
暗纹东方鲀生长性状相关微卫星标记筛选
2017-01-18 06:47马爱军孙建华王广宁崔文晓王新安刘大勇郭正龙
海洋科学 2016年10期
马爱军, 邹 杰, 孙建华, 王 婷, 王广宁, 崔文晓, 王新安, 刘大勇, 郭正龙
暗纹东方鲀生长性状相关微卫星标记筛选
马爱军1, 2, 邹 杰1, 3, 孙建华1, 3, 王 婷1, 王广宁1, 崔文晓1, 3, 王新安1, 2, 刘大勇4, 郭正龙4
(1. 中国水产科学研究院黄海水产研究所农业部海洋渔业可持续发展重点实验室青岛市海水鱼类种子工程与生物技术重点实验室, 山东青岛266071; 2. 青岛海洋科学与技术国家实验室海洋生物学与生物技术功能实验室, 山东青岛 266071; 3. 上海海洋大学水产与生命学院, 上海201306; 4. 江苏中洋集团股份有限公司, 江苏南通226600)
为了筛选与暗纹东方鲀()生长相关的分子标记, 作者采用SSR结合BSA技术对同龄孵化群体的同池养殖暗纹东方鲀生长差异性状标记进行筛选。利用85对微卫星引物对暗纹东方鲀生长快、慢各30尾个体构建的两个基因池进行分析, 共筛选到14个差异位点。然后分析这14个差异微卫星位点在这60个暗纹东方鲀个体中的基因型差异, 结果表明: 位点TOP03、TOG01、fms15、fms75与生长性状呈现极显著负相关关系(<0.01), fms89与生长性状呈现极显著正相关关系(<0.01)。用另外30个个体(生长快慢各15个)进行验证实验后, 结果只有位点fms15、fms75与生长性状显著相关, 相关系数分别为–0.411和–0.384。这两个微卫星位点对于暗纹东方鲀生长性状有显著效应, 为开展暗纹东方鲀的分子标记辅助育种提供了有价值的参考标记。
暗纹东方鲀(); 生长性状; 微卫星; 相关性分析
暗纹东方鲀()属硬骨鱼纲(Osteichthyes)、鲀形目(Telraodontiformes)、鲀科(Tetradontidae)、东方鲀属(), 其分布集中于北太平洋西部地区的东海及黄海海域, 且可进入长江到江阴一带, 并可定居太湖[1]。暗纹东方鲀不仅能够养血补气, 而且向来以肉味鲜美闻名, 为传统美食中的典范, 是具有很高经济价值的特种水产养殖品种[2-3]。在实际养殖中, 暗纹东方鲀养殖技术日益成熟, 养殖规模不断扩大, 养殖产量逐年增高, 到2014年预期产量已达到40 000t以上[2], 但都普遍存在同一暗纹东方鲀群体出现生长性状差异明显的情况, 往往一个养殖周期需要淘汰掉许多小的个体, 加之暗纹东方鲀具有相互残食的特性[3], 小个体易受伤且死亡率极高, 而且还会受伤得病感染其他鱼体[4]。因此, 进行暗纹东方鲀大小差异的微卫星标记研究既为暗纹东方鲀的良种选育提供依据, 也对提高养殖产量和经济效益具有深远的意义。
