究结果相一致[2]。由表5可知,低钾胁迫下,不同甘薯材料间在其淀粉糊化过程中的起始温度(T0)、峰值温度(Tp)和热焓值(△H)普遍低于CK处理的相应指标,应与淀粉组分变化有一定关联;宁紫薯1号在3个特征值上的差异均达显著水平(P<0.05),其△H也高于徐薯18与徐薯32,表明钾对宁紫薯 1号淀粉热特性影响高于其他两材料;徐薯18的Tp在两处理下均显示最高,T0受钾影响小,两者差异不显著;徐薯32的△H最低,T0最高,显示该材料热稳定性好;不同甘薯材料间Tc也有差异,徐薯18显著高于徐薯32与宁紫薯1号,但3个材料的Tc受钾影响均不显著。

图4 低钾胁迫下甘薯材料块根淀粉DSC曲线图Fig. 4 Differential scanning calorimetry (DSC) pattern from three kinds of sweetpotato storage root starch under low K stress

表5 低钾胁迫对甘薯材料块根淀粉DSC特征参数的影响Table 5 DSC characteristic parameter from three kinds of sweetpotato storage root starch under low K stress
2.4 低钾胁迫对甘薯块根淀粉 RVA特性的影响及其差异
由表6可知,宁紫薯1号的最高粘度(PKV)、最低粘度(HPV)和崩解值(BDV)显著高于徐薯18与徐薯32,但回复值(CSV)与糊化温度(PT)低于这两个材料,该结果可能与其直链淀粉含量及淀粉粒径大小相关;徐薯18的PKV最低,PT最高,与淀粉DSC分析结果相一致;两处理间PT差异不显著,但材料间差异显著(P<0.05),表明PT更多受基因型控制,其他外界因素影响较小。对比CK处理,低钾胁迫下,3个材料的PKV、BDV均明显下降,徐薯18、宁紫薯1号两处理间差异达显著水平(P<0.05);徐薯18与宁紫薯1号的HPV、CPV受低钾胁迫影响显著上升,但徐薯32表现相反;CSV是衡量淀粉老化的重要指标,低钾胁迫下徐薯18的CSV有小幅上升,徐薯32与宁紫薯1号均显著下降,宁紫薯1号下降幅度达显著水平(P<0.05)。
2.5 低钾胁迫对甘薯块根淀粉吸湿性的影响及其差异
由图5所示,同一处理下,3个不同甘薯块根淀粉的吸湿性差异未达显著水平,且吸湿性均不高,可能原因是淀粉颗粒外围被包着一层支链淀粉,一般不溶于冷水,只形成悬浮液,只有淀粉悬浮液被加热到一定温度时,淀粉颗粒开始不可逆地剧烈膨胀,颗粒外围的支链淀粉层被胀裂,内部的直链淀粉分子才游离出来,形成不同的淀粉溶解力。对比CK处理,低钾胁迫下3个材料淀粉吸湿性均不同程度降低,徐薯18与宁紫薯1号降低达显著水平(P<0.05),两个处理下徐薯32的吸湿性均最大,显示其淀粉水渗透性强。
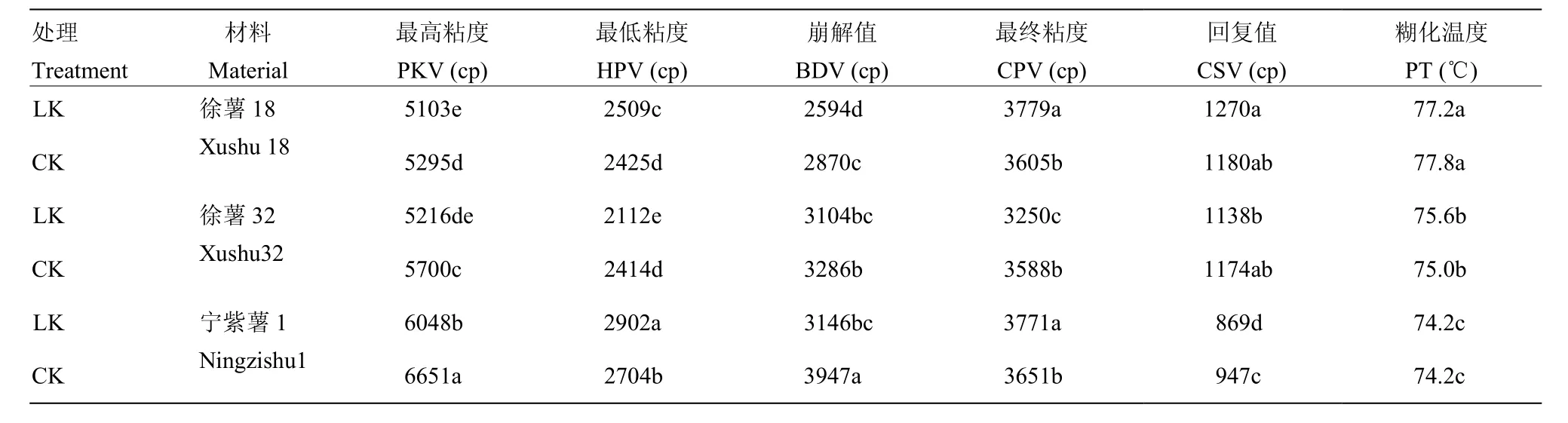
表6 低钾胁迫对甘薯材料块根淀粉RVA特征参数的影响Table 6 RVA characteristic parameter from three kinds of sweetpotato storage root starch under low K stress
