
10个白杨派无性系抗寒性的比较与评价1)
2017-01-17 02:23藕丹樊军锋周永学高建社
东北林业大学学报 2017年1期
藕丹 樊军锋 周永学 高建社
(西北农林科技大学,杨凌,712100)
10个白杨派无性系抗寒性的比较与评价1)
藕丹 樊军锋 周永学 高建社
(西北农林科技大学,杨凌,712100)
以西北农林科技大学选育的6个I-101(Populusalba)×84K(P.alba×P.glandulosa)优良杂种无性系(‘02-3-32’‘02-8-21’‘02-21-13’、秦白杨1号、秦白杨2号、秦白杨3号)、4个I-101×毛白杨优良杂种无性系(‘04-16-10’‘04-14-15’‘04-17-12’‘04-19-14’),以及I-101、84K、毛白杨30号3个对照无性系的1年生休眠枝条为试验材料,在一系列低温胁迫(-15、-20、-25、-30、-35 ℃)处理24 h后,测定他们的电导率、丙二醛(MDA)质量摩尔浓度、可溶性蛋白质量分数、超氧化物歧化酶(SOD)活性等4个不同的生理生化指标变化,并通过隶属函数法对各无性系的抗寒性的结果进行比较与评价。结果表明:各无性系间抗寒性差异明显,抗寒性由强到弱表现为秦白杨2号、‘04-16-10’‘04-14-15’‘02-21-13’、秦白杨3号、84K、I-101、秦白杨1号、毛白杨30号、‘04-17-12’‘04-19-14’‘02-3-32’‘02-8-21’。研究表明,在4个抗寒指标综合评价中,秦白杨2号抗寒性表现最好;‘04-16-10’与‘04-14-15’抗寒性也明显强于其他无性系,可为抗寒性优良新品种选择及推广提供参考。
白杨派;电导率;半致死温度;抗寒能力
Taking annual dormancy branches of six hybrids of I-101 (Populusalba)×84K (P.alba×P.glandulosa), four hybrids of I-101×P.tomentosaselected and bred by Northwest A&F University and three control clones (84K, I-101,P.tomentosaNO.30) as experimental materials, we studied the changes of conductivity, the molality of MDA, the contents of soluble protein and SOD activities after the plants were treated under a series of low temperature (-15 ℃, -20 ℃, -25 ℃, -30 ℃, and -35 ℃) for 24 