微卫星(Microsatellite)目前不仅广泛应用于遗传多样性分析重要性状标记的筛选[5]、遗传图谱的构建[6]和发育研究[7]等, 而且可用于遗传性疾病的连锁分析和基因诊断、品种鉴定、农作物及动物育种等领域[8]。微卫星在水产动物生长性状分子标记方面的运用上也取到了一定的成果, Cnaani等[9]在利用微卫星标记对橙色莫桑比克罗非鱼()和奥利亚罗非鱼()的杂交F2代进行与生长性状的相关性分析, 筛选得到位点UNH130和体长相关。佟广香等[10]研究哲罗鱼 ()生长速度相关性状的微卫星筛选得到标记106INRA、166TUF与测量的7个生长性状显著相关; 111TUF和210TUF与6个生长性状显著相关。刘伟等[11]研究3种鲤鱼()群体的生长性状与微卫星标记的关系时发现: 黄河鲤()的生长性状和位点Mfw5相关; 建鲤()的生长性状与位点Cca09、Hlj013、Mfw2、MfW7和 Mfw29相关; 黑龙江野鲤()的生长性状与位点Mfw4、Mfw6和Mfwl1相关。王桂兴等[12]研究利用30对牙鲆()微卫星标记对雌核发育个体进行分析, 得到有8个座位分别与体质量、体长、体高性状的相关性显著。目前有关利用微卫星标记对暗纹东方鲀生长差异性状分析研究较少, 为提高暗纹东方鲀的育种效率本研究拟运用85对微卫星标记, 采用SSR结合BSA技术比较分析暗纹东方鲀群体的生长差异性状, 筛选出可能与暗纹东方鲀生长性状相连锁的微卫星标记, 为进行暗纹东方鲀的苗种选育提供理论依据。
1 材料与方法
1.1 实验材料
本研究所用暗纹东方鲀样本由江苏中洋河豚庄园公司提供, 取自2010年的同池同时孵化的混合群体, 约180日龄的个体。分别选取生长性状处于两极端的差异个体各30尾, 其中, 生长快的F(fast)组个体全长都大于13 cm(14.8 cm±1.8 cm); 生长慢的S(slow)组个体全长都小于6 cm(4.9 cm±1.1 cm)。每个样本形态学测量完后, 剪取部分尾鳍鳍条分装于离心管中并置于–80℃冰箱中保存以备后用。
1.2 实验引物
使用引物序列参照Ma等[13]和自己设计暗纹东方鲀引物共16对, 古川聪史[14]、郝君[15]和Kai等[16]红鳍东方鲀()微卫星引物69对, 由上海生工生物科技有限公司合成, 实验所用部分微卫星引物信息见表1。
1.3 实验方法
1.3.1 基因组DNA的提取及检测
取保存两组样本F组和S组的尾鳍各30 mg, 按照基因组DNA提取试剂盒提取DNA。并用1%的琼脂糖凝胶电泳进行鉴定, 核酸蛋白测定仪检测DNA浓度, 无菌水稀释到浓度为50 ng/μL, –20℃冷冻保存。
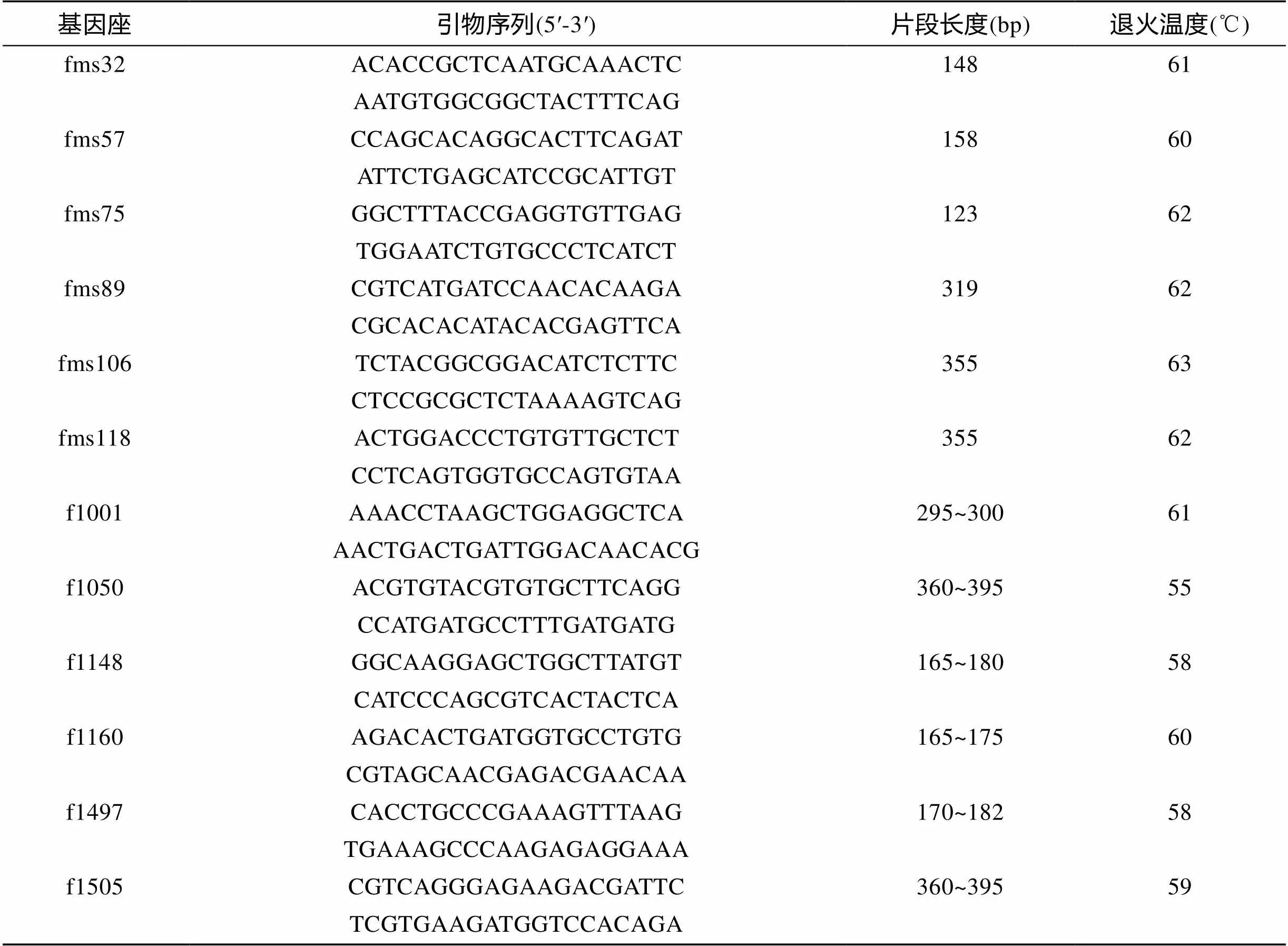
表1 本实验所用部分微卫星引物的核心序列、引物序列、退火温度和片段长度
续表
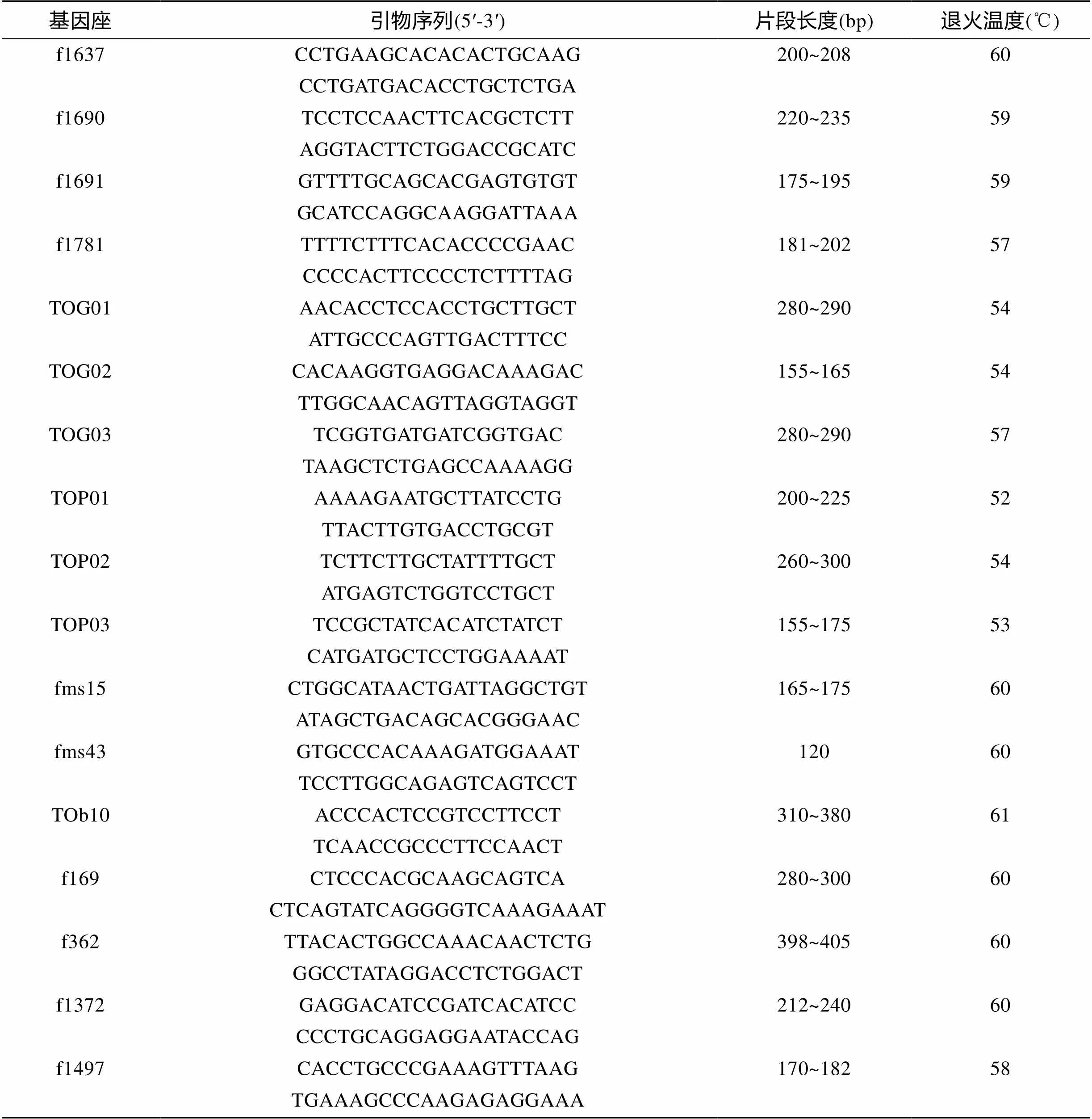
基因座引物序列(5′-3′)片段长度(bp)退火温度(℃) f1637CCTGAAGCACACACTGCAAG200~20860 CCTGATGACACCTGCTCTGA f1690TCCTCCAACTTCACGCTCTT220~23559 AGGTACTTCTGGACCGCATC f1691GTTTTGCAGCACGAGTGTGT175~19559 GCATCCAGGCAAGGATTAAA f1781TTTTCTTTCACACCCCGAAC181~20257 CCCCACTTCCCCTCTTTTAG TOG01AACACCTCCACCTGCTTGCT280~29054 ATTGCCCAGTTGACTTTCC TOG02CACAAGGTGAGGACAAAGAC155~16554 TTGGCAACAGTTAGGTAGGT