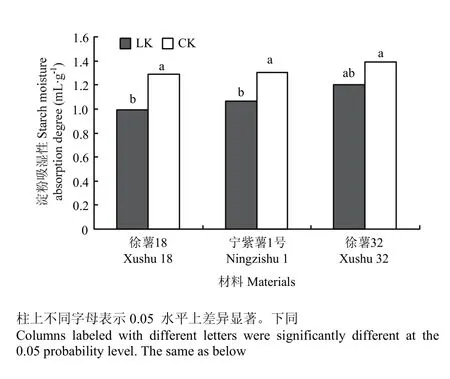
图5 不同处理下3个甘薯材料块根淀粉吸湿性Fig. 5 Moisture absorption degree of starch from three kinds of sweetpotato storage root starch under low K and CK treatments
2.6 低钾胁迫对甘薯块根淀粉膨胀力的影响及其差异
由图6可知,块根淀粉颗粒膨胀力在3个材料间均有一定差异;徐薯 18的淀粉膨胀力最低,与徐薯32、宁紫薯1号间差异显著(P<0.05);对比CK处理,低钾胁迫下3个材料块根淀粉膨胀力不同程度上升,宁紫薯 1号增幅达显著水平(P<0.05),徐薯32淀粉颗粒膨胀力较高,受低钾胁迫影响效应小。甘薯块根淀粉颗粒的膨胀力差异,可能与淀粉颗粒中直链淀粉含量相关,因低直链淀粉颗粒不够坚硬,加热时更易膨胀。
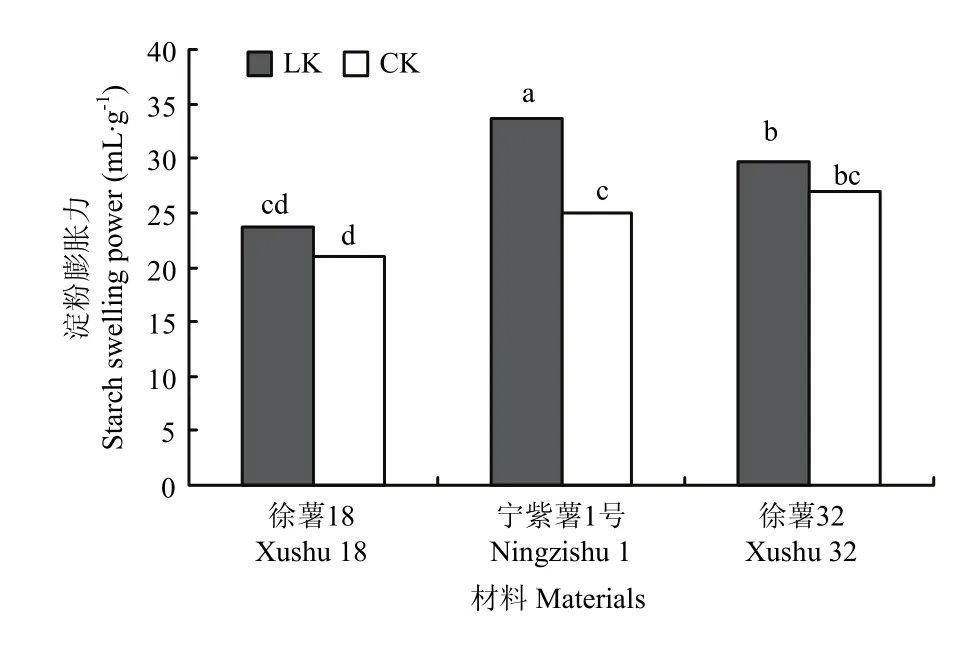
图6 不同处理下3个甘薯材料块根淀粉膨胀力Fig. 6 Swelling power of starch from three kinds of sweetpotato storage root starch under low K and CK treatments
2.7 甘薯块根淀粉主要理化指标相关性分析
相关分析结果显示(表 7),直链淀粉含量直接影响淀粉糊化特性,其含量与 RVA最高粘度值显著负相关,与回复值、糊化温度极显著正相关,与淀粉膨胀力极显著负相关,与DSC峰值温度显著正相关。淀粉体积平均粒径与表面积平均粒径间呈极显著正相关,与淀粉 RVA最高粘度值、最低粘度值显著正相关;体积平均粒径与最终粘度值呈显著正相关。淀粉RVA最高粘度值与崩解值、DSC热焓值显著正相关,与回复值、糊化温度显著负相关;DSC峰值温度与RVA糊化温度显著正相关,与淀粉膨胀力呈极显著负相关。因此,甘薯块根淀粉组分及颗粒粒径大小与淀粉的糊化特性、热特性等理化特性密切相关。
3 讨论
3.1 钾对甘薯块根淀粉主要组分及理化特性差异的影响
环境等外界因素是影响作物淀粉特性的重要因素[22]。研究已表明,施钾减少小麦面粉中直链淀粉含量,显著增加支链淀粉和总淀粉含量,且随钾肥水平的提高,淀粉RVA粘度峰值显著升高[23];施钾增加甘薯块根中干物质的分配比例,使干物质在地上部的分配率降低,增加块根中的淀粉含量,提高块根产量[5];不同施钾期影响甘薯块根干物质积累量和淀粉特性,如追施钾显著提高支链淀粉含量、降低直链淀粉含量,基施钾块根淀粉中大型淀粉粒体积百分数高,高峰期施钾的中、小型淀粉粒体积百分数高[14];但也有研究认为,钾对甘薯淀粉特性的影响甚微[24]。低钾导致植物形态、生理及生化等多方面不利反应[4,18],减少叶片光合产物输出,抑制植物根部生长[25],影响蛋白质与碳水化合物等合成代谢过程[26],最终限制作物产量与品质特性形成[27]。因此,钾作为重要“品质”元素,对提高作物产量和改进品质均有明显的作用[2,28]。