h, and evaluated the differences of the clones in cold resistance by using membership function method. There were obvious differences among the clones of the ability of cold resistance. The descending order was Qin Bai yang NO.2, ‘04-16-10’, ‘04-14-15’, ‘02-21-13’, Qin Baiyang NO.3, 84K, I-101, Qin Baiyang NO.1,P.tomentosaNO.30, ‘04-17-12’, ‘04-19-14’, ‘02-3-32’, and ‘02-8-21’. NO.2 Qin Baiyang was the best in all the test results. ‘04-16-10’ and ‘04-14-15’ clone materials had better cold resistance than others, which can provide reference for the popularization of new varieties with excellent cold resistance.
白杨派杨树是西北地区乡土杨树种,因其生长迅速、材质优良、树型优美,所以成为北方地区造林绿化的重要树种之一。但其栽培区域受寒冷、干旱等气候条件的限制[1],因此,选育抗寒性强的白杨派树种具有重要意义。在低温胁迫下,植物体内的细胞酶活性,可溶性蛋白质、糖类质量分数等会随之发生改变,直接或间接影响到植物体内的正常代谢。为适应逆境,植物体内会发生一系列复杂的生理生化反应来抵抗恶劣环境。近年来,在鉴定和评价植物的抗寒性研究中,多利用相对电导率拟合Logistic方程回归[2]来计算植物半致死温度(LT50),同时测定若干个生理指标值,最后利用隶属函数法对各指标进行综合评价来判断植物的抗寒性强弱,具有一定的科学意义[3-5]。
本研究选用西北农林科技大学林学院杨树课题组经十年多时间筛选的10个白杨派优良杂种无性系为供试材料,人工低温胁迫处理后,测定与抗寒有关的生理指标并进行分析,综合评价它们的抗寒性强弱,筛选出抗寒力较强的无性系,为白杨派抗寒性优良新品种的选育与推广提供借鉴。
1 材料与方法
亲本84K、I-101和毛白杨30号为对照种,6个I-101(P.alba)×84K(P.alba×P.glandulosa)的优良杂种无性系(‘02-3-32’‘02-8-21’‘02-21-13’、秦白杨1号、秦白杨2号、秦白杨3号)和4个I-101×毛白杨的优良杂种无性系(‘04-16-10’‘04-14-15’‘04-17-12’‘04-19-14’)为供试树种。13个样本于2016年1月下旬取自西北农林科技大学渭河试验站苗圃。各无性系选择无病虫害、生长状况一致、具有代表性1年生休眠苗5株,取地径约2 cm以上枝条。