TOG03TCGGTGATGATCGGTGAC280~29057 TAAGCTCTGAGCCAAAAGG TOP01AAAAGAATGCTTATCCTG200~22552 TTACTTGTGACCTGCGT TOP02TCTTCTTGCTATTTTGCT260~30054 ATGAGTCTGGTCCTGCT TOP03TCCGCTATCACATCTATCT155~17553 CATGATGCTCCTGGAAAAT fms15CTGGCATAACTGATTAGGCTGT165~17560 ATAGCTGACAGCACGGGAAC fms43GTGCCCACAAAGATGGAAAT12060 TCCTTGGCAGAGTCAGTCCT TOb10ACCCACTCCGTCCTTCCT310~38061 TCAACCGCCCTTCCAACT f169CTCCCACGCAAGCAGTCA280~30060 CTCAGTATCAGGGGTCAAAGAAAT f362TTACACTGGCCAAACAACTCTG398~40560 GGCCTATAGGACCTCTGGACT f1372GAGGACATCCGATCACATCC212~24060 CCCTGCAGGAGGAATACCAG f1497CACCTGCCCGAAAGTTTAAG170~18258 TGAAAGCCCAAGAGAGGAAA
1.3.2 BSA基因池的建立及其PCR扩增
F和S两组暗纹东方鲀各30个样本, 从F组每个样本抽取10 μL DNA溶液混合成F组基因池, 同样方法获得S组基因池。用85对微卫星引物对F和S两组基因池进行PCR扩增, 体系为: buffer 1.4 μL、dNTP 1.2 μL (mmoL/μL)、上下游引物各0.6 μL(10 μmol)、模板1 μL (50 ng/μL)、taq酶 0.2 μL(5 U/μL)、补充去离子水10 μL。PCR程序是: 预变性(95℃5 min), 循环30次(变性94℃30 s、退火30 s、复性72℃30 s), 延伸(72℃10 min), 结束(4℃)。将85对微卫星引物的PCR扩增产物变性, 在8%的变性聚丙烯酰胺凝胶中电泳分离, 使用硝酸银法染色[17], 记录带型并用扫描保存电泳结果。
1.3.3 筛选差异条带及个体PCR扩增
比较分析两个基因池聚丙烯酰胺电泳条带, 初步找出F和S两组具有差异性条带的微卫星位点作为候选的生长差异的微卫星分子标记, 将筛选出的具有差异的引物按照上述的PCR条件, 对构建基因池的60个暗纹东方鲀个体进行PCR扩增、聚丙烯酰胺凝胶电泳及硝酸银法染色, 再次记录差异条带。
1.3.4 生长性状差异条带分析
使用SPSS软件对筛选出的差异等位基因片段与暗纹东方鲀生长性状这两个变量的进行相关性检验, 判断差异等位基因片段和暗纹东方鲀生长性状的相关性情况, 筛选出与暗纹东方鲀生长差异性状显著相关的位点。
1.3.5 验证生长性状差异位点
从江苏中洋河豚庄园公司再次采样, 来自不同家系混养的同日龄的暗纹东方鲀, 生长显著快、慢各15尾, 将筛选的微卫星位点, 对上述新样本个体进行PCR扩增、聚丙烯酰胺凝胶电泳及硝酸银法染色, 统计特异性条带并进行相关性检测, 以验证筛选到标记的准确性。
1.3.6 差异等位基因片段切胶测序