本研究中,不同钾效率型甘薯块根发育对低钾的响应有一定差异。低钾胁迫对3个甘薯材料块根中直链淀粉含量的影响不一,可能与其相关合成酶活性受限与品种选择有关。低钾胁迫小幅降低淀粉颗粒粒径大小,材料间粒径大小与体积、表面积及数目分布存在基因型差异,粒径上的差异应是造成淀粉理化特性差异的重要因素,这些差异需进一步从酶学机理及淀粉精细结构上分析与验证。甘薯淀粉 RVA特征值与粉丝硬度、拉伸强度、断条率等特性密切相关[10];低钾胁迫下块根淀粉糊化的热焓值与 RVA最高粘度均显著下降,改变淀粉的热稳定性,可能对淀粉加工品质产生不利影响。统计分析显示,钾改变淀粉特性有限,表明遗传因素仍是决定甘薯淀粉品质优劣的关键因素,施肥等栽培手段是品质改良辅助措施。但改进栽培措施,实现作物产量提升与品质改良是现实需求,因此,如何利用肥水管理、外源激素调控等手段来缓解如缺钾等逆境下作物生长与品质形成的不利影响,是研究管控、改良甘薯淀粉理化特性的重要内容。
3.2 不同类型甘薯块根淀粉主要组分及理化特性差异
甘薯淀粉颗粒主要由直链淀粉和支链淀粉组成,直链淀粉是衡量淀粉特征重要指标[29]。相关文献报道[2],甘薯块根中直链淀粉含量差异大。笔者曾根据甘薯不同肉色、生物特性等,从甘薯鉴定圃、国家区试、北方区试、省区试及国家甘薯资源库中筛选 50余份甘薯材料测定其块根直链淀粉含量,甘薯淀粉中直链淀粉含量的平均值为 28.32%,变幅为 14.22%—35.12%,变异系数为17.50%,不同基因型甘薯直链淀粉含量差异显著[20]。直链淀粉也直接影响淀粉加工品质特性[2,18]。一般认为,甘薯粉丝品质随直链淀粉含量的提高而改善,直链淀粉含量高,则粉丝品质好[2]。本研究显示,直链淀粉含量影响甘薯淀粉糊化特性及热特性,但与淀粉颗粒粒径大小之间的相关系数较小。
甘薯淀粉的颗粒形状多呈多边形或圆形,部分呈椭圆和铃铛状,粒径分布范围在3.4—27.5 μm之间,平均粒径在8.4—15.6 μm之间[30]。甘薯淀粉大粒率是评价粉丝膨润度的重要指标,随着大粒率的增加,粉丝膨润度呈下降趋势;甘薯淀粉小粒率是评价块根食味、粉丝煮沸损失和膨润度的重要指标,小粒率的降低,煮沸损失减少,粉丝品质改善[2,10]。本研究中,甘薯块根淀粉颗粒按照体积与表面积分布,出现3个峰,且峰界明显,故以此为淀粉粒径大小划分依据,划为小于2.5、2.50—5.0与5.0—30 μm等3个等级,与史春余等[31]划分不一致,可能与测定方法及测定结果不同所致,需进一步验证分析。不同材料间平均粒径均存在一定差异,从体积及表面积分布分析,平均粒径范围分别为12.8—16.25 μm与5.86—7.53 μm,3个材料平均粒径大小顺序为徐薯 32<徐薯 18<宁紫薯1号,从数目分布分析,粒径范围为0.88—0.92 μm,其顺序为徐薯18<宁紫薯1号<徐薯32。