用自来水将枝条冲洗干净后剪成约20 cm的枝段,再用去离子水漂洗3~5次,用滤纸将表面的水分吸干,分6组用密封袋分装好,放置于超低温冰箱中进行低温胁迫处理。设定5个温度梯度,即-15、-20、-25、-30、-35 ℃,并以常温为对照。以4 ℃·h-1降温[6],在既定温度下处理24 h。
相对电导率的测定:用去离子水将低温处理好的各无性系枝条冲洗3次,滤纸吸干枝条表面的水分,避开芽眼,切成0.5 cm左右的小段再称取约1 g,分别置于三角瓶中,各加入去离子水25 mL,用封口膜密封,放置于摇床上25 ℃振荡浸提24 h后,用雷磁DDS-307A型电导率仪测定其浸提液初始电导值;之后将样品浸提液在沸水浴中加热35 min,冷却后摇匀测定其电导率作为终电导率值。三角瓶用封口膜密封以确保整个处理过程中水量不变[7]。计算公式为相对电导率=(C1/C2)×100%(其中,C1为煮前电导率;C2为煮后电导率),各无性系重复3次。
生理生化指标的测定:采用巴比妥酸法测定丙二醛(MDA)的质量摩尔浓度,采用考马斯亮蓝染色法测定可溶性蛋白质量分数,采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性[8-9],各无性系重复3次。
数据处理:试验数据采用Excel 2003软件进行处理,并依据隶属函数法综合各项指标进行抗寒性评价;用IBM SPSS Statistics19.0拟合不同温度处理下的相对电导率的Logistic方程,确定低温半致死温度;采用DPS 7.05完成数据的差异显著性测验。其中,
隶属函数:
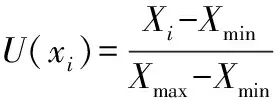
反隶属函数:
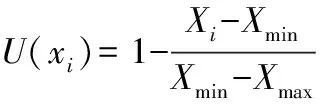
式中:U(xi)为隶属函数值;Xi为无性系某项指标的测定值;Xmin和Xmax分别为所有参试无性系中某一指标的最小值和最大值。如果某一指标与综合评判结果为负相关,则用反隶属函数进行转换。
2 结果与分析
2.1 低温处理下各无性系的半致死温度(LT50)
利用Logistic求得各无性系的半致死温度(表1)。13个白杨派无性系的半致死温度差异显著,LT50在-40~-20 ℃。据此得出的半致死温度可将13个无性系抗寒性大致分为3种不同的类型,第I类抗寒性最强(LT50<-34 ℃)(包括秦白杨2号、‘04-16-10’‘04-14-15’‘02-21-13’);第II类抗寒性较强(-34 ℃≤LT50<-29 ℃)(包括秦白杨3号、84K、秦白杨1号、I-101、毛白杨30号);第III类抗寒性较弱(LT50≥-29 ℃)(包括‘04-17-12’‘02-3-32’‘04-19-14’‘02-8-21’)。