将验证实验的差异条带进行切胶回收、纯化, 送至上海生工生物科技有限公司进行克隆、测序, 以验证差异片段的是否为相应序列。
2 结果
2.1 BSA基因池的差异条带筛选
使用试剂盒提取基因组DNA, 紫外线分光光度计进行测定及琼脂糖检测后, 选取质量好的DNA用于下一步实验。用提取的高质量DNA构建了F和S两组基因池。85对引物对构建的F和S两组基因池进行PCR扩增, 将扩增产物变性后进行8%的聚丙烯酰胺凝胶电泳及银染, 结果显示有78对引物具有清晰目的条带, 筛选出TOP01、TOP03、TOG01、TOG02、Tob10、Tob13、fms15、fms75、fms89、f169、f362、f383、f1372和f1497共14对引物的电泳条带在F组和S组间具有差异等位基因, 如图1为F和S基因池的差异等位基因的聚丙烯酰胺电泳图。
2.2 统计个体中带型的差异等位基因片段
将F和S两组基因池筛选的14对具有差异等位基因片段的微卫星引物, 对构建基因池的60个个体进行PCR扩增, 通过聚丙烯酰胺凝胶电泳分析结果。结果显示微卫星位点f383扩增产物的PAGE条带模糊无法辨认, 位点Tob13的产物则无多态性、无差异, 所以两个微卫星位点被舍弃。其余位点差异带型统计结果见表2。图2为引物fms15、fms75在两组个体中的PCR扩增带谱, 可以看出两微卫星位点在F组和S组的目的等位基因片段有着较明显的差异, fms15位点180 bp和fms75位点130 bp的差异等位基因片段在F组的扩增带谱中出现的频率较低, 在S组的扩增带谱出现的频率较高(箭头指示的为差异等位基因片段)。
F. 生长快组; S. 生长慢组
F. fast; S. slow
2.3 差异等位基因片段的SPSS相关性分析
根据表2结果, 通过SPSS13.0软件对筛选的12个微卫星位点的差异等位基因片段与暗纹东方鲀的生长差异性状进行相关性分析, 已知在0.00~0.33呈现弱相关性, 0.33~0.67呈现中相关性, 0.67~1.00呈现强相关性。由表3可得, 位点TOP01、TOG02、Tob10、f169、f362、f1372的生长差异相关性不显著(>0.05); 位点TOP03、TOG01、fms15、fms75、fms89生长差异相关性极显著(<0.01), 且为中度相关(0.33<<0.67), 其中有1个微卫星位点fms89 310 bp的等位基因片段与暗纹东方鲀的F组性状存在一定的正相关性, 其余4个微卫星位点则为负相关性。
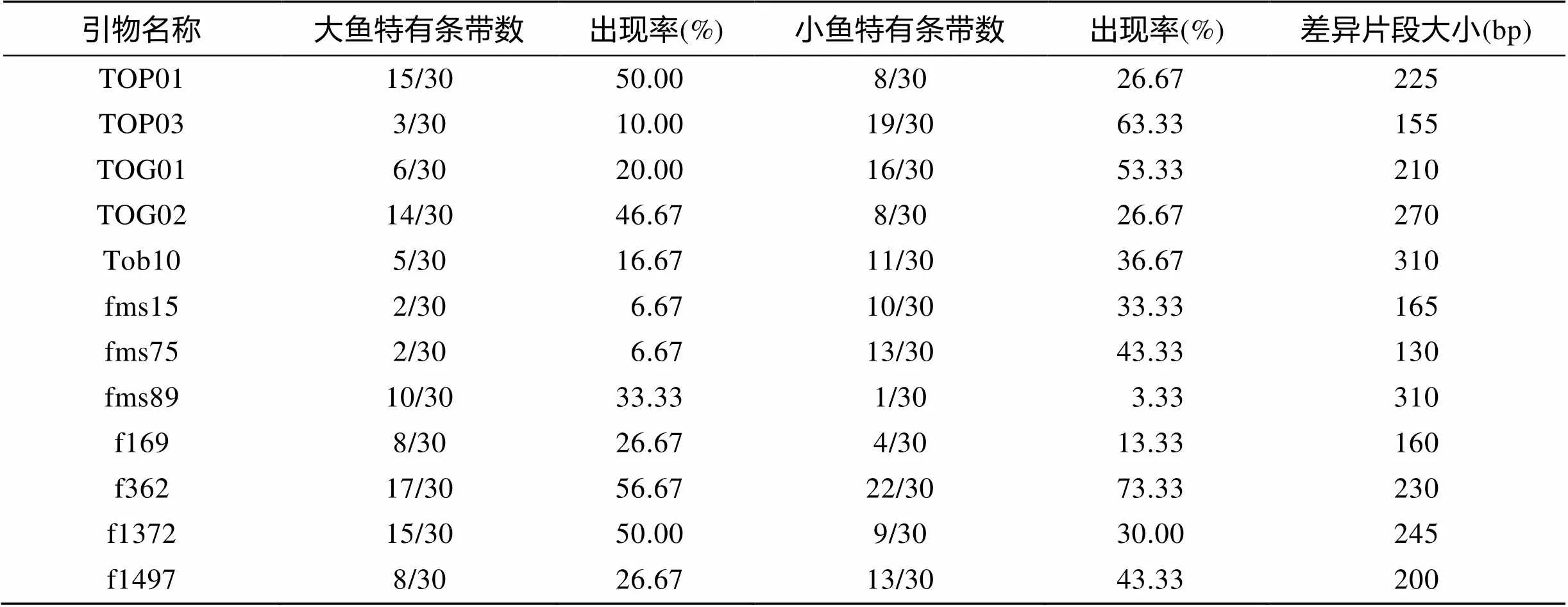
表2 F和S两组12个位点的差异等位基因片段统计表
M. DL500bp分子标记; F. 生长快组; S. 生长慢组
M. DL500bp DNA marker ; F. fast; S. slow
2.4 验证生长显著差异的微卫星位点