淀粉DSC糊化是淀粉颗粒因吸水溶胀,分子间和分子内氢键断裂,淀粉水合分子扩散的过程,其热焓值变化是评价淀粉粉丝膨润度的重要指标;淀粉热焓值变化减少,膨润度增加,粉丝品质改善[2]。甘薯淀粉热焓值一般在9.6—13.6 J·g-1,糊化起始温度在68.6—71.0℃[29],与本研究结果基本一致。诸多文献报道甘薯淀粉糊化特性的基因型差异[13,18,29],尤其新发现一个甘薯品种,其糊化温度更低,其 DSC结果是39.0-46.9-64.8℃(T0-TP-TC)[32];土温等环境因素也显著影响糊化温度与热焓值(温度越高,糊化温度与热焓值越大)[12,33];研究认为,糊化温度与热焓值的上升主要与减少的 DP6-7短链的支链淀粉量[12],以及降低的颗粒晶体表面能量及增加的淀粉晶体片厚度有关[33]。本研究中,不同处理间淀粉热焓值等指标上的差异与其淀粉颗粒的大小、结构及化学组成均有较为密切的关系。淀粉 RVA谱及其特征值是反映甘薯淀粉糊化特性另一重要指标。在水稻上,崩解值、消减值和回复值是评价其食味品质的主要特征值,一般认为峰值粘度大,崩解值高,而消减值绝对值和回复值低的米饭软而粘,适口性好;崩解值低,而消减值绝对值和回复值高的米饭硬而粘性小,适口性差[34]。目前甘薯上尚未有相关研究结果,可能与甘薯块根中复杂的组分相关。甘薯淀粉 RVA谱及其特征值基因型间存在明显差异[13,35],本研究中,宁紫薯1号直链淀粉含量在3个材料中最低,但发现其最高粘度值、最低粘度与最终粘度均较高,可能与其淀粉颗粒粒径大、磷和脂肪等含量较高有关。
4 结论
低钾胁迫对甘薯块根淀粉组分及其品质理化特性产生影响,改变了淀粉粒径分布,降低淀粉平均粒径、吸湿性和糊化过程中热稳定性。同时,不同类型甘薯淀粉理化特性对低钾响应不同,宁紫薯1号淀粉的起始温度、峰值温度和热焓值变异幅度大于徐薯32,徐薯18与宁紫薯1号的最低粘度值、最终粘度值受低钾胁迫影响显著性上升,而徐薯32的表现则相反。因此,钾是影响甘薯淀粉理化品质的重要元素,低钾胁迫下甘薯块根淀粉理化特性变化与甘薯钾利用效率密切相关。
[1] 唐忠厚, 张允刚, 魏猛, 陈晓光, 史新敏, 张爱君, 李洪民, 丁艳锋.耐低钾与钾高效型甘薯品种(系)的筛选及评价指标. 作物学报, 2014, 40(3): 542-549.
TANG Z H, ZHANG Y G, WEI M, CHEN X G, SHI X M, ZHANG A J, LI H M, DING Y F. Screening and evaluation indicators for low potassium-tolerant and potassium efficient sweetpotato (Ipomoea batatas L.) varieties (lines). Acta Agronomica Sinica, 2014, 40(3): 542-549. (in Chinese)
[2] 陆国权. 甘薯品质性状的基因型与环境效应研究. 气象出版社, 2003.
LU G Q. Genotypic and Environmental Effects on Sweetpotato Quality Traits. Beijing: Meteorological Press, 2003. (in Chinese)
[3] 唐忠厚, 魏猛, 陈晓光, 史新敏, 张爱君, 李洪民, 丁艳锋. 不同肉色甘薯块根主要营养品质特征与综合评价. 中国农业科学, 2014, 47(9): 1705-1714.