各无性系按LT50从低到高的顺序依次为秦白杨2号、‘04-16-10’‘04-14-15’‘02-21-13’、秦白杨3号、84K、秦白杨1号、I-101、毛白杨30号、‘04-17-12’‘02-3-32’‘04-19-14’‘02-8-21’。
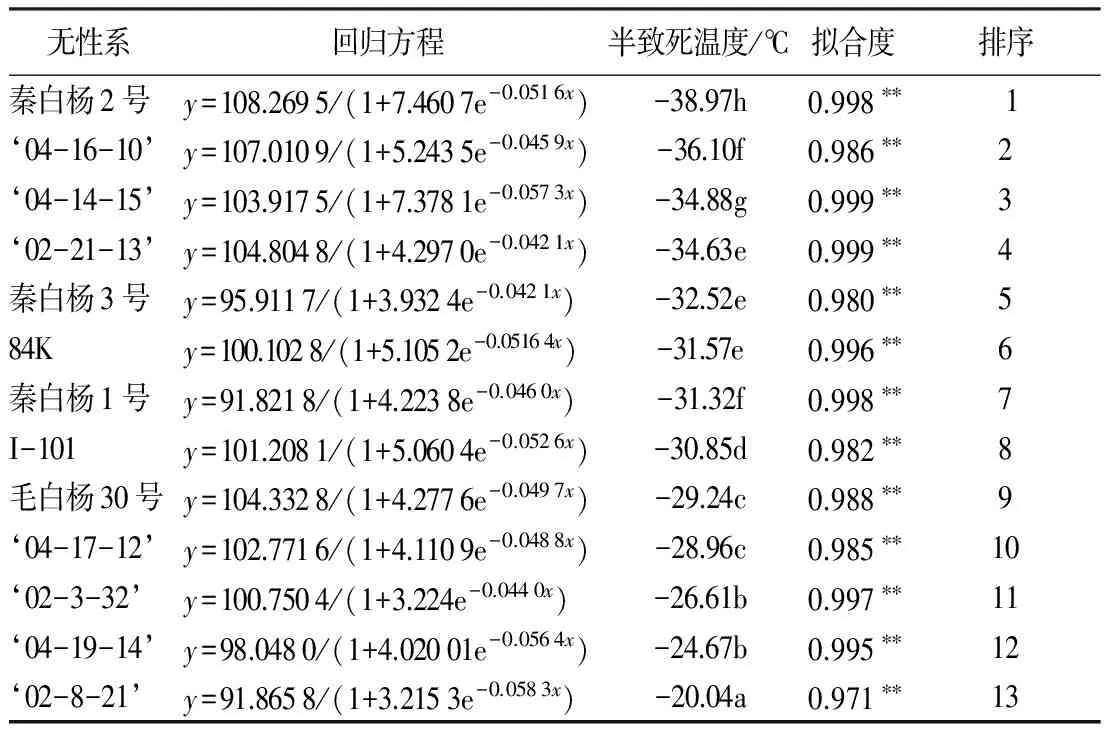
表1 低温处理下各无性系的Logistic方程及其半致死温度
注:** 表示拟合度达到极显著水平;同列数据后不同小写字母表示差异显著(P<0.05)。
2.2 低温处理下各无性系的MDA质量摩尔浓度、可溶性蛋白质量分数和SOD活性变化
丙二醛(MDA)质量摩尔浓度的高低可以反映植物膜系统的受伤程度,与植物抗寒性呈负相关[10-11]。随着温度的降低,13个白杨派树种的MDA质量摩尔浓度出现先升高后下降的变化趋势,但是达到最大值时所对应的温度不同(表2),例如,秦白杨2号,‘04-14-15’和‘04-16-10’在-30 ℃下达到最大值;I-101、84K和秦白杨3号在-25 ℃处理下达到最大值;而‘02-3-32’和‘02-8-21’在-20 ℃时就达到最大值。
可溶性蛋白质量分数越大,植物体的抗寒性也就越高[12]。在整个低温处理过程中,秦白杨2号、‘04-16-10’‘04-14-15’‘02-21-13’可溶性蛋白质量分数均达到23.00 mg·g-1(表2),增幅均达到19.40%;而‘2-8-21’在整个处理过程中,可溶性蛋白质量分数最小,幅度仅为11.43%,说明低温胁迫下,其无法通过迅速增加体内可溶性蛋白质量分数来应对恶劣环境,抗寒性很差。
SOD是植物体内的内源保护酶系统,研究表明,树种抗寒能力越强,SOD活性越高[13]。在低温胁迫下,各枝条中SOD的活性发生明显变化,但是变化规律相似,为先升高后降低(表2)。其中秦白杨2号SOD活性在-30 ℃时达到最大值(400 U·g-1),增幅达到42.02%,说明其抗寒性最强;‘02-8-21’增加幅度最小,为17.29%,整个过程中增加缓慢,说明其相对于其他无性系抗寒性弱。
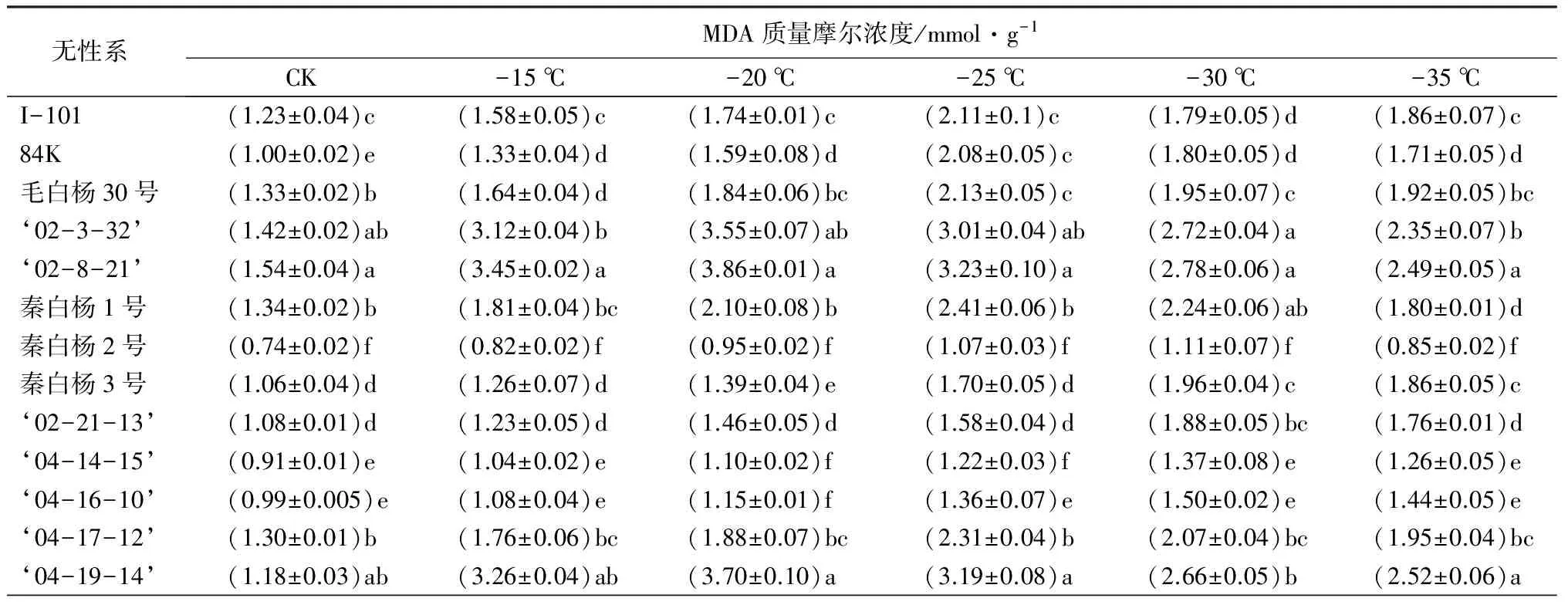
表2 低温处理下各无性系的生理生化指标变化
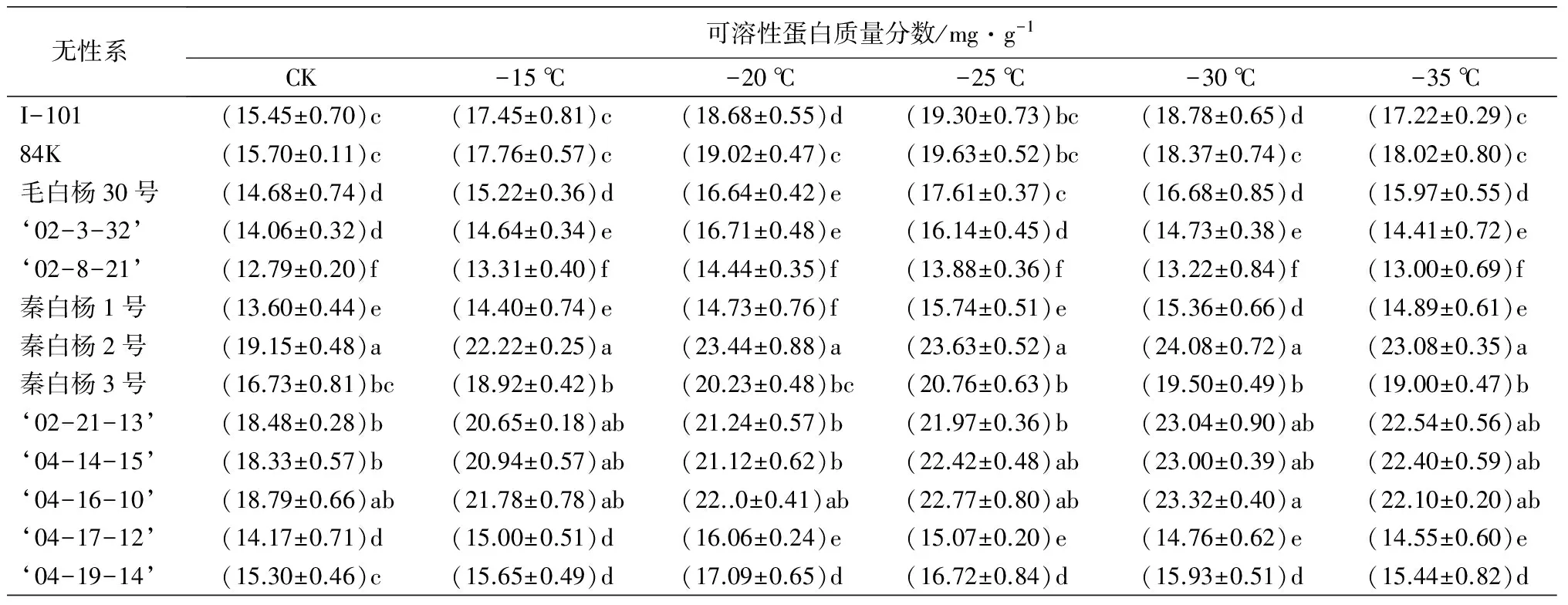