经相关性分析得到TOP03、TOG01、fms15、fms75和fms89 5个微卫星位点与生长差异性状存在极显著相关关系, 且为中等相关; 位点f1497与生长性状存在显著相关, 但相关强度较弱。然后, 将重新选取的不同群体的暗纹东方鲀生长差异显著的快和慢各15个个体进行5个微卫星引物的PCR扩增、聚丙烯酰胺凝胶电泳及硝酸银染色, 统计差异片段, 并进行相关性分析(表4)。由表4可得, 位点fms15扩增的差异等位基因片段在生长差异的F和S组重新出现的频率分别为0和33.33%; 位点fms75扩增的差异等位基因片段在生长差异的F和S组重新出现的频率分别为6.67%和40.00%, 且分析相关性关系均呈现显著性, 而其余3个则相关性不显著。得到的微卫星位点fms15和fms75的结果基本与实验预期一致, 相关系数分别为–0.411和–0.384, 进一步验证了作者所筛选的与暗纹东方鲀生长性状存在显著相关性的微卫星位点的准确性。图3为fms15、fms75两个位点的PCR扩增带谱(箭头指示的为差异等位基因片段)。
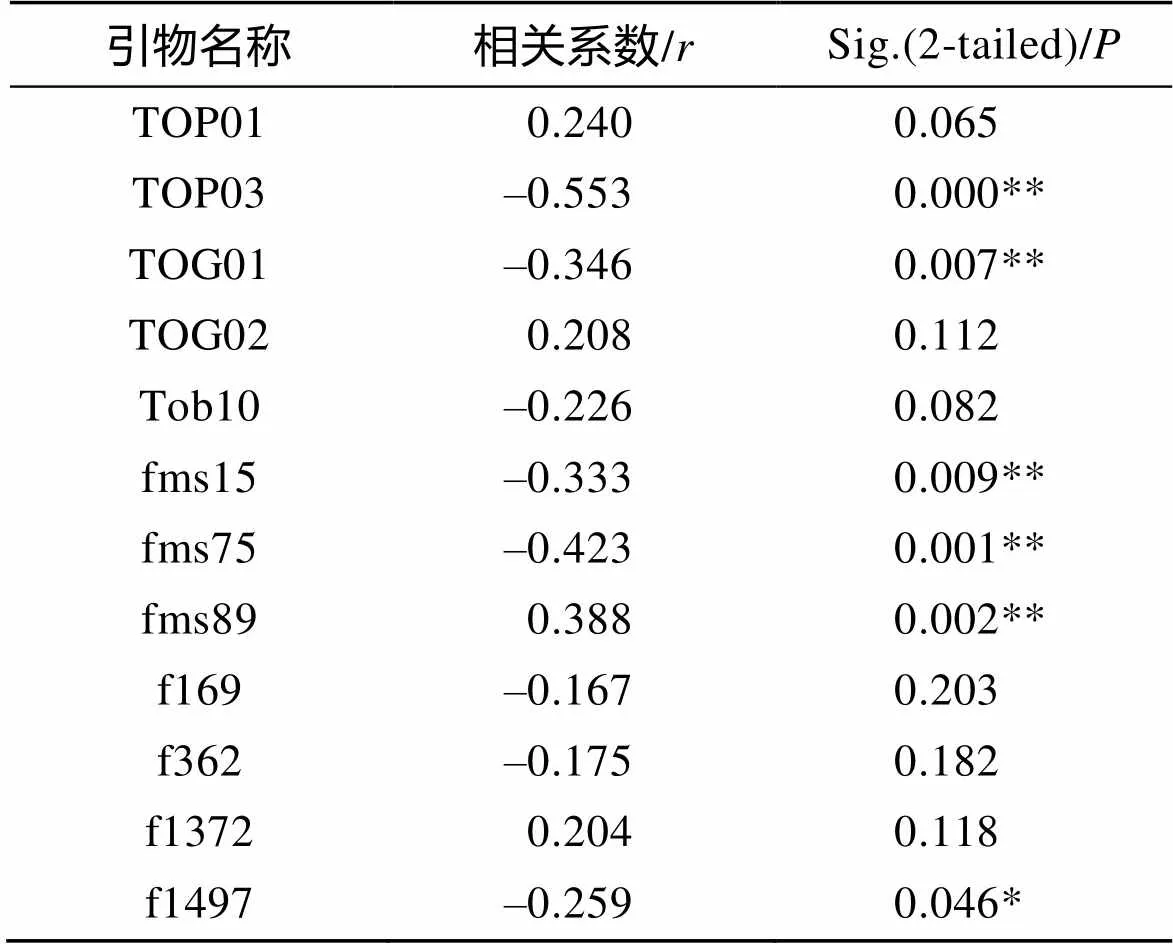
表3 微卫星位点与暗纹东方鲀生长差异性状的相关性分析
注: *. 显著相关(<0.05); **. 极显著相关(<0.01)
2.5 差异等位基因片段切胶测序及BLAST比对
对具有差异条带的微卫星引物进行验证的实验, 得到fms15和fms75存在显著性的差异。然后将其差异条带进行切胶回收, 送至上海生工生物科技有限公司进行序列克隆、测定, 获得微卫星位点fms15差异条带的DNA序列。位点fms15的测序结果为: TCTGGCATAACTGATTAGGCTGTAGCATGAATGTAGCATGTAGCAAGAATGCCAGCATCCTCTTACGGTGTGGAACACACACACACACACACACACACACACACACACACACACACACAGAGAGGGCTATCCATTAGCGAAAGCAATCTGCAGTTCCCGTGCTG TCAGCTATA, 通过NCBI数据库BLAST比对, 证实该片段序列和红鳍东方鲀发布的基因组一段高度吻合, 同源性达到98%, 差别在于序列中有3个碱基发生置换(G-C、C-G、G-A); 同时发现该序列预测功能可能与RNA结合蛋白和CXADR膜蛋白有关。而位点fms75因为基因片段较小、非特异性扩增条带较多、回收DNA浓度较低, 导致未能获得理想扩增结果。
M. DL500bp分子标记; F. 生长快组; S. 生长慢组
M. DL500bp DNA marker; F. fast; S. slow
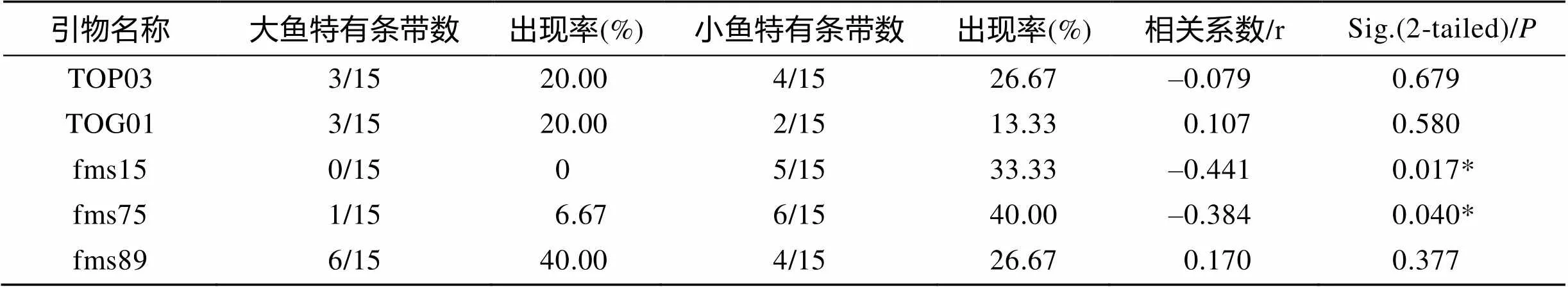
表4 差异等位基因片段在个体扩增带型中的出现次数统计
3 讨论
3.1 微卫星分析方法
分群分离分析法(BSA)的基本原理是从某一分离群体中筛选出一定数量具有目标基因表型差异的个体, 分别构成2个亚群或集团; 该方法可用来快速鉴别与特定基因或染色体区域连锁的标记[18]。现在已经广泛应用于动植物抗性基因、生长相关性状的筛选鉴定工作当中[19-20]。王美玉等[21]采用69个微卫星位点对半滑舌鳎()极大群体和极小群体的基因型分布进行筛选, 得到12个微卫星位点存在差异, 在其后的验证群体与生长性状相关的检验中也呈现出显著差异, 说明采用极端群体进行初筛有效。张天时等[22]采用分离群体标记法也筛选出与中国对虾() 生长性状相关的微卫星位点。本实验则也采用BSA法将收集的暗纹东方鲀分为F和S组, 进行差异微卫星位点的初筛选。然后将这些标记对群体内个体进行相关分析, 并验证筛选极端群体标记的有效性。