TANG Z H, WEI M, CHEN X G, SHI X M, ZHANG A J, LI H M, DING Y F. Characters and comprehensive evaluation of nutrient quality of sweetpotato storage root with different flesh colors. Scientia Agricultura Sinica, 2014, 47(9): 1705-1714. (in Chinese)
[4] TANG Z H, ZHANG A J, WEI M, CHEN X G, LIU Z H, LI H M, DING Y F. Physiological response to potassium deficiency in three sweet potato [Ipomoea batatas(L.)Lam) genotypes differing in potassium utilization efficiency. Acta Physiologiae Plantarum, 2015, 37(8): 1-10.
[5] 唐忠厚, 李洪民, 张爱君, 史新敏, 朱红, 孙健. 长期定位施肥对甘薯块根产量及其主要品质的影响. 浙江农业学报, 2010, 22(1): 57-61.
TANG Z H, LI H M, ZHANG A J, SHI X M, ZHU H, SUN J. Effects of long-term located fertilization on yield and quality of sweetpotato. Acta Agriculturae Zhejiangensis, 2010, 22(1): 57-61. (in Chinese)
[6] 谢一芝, 邱瑞镰, 戴起伟, 吴红中. 甘薯品质的环境效应. 中国甘薯, 1996, 8: 106-109.
XIE Y Z, QIU R L, DAI Q W, WU H Z. Environmental effects on sweet potato quality traits. Chinese Sweetpotato, 1996, 8: 106-109. (in Chinese)
[7] CONSTANTIN R J, JONES L G, HERNANDEZ T P. Effect of potassium and phosphorous fertilization on quality of sweetpotato. Journal of American Society Horticultural Science, 1977, 102: 779-781.
[8] MELVIN S G, LU G Q, ZHOU W J. Genotypic variation for potassium uptake and utilization efficiency in sweet potato (Ipomoeabatatas L.). Field Crop Research, 2002, 77(1): 7-15.