无性系可溶性蛋白质量分数/mg·g-1CK-15℃-20℃-25℃-30℃-35℃I-101(15.45±0.70)c(17.45±0.81)c(18.68±0.55)d(19.30±0.73)bc(18.78±0.65)d(17.22±0.29)c84K(15.70±0.11)c(17.76±0.57)c(19.02±0.47)c(19.63±0.52)bc(18.37±0.74)c(18.02±0.80)c毛白杨30号(14.68±0.74)d(15.22±0.36)d(16.64±0.42)e(17.61±0.37)c(16.68±0.85)d(15.97±0.55)d‘02-3-32’(14.06±0.32)d(14.64±0.34)e(16.71±0.48)e(16.14±0.45)d(14.73±0.38)e(14.41±0.72)e‘02-8-21’(12.79±0.20)f(13.31±0.40)f(14.44±0.35)f(13.88±0.36)f(13.22±0.84)f(13.00±0.69)f秦白杨1号(13.60±0.44)e(14.40±0.74)e(14.73±0.76)f(15.74±0.51)e(15.36±0.66)d(14.89±0.61)e秦白杨2号(19.15±0.48)a(22.22±0.25)a(23.44±0.88)a(23.63±0.52)a(24.08±0.72)a(23.08±0.35)a秦白杨3号(16.73±0.81)bc(18.92±0.42)b(20.23±0.48)bc(20.76±0.63)b(19.50±0.49)b(19.00±0.47)b‘02-21-13’(18.48±0.28)b(20.65±0.18)ab(21.24±0.57)b(21.97±0.36)b(23.04±0.90)ab(22.54±0.56)ab‘04-14-15’(18.33±0.57)b(20.94±0.57)ab(21.12±0.62)b(22.42±0.48)ab(23.00±0.39)ab(22.40±0.59)ab‘04-16-10’(18.79±0.66)ab(21.78±0.78)ab(22..0±0.41)ab(22.77±0.80)ab(23.32±0.40)a(22.10±0.20)ab‘04-17-12’(14.17±0.71)d(15.00±0.51)d(16.06±0.24)e(15.07±0.20)e(14.76±0.62)e(14.55±0.60)e‘04-19-14’(15.30±0.46)c(15.65±0.49)d(17.09±0.65)d(16.72±0.84)d(15.93±0.51)d(15.44±0.82)d
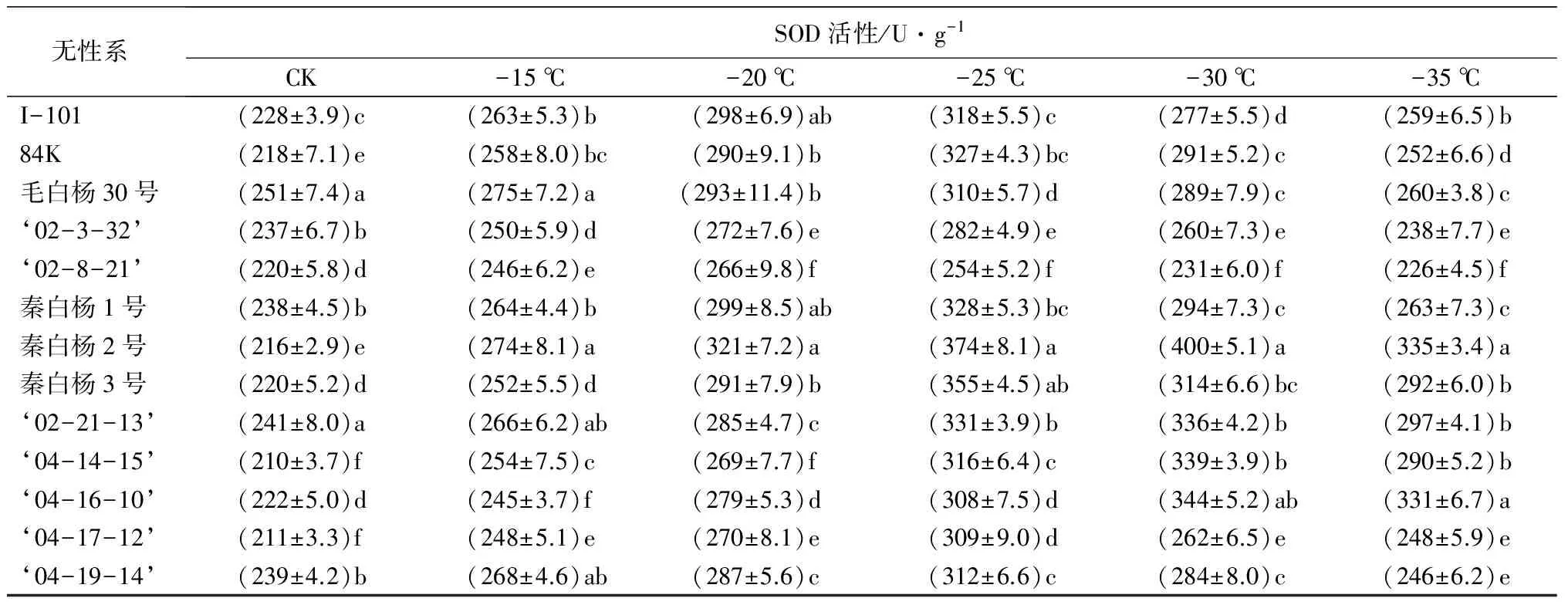