微卫星遗传标记为共显性标记, 且检测方便, 数量性状基因位点定位常采用微卫星遗传标记分析, 是目前水产动物数量性状基因位点定位的常用方法之一。近几年, 国内研究者利用微卫星标记也得到了鲫()[23]、青虾()[24]、刺参()[25]和中华绒螯蟹()[26]等生长性状相关联的微卫星位点。因鱼类大多数的经济性状属于数量性状, 遗传基础复杂且受多基因调控, 易受环境影响, 表现为连续变异, 从而不能明确表现型与基因型之间的对应关系[27]。但在基因组时代分析大批DNA分子标记与性状连锁关系、分子标记与数量性状的相关关系, 得到与一个或多个标记的遗传相关的数量性状, 通过改变相应基因型频率, 达到改变表型的目的[28]。本研究采用微卫星分子标记的方法对暗纹东方鲀生长差异性状进行研究, 从分子层面揭示数量性状的遗传基础, 后续的工作将对筛选获得的微卫星标记进行其他群体验证和多代验证, 进一步证实其适应范围、准确性和有效性, 以达到能合理地应用该标记指导育种, 提高育种速度, 改良重要生长性状。
3.2 与生长性状显著相关的微卫星标记
Reed等[29]通过对小鼠进行基因敲除试验后, 发现小鼠()的体质量性状是受多个基因控制的性状。刘伟等[11]对3种不同地理群体的鲤研究发现鲤的生长性状也与多个基因相关, 本研究也得到了类似的结果: 利用85对东方鲀微卫星引物对60个生长性状差异显著的个体进行SSR结合BSA技术的分析, 得到了14个微卫星位点能在暗纹东方鲀F和S两组基因池中扩增出差异条带, 并通过单个样本的SSR分析验证得到了其中5个微卫星位点扩增的差异条带显著, SPSS分析得到该5个微卫星位点与生长差异性状相关极显著, 说明这些微卫星位点可能与暗纹东方鲀的生长差异性状有一定的关联, 如果条带所对应基因位点与生长差异性状无关, 则在大小群体之间出现的频率应该基本一致。再用新的生长性状差异明显的群体进行验证实验, 结果只有fms15和fms75两个位点与生长差异形状存在显著相关, 进一步说明这两个位点与暗纹东方鲀的生长性状存在着一定的关联。为进一步确认本试验中微卫星位点是否与暗纹东方鲀的生长性状有关, 我们将fms15位点测序所得全序列与NCBI的BLAST数据库进行比对, 去寻找与生长相关的基因及蛋白, 作为辅助数据验证本试验结果的可靠性。对比完后发现, 该序列可能与RNA结合蛋白和CXADR膜蛋白有关, 但目前并没有具体的研究结果。
本研究所用BSA法只能对目标基因进行分子标记, 而目标基因与分子标记间连锁的紧密程度不能确定, 也无法得知在遗传连锁图上的位置, 更不能完成后续的QTL定位, 因此目前还不能完全确定fms15或fms75两个位点是否存在多位点关联效应, 该遗传标记是否可以作为暗纹东方鲀优势生长性状分子辅助育种标记还需要进一步深入验证和研究。
[1] 成庆泰, 王存信, 田明诚, 等. 中国东方鲀属鱼类分类研究[J]. 动物学报, 1975, 21(4): 353-378.Cheng Qingtai, Wang Cunxin, Tian Mingcheng, et al. Studies on the Chinese tetraodondid fishes of the genus[J]. Acta Zoologica Sinica, 1975, 21(4): 353-378.