[9] HOOVER R. Composition, molecular structure, and physicochemical properties of tuber and root starches: A review. Carbohydrate Polymer, 2001, 45(3): 253-267.
[10] 谭洪卓, 谭斌, 刘明. 甘薯淀粉性质与其粉丝品质的关系. 农业工程学报, 2009, 25(4): 286-292.
TAN H Z, TAN B, LIU M. Relationship between properties of sweet potato starch and qualities of sweet potato starch noodles. Transactions of The Chinese Society of Agricultural Engineering, 2009, 25(4): 286-292. (in Chinese)
[11] ZHU F, WANG S. Physicochemical properties, molecular structure, and uses of sweetpotato starch. Trends in Food Science and Technology, 2014, 36(2): 68-78.
[12] NODA T, KOBAYASHI T, SUDA I. Effect of soil temperature on starch properties of sweetpotatoes. Carbohydrate Polymers, 2001, 44(3): 239-246.
[13] 唐忠厚, 李洪民, 张爱君, 史新敏, 孙健, 徐飞. 施钾对甘薯常规品质性状及其淀粉 RVA特性影响. 浙江农业学报, 2011, 23(1): 46-51.
TANG Z H, LI H M, ZHANG A J, SHI X M, SUN J, XU F. Effect of potassium fertilizer application on main quality traits and starch RVA characters of sweetpotato. Acta Agriculturae Zhejiangensis, 2011, 23(1): 46-51. (in Chinese)
[14] 柳洪鹃, 姚海兰, 史春余, 张立明. 施钾时期对甘薯济徐23块根淀粉积累与品质的影响及酶学生理机制. 中国农业科学, 2014, 47(1): 43-52.
LIU H J, YAO H L, SHI C Y, ZHANG L M. Effect of potassium application time on starch accumulation and related enzyme activities of sweet potato variety Jixu 23. Scientia Agricultura Sinica, 2014, 47(1): 43-52. (in Chinese)
[15] COLLADO L S, MABESA R C, CORKE H. Genetic variation in the physical properties of sweet potato starch. Journal of Agriculture and Food Chemistry, 1999, 47(10): 4195-4201.
[16] ISHIGURO K, NODA T, YAMAKAWA O. Effect of cultivation conditions on retrogradation of sweet-potato starch. Starch-Stärke, 2003, 55(12): 564-568.
[17] ZHOU W, YANG J, HONG Y, LIU G, ZHENG J, GU Z, ZHANG P. Impact of amylose content on starch physicochemical properties in transgenic sweet potato. Carbohydrate Polymers, 2015, 122: 417-427.
[18] HAFSI C, DEBEZ A, ABDELLY C. Potassium defciency in plants: Effects and signaling cascades. Acta Physiologiae Plantarum, 2014, 36: 1055-1070.
[19] 宁运旺, 马洪波, 许仙菊, 汪吉东, 张辉, 许建平, 陈杰, 张永春.氮磷钾缺乏对甘薯前期生长和养分吸收的影响. 中国农业科学, 2013, 46(3): 486-495.
NING Y W, MA H B, XU X J, WANG J D, ZHANG H, XU J P, CHEN J, ZHANG Y C. Effects of deficiency of N, P, or K on growth traits and nutrient uptakes of sweetpotato at early growing stage. Scientia Agricultura Sinica, 2013, 46(3): 486-495. (in Chinese)
[20] 唐忠厚, 朱晓倩, 李强, 李洪民, 徐飞. 不同基因型甘薯直链淀粉含量差异研究. 食品工业科技, 2011, 32(11): 108-110.
TANG Z H, ZHU X Q, LI Q, LI H M, XU F. Genotype variation in amylase content of sweetpotato. Science and Technology of Food Industry, 2011, 32(11): 108-110. (in Chinese)
[21] RASPER V. Investigations on starches from major starch crops grown in Ghana: Particle size and size distribution. Journal of the Science of Food and Agriculture, 1971, 22(11): 572-580.
[22] 黄华宏, 陆国权, 郑遗凡. 不同生育期甘薯块根淀粉糊化特性的差异. 中国农业科学, 2005, 38(3): 462-467.