无性系SOD活性/U·g-1CK-15℃-20℃-25℃-30℃-35℃I-101(228±3.9)c(263±5.3)b(298±6.9)ab(318±5.5)c(277±5.5)d(259±6.5)b84K(218±7.1)e(258±8.0)bc(290±9.1)b(327±4.3)bc(291±5.2)c(252±6.6)d毛白杨30号(251±7.4)a(275±7.2)a(293±11.4)b(310±5.7)d(289±7.9)c(260±3.8)c‘02-3-32’(237±6.7)b(250±5.9)d(272±7.6)e(282±4.9)e(260±7.3)e(238±7.7)e‘02-8-21’(220±5.8)d(246±6.2)e(266±9.8)f(254±5.2)f(231±6.0)f(226±4.5)f秦白杨1号(238±4.5)b(264±4.4)b(299±8.5)ab(328±5.3)bc(294±7.3)c(263±7.3)c秦白杨2号(216±2.9)e(274±8.1)a(321±7.2)a(374±8.1)a(400±5.1)a(335±3.4)a秦白杨3号(220±5.2)d(252±5.5)d(291±7.9)b(355±4.5)ab(314±6.6)bc(292±6.0)b‘02-21-13’(241±8.0)a(266±6.2)ab(285±4.7)c(331±3.9)b(336±4.2)b(297±4.1)b‘04-14-15’(210±3.7)f(254±7.5)c(269±7.7)f(316±6.4)c(339±3.9)b(290±5.2)b‘04-16-10’(222±5.0)d(245±3.7)f(279±5.3)d(308±7.5)d(344±5.2)ab(331±6.7)a‘04-17-12’(211±3.3)f(248±5.1)e(270±8.1)e(309±9.0)d(262±6.5)e(248±5.9)e‘04-19-14’(239±4.2)b(268±4.6)ab(287±5.6)c(312±6.6)c(284±8.0)c(246±6.2)e
注:表中数据为平均值±标准差;同列不同字母表示差异显著(P<0.05)。
2.3 各无性系的抗寒性综合评价
利用隶属函数法对13个白杨派无性系的相对导电率、MDA质量摩尔浓度、可溶性蛋白质量分数及SOD活性4项指标综合分析发现,枝条的可溶性蛋白质量分数、SOD活性与各无性系的抗寒性呈正相关,而丙二醛质量摩尔浓度和电解质渗出率与其呈负相关[14-15];同时,隶属函数值(表3)以秦白杨2号最大(0.577 2),其次是‘04-16-10’(0.556 8),‘04-14-15’(0.550 4),‘02-8-21’的隶属函数值最小(0.422 9)。13个白杨无性系的抗寒性由强到弱的顺序为秦白杨2号、‘04-16-10’‘04-16-15’‘02-21-13’、秦白杨3号、84K、I-101、秦白杨1号、毛白杨30号、‘04-17-12’‘04-19-14’‘02-3-32’‘02-8-21’,该结果与通过建立Logistic方程而推算出的抗寒性强弱顺序基本吻合。
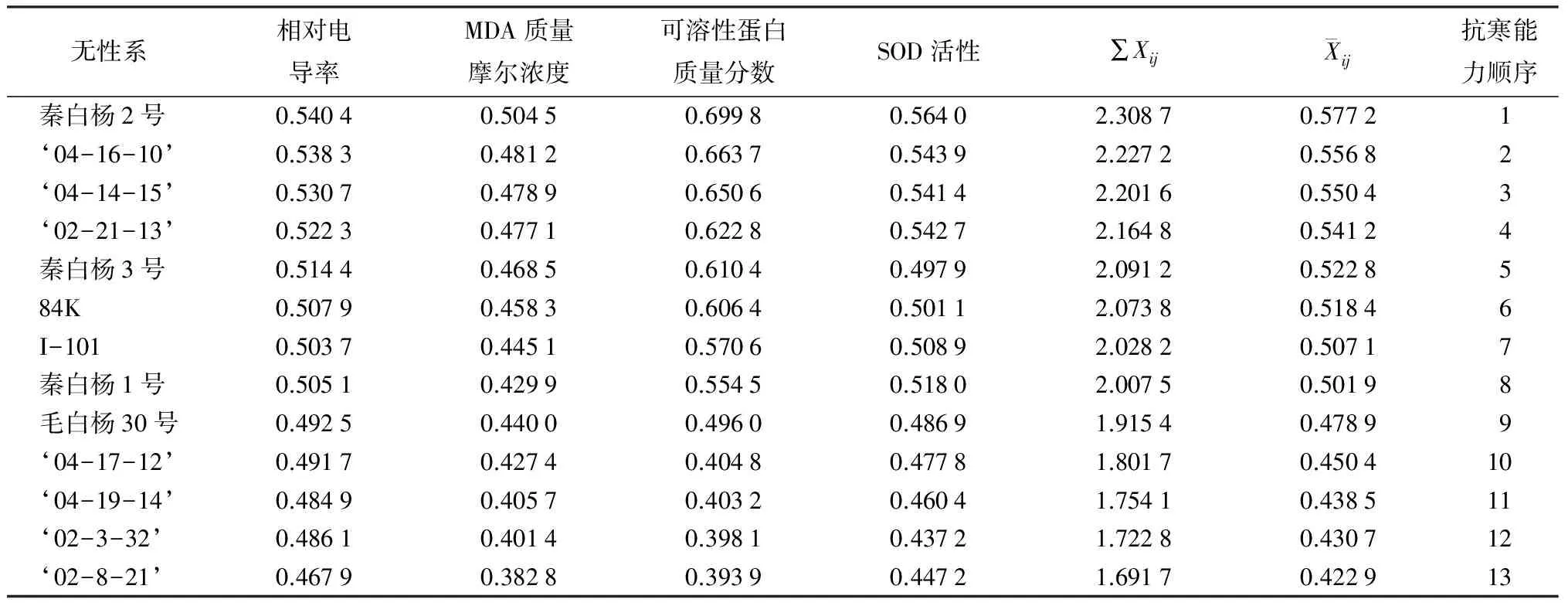