[2] 马爱军, 房金岑, 陈蓝荪, 等. 河豚鱼产业调研报告(上)[J]. 海洋与渔业, 2014(7): 87-92. Ma Aijun, Fang Jincen, Chen Lansun. Research report on puffer fish industry[J]. Ocean and Fishery, 2014(7): 87-92.
[3] 窦海鸽, 刘彦. 暗纹东方鲀苗种互相残食原因及控制措施[J]. 北京水产, 2004, 3: 13-14.Dou Haige, Liu Yan. Reason and control measure of cannibalism inyoung fish[J].Journal of Beijing Fisheries, 2004, 3: 13-14.
[4] 郭正龙. 暗纹东方鲀常见疾病及防治方法[J]. 科学养鱼, 2009, 2: 57-58.Guo Zhenglong.Prevention and treatment methods for common disease of bubble fish[J]. Scientific Fish Farming, 2009, 2: 57-58.
[5] Miyao A, Zhong H S, Monna L, et al . Characterization and genetic mapping of simple sequence repeats in the rice genome[J]. DNA Research, 1996, 3(4): 233-238.
[6] Norri A T, Bradley D G, Cunningham E P. Microsatellite genetic variation between and within farmed and wild Atlantic salmon () populations[J]. Aquaculture, 1999, 180(3): 247-264.
[7] Bierne N, Launey S, Naciri Graven Y, et al. Early effect of inbreeding as revealed by microsatellite analyses onlarvae[J]. Genetics, 1998, 148(4): 1893- 1906.
[8] Beheregaray L B, Ciofi C, Geist D, et al . Genes record a prehistoric volcano eruption in the Galapagos[J]. Science , 2003 , 302 (5642) : 75.
[9] Cnaani A, Hallerman E M, Ron M, et al. Detection of a chromosomal region with two quantitative trait loci, affecting cold tolerance and fish size, in an F2 tilapia hybrid[J]. Aquaculture, 2003, 223(1-4): 117-128.
[10] 佟广香, 匡友谊, 张超, 等. 哲罗鱼生长速度相关性状的微卫星标记筛选[J]. 华北农学报, 2012, 27(z1): 33-39.Tong Guangxiang, Kuang Youyi, Zhang Chao, et al. Identification of microsatellite markers associated with growth rate trails in[J]. Acta Agriculturae Boreali-Sinica, 2012, 27(z1): 33-39.
[11] 刘伟, 苏胜彦, 董在杰, 等. 3个鲤群体的微卫星标记与生长性状相关性分析[J]. 南方水产科学, 2012, 8(3): 17-24.Liu Wei, Su Shengyan, Dong Zaijie, et al. Correlation analysis of microsatellite DNA markers with growth trait among 3 breeding populations of common carp[J].South China Fisheries Science, 2012, 8(3): 17-24.
[12] 王桂兴, 刘永新, 孙效文, 等. 牙鲆微卫星分子标记与生长性状的相关性分析[J]. 东北农业大学学报, 2009, 40(7): 77-84.Wang Guixing, Liu Yongxin, Sun Xiaowen, et al. Correlation analysis of microsatellite DNA marks with growth trait of Japanese flounder ()[J]. Journal of Northeast Agricultural University, 2009, 40(7): 77-84.
[13] Ma Hongyu, Chen Songlin, LiaoXiaolin, et al. Isolation and characterization of polymorphic microsatellite loci from a dinucleotide-enriched genomic library of obscure puffer () and cross-species amplification[J]. Conserv Genet, 2009, 10(4): 955-957.
[14] 古川聪史. 红鳍东方鲀与高生长性状有关的遗传学及分子生物学的研究[D]. 日本: 东京大学农学学院生命科学研究科, 2009.Guchuan Congshi. Study on the genetics and molecular biology of growth-related traits in[D].Japan: Tokyo University of Agriculture and Life Sciences, 2009.
[15] 郝君, 孙效文, 孟雪松. 红鳍东方鲀微卫星DNA多态性初步分析[J]. 上海水产大学学报, 2006, 15(1): 21-24.Hao Jun, Sun Xiaowen, Meng Xuesong. Analyzing the polymorphisms ofwith microsatellite[J].Journal of Shanghai Fisheries University, 2006, 15(1): 21-24.
[16] Kai W, Kikuchi K, Tohari S, et al. Integration of the genetic map and genome assembly of fugu facilitates insights into distinct features of genome evolution in teleosts and mammals[J]. Genome Biol Evol, 2011, 3: 424-442.
[17] 梁宏伟, 王长忠, 李忠, 等. 聚丙烯酰胺凝胶快速、高效银染方法的建立[J]. 遗传, 2008, 30(10): 1379- 1382.Liang Hongwei, Wang Changzhong, Li Zhong, et al. Improvement of the silver-stained technique of polyacrylamide gel electrophoresis[J].Hereditas, 2008, 30(10): 1379-1382.
[18] Michelmore R W, Paran I, Kesseli R V. Identification of markers linked to disease resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations[J]. Proc Nati Acad Sci USA, 1991, 88(21): 9828- 9832.
[19] 陈超, 石拓, 孙曙光, 等. 应用RAPD标记对东方鲀属进行种类鉴别及其聚类分析[J]. 海洋水产研究, 2001, 22(3): 32-36.Chen Chao, Shi Tuo, Sun Shuguang, et al.Identification and phylogenetic relationships among four species of puffer fish inas determined by RAPD markers[J]. Marine Fisheries Research, 2001, 22(3): 32-36.