HUANG H H, LU G Q, ZHENG Y F. Variation in root starch gelatinization characteristics during the growth and development of sweetpotato. Scientia Agricultura Sinica, 2005, 38(3): 462-467. (in Chinese)
[23] 戴双, 李豪圣, 刘爱峰, 宋健民, 刘建军, 赵振东. 氮钾配施对济南17淀粉理化特性的影响. 麦类作物学报, 2006, 26(4): 107-110.
DAI S, LI H S, LIU A F, SONG J M, LIU J J,ZHAO Z D. Effect of nitrogen and potassium fertilizers on the starch physiochemical properties of Jinan 17. Journal of Triticeae Crops, 2006, 26(4): 107-110. (in Chinese)
[24] NODA T, TAKAHATA Y, SATA T. Physicochemical properties of starches from purple and orange fleshed sweetpotato roots at two levels of fertilizer. Starch-Stärke, 1996, 48(11/12): 395-399.
[25] GERARDEAUX E, JORDAN-MEILLE L, CONSTANTIN J, PELLERIN S, DINGKUHN M. Changes in plant orphology and dry matter partitioning caused by potassium defciency in Gossypium hirsutum(L.). Environmental and Experimental Botany, 2010, 67(3): 451-459.
[26] CAKMAK I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Plant Nutrition and Soil Science, 2005, 168(4): 521-530.
[27] OOSTERHUIS D, LOKA D, KAWAKAMI E, PETTIGREW W. The physiology of potassium in crop production//Advances in Agronomy. Elsevier, 2014: 203-233.
[28] 席承藩. 中国土壤. 北京: 中国农业出版社, 1998: 843-941
XI C F. China Soil. Beijing: China Agriculture Press, 1998: 843-941. (in Chinese)
[29] ZHU F, YANG X, CAI Y, BERTOFT E, CORKE H. Physicochemical properties of sweetpotato starch. Starch- Stärke, 2011, 63(5): 249-259.
[30] NODA T, KIMURA T, OTANI M, SHIMADA T, SAITO A, SUDA I. Physicochemical properties of amylase-starch from transgenic sweet potato. Carbohydrate Polymers, 2002, 49(3): 253-260.
[31] 史春余, 姚海兰, 张立明, 柳洪鹃, 张超, 刘桂玲. 不同类型甘薯品种块根淀粉粒粒度的分布特征. 中国农业科学, 2011, 44(21): 4537-4543.
SHI C Y, YAO H L, ZHANG L M, LIU H J, ZHANG C, LIU G L. Starch granule size distribution in storage loots of different types of sweetpotato cultivars. Scientia Agricultura Sinica, 2011, 44(21): 4537-4543. (in Chinese)
[32] KATAYAMA K, KOMAE K, KOHYAMA K, KATO T, TAMIYA S, KOMAKIE K. New sweet potato line having low gelatinization temperature and altered starch structure. Starch-Stärke, 2002, 54(2): 51-57.
[33] GENKINA N K, NODA T, KOLTISHEVA G I, WASSERMAN L A, TESTER R F, YURYEV V P. Effects of growth temperature on some structural properties of crystalline lamellae in starches extracted from sweet potatoes (Sunnyred and Ayamurasaki). Starch-Stärke, 2003, 55: 350-357.
[34] 宁慧峰. 氮素对稻米品质的影响及其理化基础研究[D]. 南京: 南京农业大学, 2011.
NING H F. The effect of nitrogen on rice quality and its physicochemical foundation[D]. Nanjing: Nanjing Agricultural University, 2011. (in Chinese)
[35] COLLADO L S, MABESA R C, CORKE H. Genetic variation in the physical properties of sweet potato starch. Journal of Agriculture and Food Chemistry,1999, 47(10): 4195-4201.