表3 无性系各指标隶属函数值
3 结论与讨论
相对电导率与植物组织在低温胁迫后细胞原生质膜受害程度呈正相关[16],根据相对电导率与温度的关系拟合出的Logistic方程,推算出植物的半致死温度(LT50)。本试验计算出I-101、84K、毛白杨30号、‘02-3-32’‘02-8-21’、秦白杨1号、秦白杨2号、秦白杨3号、‘02-21-13’‘04-14-15’‘04-16-10’‘04-17-12’和‘04-19-14’的LT50分别为-30.85、-31.57、-29.24、-26.61、-20.04、-31.32、-38.97、-32.52、-34.63、-34.88、-36.10、-28.96和-24.67 ℃。由此得到,白杨派无性系的抗寒性由强到弱的顺序为秦白杨2号、‘04-16-10’‘04-14-15’‘02-21-13’、秦白杨3号、84K、秦白杨1号、I-101、毛白杨30号、‘04-17-12’‘04-19-14’‘02-3-32’‘02-8-21’,这与采用隶属函数分析法综合评价结果基本一致,只有秦白杨1号与I-101顺序略有差异。说明通过电导率的测定计算的LT50就能判断植物的抗寒性强弱,这与之前的研究一致[17]。同时,各无性系的半致死温度也为以后的引种提供借鉴。
MDA质量摩尔浓度、可溶性蛋白质量分数以及SOD活性这三个指标在整个低温处理过程中,均在不断变化,且达到最大值的温度有所不同,说明各个无性系对低温调节的敏感度有一定的差异。这三个指标都可以在一定程度上反映出植物的抗寒性大小,但是与综合分析得到的结果差异较大,如仅根据SOD活性隶属函数值得到的抗寒性由大到小的排序是秦白杨2号、‘04-16-10’‘02-21-13’‘04-14-15’、秦白杨1号、I-101、84K、秦白杨3号、毛白杨30号、‘04-17-12’‘04-19-14’‘02-3-32’,所以不能单独依据某一生理生化指标判断植物的抗寒性大小,但这些指标可以粗略判断植物抗寒性强弱,如‘02-21-13’和‘04-14-15’、‘04-19-14’和‘02-3-32’虽然排序中位置都颠倒,但是前者都属于抗寒性强的,后者都属于抗寒性弱的。
自然环境复杂多变,人工低温胁迫不可能完全反映出自然条件的冻害情况,所以本试验在模拟低温环境下测定的结果,可能与各无性系实际抗寒性不完全一致。但本试验所有材料均经过相同的处理且在同一条件下测定,因此,测定结果具有可对比性,可以反映出各无性系之间的抗寒性强弱。并且利用隶属函数法对其抗寒性进行了综合评价,筛选出抗寒性强的优良无性系,即秦白杨2号、‘04-16-10’和‘04-14-15’,它们可作为白杨派抗寒性树种的重要补充,也为后续抗寒性树种的培育与推广提供参考。
[1] 杨敏生,王春荣,裴保华.白杨杂种无性系的抗寒性[J].东北林业大学学报,1997,25(4):20-23.