[20] 崔建洲, 申雪艳, 杨官品, 等. 红鳍东方鲀基因组微卫星特征分析[J]. 中国海洋大学学报, 2006, 36(2): 249-254.Cui Jianzhou, Shen Xueyan, Yang Guanpin, et al. The analysis of simple sequence repeats ingenome[J]. Periodical of Ocean University of China, 2006, 36(2): 249-254.
[21] 王美玉, 刘海金. 半滑舌鳎生长性状的微卫星标记筛选[J]. 海洋渔业, 2012, 34(2): 121-129.Wang Meiyu, Liu Haijin. Correlation analysis between growth-related traits and microsatellite markers in half smooth tongue sole ()[J]. Marine Fisheries, 2012, 34(2): 121-129.
[22] 张天时, 刘萍, 李健, 等. 中国对虾与生长性状相关微卫星DNA分子标记的初步研究[J]. 海洋水产研究, 2006, 27(5): 201-209. Zhang Tianshi, Liu Ping, Li Jian, et al. Preliminary study on specific microsatellites markers related to growth trait in[J]. Marine Fisheries Research, 2006, 27(5): 201-209.
[23] 贾志武, 郑先虎, 匡友谊, 等. 鲫微卫星标记与几个生长性状的相关性分析[J]. 水产学杂志, 2012, 25(6): 1-6.Jia Zhiwu, Zheng Xianhu, Kuang Youyi, et al.Correlation analysis on four growth traits in crucian carpby SSR and EST-SSR markers[J]. Chinese Journal of Fisheries, 2012, 25(6): 1-6.
[24] 傅洪拓, 万山青, 付春鹏, 等. 青虾生长性状相关的微卫星标记筛选[J]. 水生生物学报, 2010, 34(5): 1043-1048.Fu Hongtuo, Wan Shanqing, Fu Chunpeng, et al.Screening of microsatellite markers associated with growth trats in[J]. Acta Hydrobiologica Sinica, 2010, 34(5): 1043-1048.
[25] 孙国华, 杨建敏, 孙孝德, 等. 刺参微卫星标记与生长性状体重、体长的相关分析[J]. 水产学报, 2011, 35(4): 501-508.Sun Guohua, Yang Jianmin, Sun Xiaode, et al. Correlation analysis of microsatellite DNA markers with growth traits of body weight and length in[J]. Journal of Fisheries of China, 2011, 35(4): 501-508.
[26] 吴滟, 付春鹏, 蒋速飞, 等. 中华绒螯蟹微卫星标记与生长性状相关性的初步分析[J]. 水生生物学报, 2011, 35(2): 197-202.Wu Yan, Fu Chunpeng, Jiang Sufei, et al. Preliminary studies on the correlation between microsatellite markers and growth traits in chinese mitten crab ()[J]. Acta Hydrobiologica Sinica, 2011, 35(2): 197-202.
[27] 许可, 马爱军, 王新安, 等. 大菱鲆()生长性状相关的微卫星标记筛选[J]. 海洋与湖沼, 2009, 40(3): 577-583.Xu Ke, Ma Aijun, Wang Xinan, et al. Microsatellites molecular markers and the correlation to growth trait of[J].Oceanologia Et Limnologia Sinica, 2009, 40(3): 577-583.
[28] Wang Z, Zhou J, Ye Y, et al. Genetic structure and low genetic diversity suggesting the necessity for conversation of the Chinese longsnout catfish,(Pisces: Bagriidae)[J]. Environ Biol Fish, 2006, 75(4): 455-463.
[29] Reed D R, Lawler M P, Tordoff M G, et al. Reduced body weight is a common effect of gene knockout in mice[J]. BMC Genet , 2008, 9(4): 1-6.
Screening growth-related microsatellite markers in
MA Ai-jun1, 2, ZOU Jie1, 3, SUN Jian-hua1, 3, WANG Ting1, WANG Guang-ning1, CUI Wen-xiao1, 3, WANG Xin-an1, 2, LIU Da-yong4, GUO Zheng-long4
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences; Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture; Qingdao Key Laboratory for Marine Fish Breeding and Biotechnology, Qingdao 266071, China; 2. Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China; 3. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China; 4.Jiangsu Zhongyang Group Limited by Share Ltd, Nantong 226600, China)
A population ofwith significant growth differences was investigated for screening molecular markers linked to growth traits, using SSR combined with bulked segregation analysis (BSA). Two gene pools were respectively constructed using 30 DNA templates isolated fromsamples with significantly different growth rates and then amplified by 85 pairs of microsatellite primers. The unique alleles were found in 14 loci. These 60individuals were genotyped by SSRs. The results showed that loci TOP03, TOG01, fms15, and fms75 had a highly significant negative correlation with growth trait (< 0.01), while fms89 showed extremely significant positive correlation with growth trait (< 0.01). The verification test using another population of 30 randomly selectedwith significantly different growth rates indicated that only loci fms15 and fms75 were significantly correlated with growth trait, with correlation coefficients of –0.384 and –0.411, respectively. Therefore, these two microsatellite loci with a significant linkage with growth traits could be used as reference markers for marker-assisted breeding of.
; growth trait; microsatellite; correlation analysis
(本文编辑: 谭雪静)
Sept. 17, 2015
[Jiangsu Science and Technology Support Program, No. BE2013345; Jiangsu Aquatic Three New Projects , No. Y2013-12]
S917.4
A
1000-3096(2016)10-0016-09
10.11759//hykx 20150917002
2015-09-17;
2015-12-03
江苏省科技支撑计划资助项目(BE2013345); 江苏省水产三新工程资助项目(Y2013-12)
马爱军(1971-), 女, 山东荣成人, 博士, 研究员, 博士生导师, 主要从事鱼类繁育养殖生物学与遗传育种研究, E-mail: maaj@ ysfri.ac.cn
猜你喜欢
四川蚕业(2022年2期)2022-11-19
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
智慧健康(2021年17期)2021-07-30
世界科学技术-中医药现代化(2020年2期)2020-07-25
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
四川动物(2017年6期)2017-12-12
四川动物(2017年4期)2017-07-31
临床医药文献杂志(电子版)(2017年11期)2017-05-17
现代检验医学杂志(2016年5期)2016-08-20