(责任编辑 杨鑫浩)
Starch Physico-Chemical Properties and Their Difference in Three Sweetpotato (Ipomoea batatas(L.) Lam) Genotypes Under Low Potassium Stress
TANG ZhongHou1,2, ZHANG AiJun1, CHEN XiaoGuang1, JIN Rong1, LIU Ming1, LI HongMin1, DING YanFeng2
(1Xuzhou Sweetpotato Research Center/Key Laboratory of Sweetpotato Biology and Genetic Breeding, Ministry of Agriculture, Xuzhou 221131, Jiangsu ;2College of Agronomy, Nanjing Agricultural University, Nanjing 210095)
【Objective】Sweetpotato (Ipomoea batatas (L.) Lam) is a typical K-preferred starchy root crop. Potassium (K)deficiency (namely low potassium), an important abiotic stress in plants, has become a major limiting element for obtaining crop high yield and good quality. However, in China, K deficiency in soils exists commonly in sweetpotato-growing areas. Sweetpotato starch utilization is closely related with its physico-chemical properties. The influence of low potassium on starch physic-chemical properties was investigated in an experiment conducted by using three sweetpotato cultivars with different potassium utilization efficiencies (KUE) in response to low potassium(K) field condition, aiming at providing reference for improving sweetpotato root properties and potassium utilization efficiency. 【Method】Three sweetpotato cultivars with different potassium utilization efficiencies (KUE) were chosen and cultivated under different potassium field condition, including two treatments (low potassium treatment, LK, and normal potassium treatment, CK).【Result】The results showed that under low K condition, the main components (amylose, amylopectin, nitrogen, total phosphorus and total lipid) in sweetpotato storage root starch changed to a certain extent. There were some differences among the three cultivars in the main components. Three distribution peaks in ≤2.50 μm, 2.50-5.00 μm and 5.00-30.00 μm were observed in starch granule size obviously. The slight decrease was observed in the whole average granule size under low K, which of Xushu32 had the least decrease in volume distribution. However, there was a significant difference in decrease of average granule size of Ningzishu1. The percentage, average granule size and peak value of different granule sizes in three distributions had some differences among three cultivars. Starch T0(gelatinization temperature), Tp(peak temperature) and ΔH (heat enthalpy) were basicially lower than that in CK treatment, and Ningzishu1 had significant differences in the three characteristic values. Peak viscosity and breakdown viscosity in three genotypes had an obvious decrease. Hot paste viscosity, cool paste viscosity in Xushu18 and Ningzishu1 significantly increased, but Xushu32 was on the contrary. Moisture absorption degree of starches in three genotypes was reduced differently. There was a significant difference in the decline of moisture absorption in Xushu18 and Ningzishu1. In addition, swelling power of starch increased differently in three storage root starchs. There was a significant difference in a rise of swelling power of starch in Ningzishu1. Xushu32 had high starch swelling power, but low K had a little effect on it. Correlation analysis showed that the main physico-chemical indicators of sweetpotato starch are closely related. The low K changes starch compositions and particle sizes in sweetpotato root, which led to different degrees of influence on starch gelatinization properties and thermal properties.【Conclusion】The results of the study findings indicated that potassium is a key plant element influencing sweetpotato starch physico-chemical properties. Some obvious effects on physico-chemical properties and main components exist in storage root starch under low K stress. The responses of sweetpotato genotypes differing in potassium utilization efficiency to low K stress are different.
sweetpotato (Ipomoea batatas(L.) Lam); low potassium stress; storage root; starch; physico-chemical properties
2016-07-22;接受日期:2016-10-13
国家自然科学基金(31461143017)、国家公益性行业(农业)科研专项(201403039)、国家甘薯产业体系(CARS-11-07B)、江苏省自然科学基金(BK20151162)、徐州市科技项目(KC15N0017)
联系方式:唐忠厚,E-mail:zhonghoutang@sina.com
猜你喜欢
幼儿100(2021年31期)2021-10-30 03:43:02
家教世界·V家长(2020年10期)2020-11-20 02:02:13
幼儿画刊(2020年6期)2020-07-27 14:00:50
中国粮油学报(2019年4期)2019-07-12 09:06:32
中成药(2018年2期)2018-05-09 07:19:50
现代园艺(2017年19期)2018-01-19 02:50:01
中国塑料(2016年2期)2016-06-15 20:29:57
农业工程学报(2016年6期)2016-05-17 09:36:39
江苏调味副食品(2015年3期)2015-02-28 01:56:39
云南中医学院学报(2014年4期)2014-07-31 18:22:23