[2] 令凡,焦健,李朝周,等.不同油橄榄品种对低温胁迫的生理响应及抗寒性综合评价[J].西北植物学报,2015,35(3):508-515.
[3] 姜丽娜,张黛静,宋飞,等.不同品种小麦叶片对拔节期低温的生理响应及抗寒性评价[J].生态学报,2014,34(15):4251-4261.
[4] 曹冉,王海洋.11种常绿阔叶树的抗寒力研究[J].西南大学学报(自然科学版),2014,36(8):57-64.
[5] 金明丽,徐继忠,张钢.苹果砧木枝条电阻抗参数与其抗寒性的关系[J].园艺学报,2011,38(6):1045-1051.
[6] 李晓东,樊军锋,邱兴,等.美洲黑杨×青杨派杂种无性系苗期抗寒性的鉴定与筛选[J].西北林学院学报,2015,30(2):100-104.
[7] 李轶冰,杨顺强,任广鑫,等.低温处理下不同禾本科牧草的生理变化及其抗寒性比较[J].生态学报,2009,29(3):1341-1347.
[8] 邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[9] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2004.
[10] 许瑛,陈发棣.菊花8个品种的低温半致死温度及其抗寒适应性[J].园艺学报,2008,35(4):559-564.
[11] 左利萍,李毅,焦健.渗透胁迫下河北杨叶片的生理响应及相关分析[J].林业科学,2008,44(8):56-61.
[12] 李小琴,彭明俊,段安安,等.低温胁迫对8个核桃无性系抗寒生理指标的影响[J].西北林学院学报,2012,27(6):12-15.
[13] 何丽斯,汪仁,孟祥静,等.茉莉扦插苗对模拟低温的生理响应[J].西北植物学报,2011,30(12):2451-2458.
[14] 李晓宇,杨成超,彭建东,等.杨树苗期抗寒性综合评价体系的构建[J].林业科学,2014,50(7):44-51.
[15] 马文涛,樊卫国.贵州野生柑橘的抗寒性测定和综合评价[J].西北植物学报,2014,34(10):2063-2069.
[16] 林艳,郭伟珍,徐振华,等.大叶女贞抗寒性及冬季叶片丙二醛和可溶性糖含量的变化[J].中国农学通报,2012,28(25):68-72.
[17] 张纪林,谢晓金,教忠意,等.几种冬青属植物抗冻能力比较[J].园艺学报,2005,32(3):477-481.
Comparison and Evaluation on Cold-tolerance of Ten Varieties in Sect.Leuce//
Ou Dan, Fan Junfeng, Zhou Yongxue, Gao Jianshe
(Northwest A & F University, Yangling 712100, P. R. China)//Journal of Northeast Forestry University,2017,45(1):16-19,54.
Sect.Leuce; Conductivity; Semi-lethal temperature (LT50); Cold resistance
藕丹,女,1991年12月生,西北农林科技大学林学院,硕士研究生。E-mail:18829784191@163.com。
樊军锋,西北农林科技大学林学院,教授。E-mail:fanjf28@163.com。
2016年7月4日。
S722.5;S792.12
1)“十三五”国家重点研发计划林业资源培育及高效技术创新专项(SQ2016YFNC030041)。
责任编辑:任 俐。
猜你喜欢
新农业(2021年15期)2021-08-23
新农民(2020年21期)2020-12-08
中国林副特产(2019年1期)2019-02-18
东坡赤壁诗词(2018年1期)2018-03-31
江苏农业科学(2017年10期)2017-07-21
河南林业科技(2017年1期)2017-06-10
新疆农垦科技(2016年2期)2016-08-21
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
园艺与种苗(2015年10期)2015-02-27
