
续断属植物SRAP标记的遗传亲缘关系分析
2016-12-22 09:18:49李晓玲程岁寒
西南农业学报 2016年7期
李晓玲,程岁寒,杨 进
(1.三峡大学生物与制药学院,湖北 宜昌 443002;2.三峡库区生态环境教育部工程研究中心(三峡大学),湖北 宜昌 443002)
续断属植物SRAP标记的遗传亲缘关系分析
李晓玲1,2,程岁寒1,2,杨 进1*
(1.三峡大学生物与制药学院,湖北 宜昌 443002;2.三峡库区生态环境教育部工程研究中心(三峡大学),湖北 宜昌 443002)
本文采用10 对SRAP引物,对10 份续断属植物样本进行遗传亲缘关系分析。结果表明,10 对引物共扩出566条谱带,平均每对引物扩增出 56.6 条带,片段大小为150~2000 bp,其中多态性条带为362条,平均每对引物扩增的多态性条带为 36.2 条,平均多态性条带百分率为 64 %,结果表明续断属植物具有丰富的遗传变异。UPGMA聚类分析表明10 份续断属植物分为2个大类和4个亚类,大头续断和深紫续断有一定的亲缘关系,与其它续断属植物间遗传差异较大;日本续断、恩施续断、大理续断和川续断遗传亲缘关系较近,其中大理续断和川续断聚为一个小类群,支持大理续断并入川续断。
续断属植物;SRAP;遗传亲缘关系
续断属(DipsacusL.)植物种类较多、资源较丰富,是川续断科(Dipsacaceae) 的一个属,常见于长江以南各省区,湖北西部和四川东部分布较多,甘肃和陕西南部也有少量分布,为传统中药材,始载于《神农本草经》,列为上品[1]。中药续断的原植物主要是川续断(DipsacusasperoidesC.Y.Cheng et T.M.Ai),深紫续断也少量进入中药续断中;本属主要以根或果实入药,含有三萜皂甙、黄酮甙、环烯醚萜甙、甾醇及糖类等化学成分;用于治疗肝肾不足,风湿痹痛,腰膝酸软,筋伤骨折,跌扑损伤,崩漏,胎漏[2-3]。我国川续断科共6属40余种,药用的有5属18种4变种,其中续断属共18种(包括2变种),9种2变种可作药用[3-4]。长期以来,在续断进入药典的分类及种间遗传亲缘关系分析中,存在着较多的争议[5-6]。我国续断属17种2变种植物花粉的光学显微镜和电镜扫描的结果表明该属花粉为近球形,种间差异较小,该属植物为一自然分类群[7]。随着分子生物学技术的发展,采用分子生物学技术对植物进行分类和遗传亲缘关系分析成为一种重要的方法。但目前国内续断属植物分子水平的研究比较少。
相关序列扩增多态性(Suquence-related amplified polymorphism,SRAP)是最早用于芸苔属植物中的一种基于PCR反应的分子标记技术[8]。原理是针对基因启动子、内含子中GC少AT多,而外显子中GC含量多的特点来设计引物对开放读码框(ORFs)进行扩增,因内含子、启动子与间隔区长度在不同个体中的差异从而产生多态性。SRAP标记具有简便、高效、产率高、稳定、成本低、高共显性、在基因组中分布均匀的特点,目前主要用于遗传多样性分析[9]、比较基因组学[10]、遗传作图[11]等方面的研究。本研究将采用10 对SRAP引物对10份续断属植物进行分子鉴定和亲缘关系分析,为研究续断属植物从分子水平上进行分类提供科学依据。
1 材料与方法
1.1 供试材料
本试验材料主要采自云南、重庆、湖北和四川。将新鲜叶片自生长地采集后,放入装有硅胶的自封袋中,用冰盒带回实验室置于-70 ℃ 超低温冰箱中保存,其材料名称及来源见表1。反应中Taq酶、MgCl2、10×PCR Buffer、dNTPs等均购自上海捷瑞生物工程有限公司。SRAP优化体系材料采自湖北恩施土家族自治州大山顶牛角坪,引物为Me8-Em3。SRAP引物信息见表2,引物购自北京三博志远生物工程有限公司。

表1 材料名称及来源

表2 用于SRAP分析的引物信息
1.2 试验方法
1.2.1 基因组DNA的提取 取续断属植物幼嫩叶片,采用略作改动的Doyle and Doyle[12]的常规 CTAB法提取基因组DNA,用核酸蛋白测定仪测定DNA的浓度和纯度(260 nm/280 nm),并稀释至20 ng·μl-1储存备用。
1.2.2 引物筛选及SRAP-PCR扩增反应 按照Li和 Quiros[13]提出的原则设计了100对SRAP引物,由北京三博志远生物工程有限公司合成,表2为10对筛选的多态性较高的引物组合。扩增反应在Eppendorf-PTC100 TM型 PCR 仪上进行,20 μl体系中含有dNTPs浓度为 0.25 mmol·L-1、引物浓度 0.3 μmol·L-1、Mg2+浓度 1.0 mmol·L-1、1 UTaq酶、1×PCR buffer、40 ng 模板DNA。反应程序为94 ℃ 预变性 1 min,94 ℃ 变性 1 min,33 ℃ 复性 1 min,72 ℃ 延伸 1 min,5个循环;94 ℃变性 1 min,52 ℃ 复性 1 min,72 ℃ 延伸 1 min,30个循环;72 ℃ 延伸 5 min;最后4 ℃保存。扩增产物在6 %聚丙烯酰胺凝胶上电泳,银染法检测[14]。
1.2.3 SRAP标记数据统计分析 根据聚丙烯酰胺凝胶电泳的结果,在相同迁移率位置上进行 “ 1、0” 标注,有DNA条带的记为1,没有DNA条带的记为0。在NTSYS-PC 2.10软件中,根据SM相似系数法求得各样品间的遗传相似性矩阵,再用非加权算术平均聚类(UPGMA)方法进行聚类,构建聚类图。
2 结果与分析
2.1 SRAP标记的多态性
对100对SRAP引物组合进行多态性筛选,采用得到的多态性较高的10对SRAP引物组合对10份续断样本进行PCR扩增,共扩出566条谱带,平均每对引物扩增出 56.6 条带,片段大小为150~2000 bp,其中多态性条带为362条,平均每对引物扩增的多态性条带为 36.2条,平均多态性条带百分率为 64 %。结果表明续断属植物具有丰富的遗传变异。
2.2 UPGMA 聚类分析
采用NTSYS-PC 2.10软件对0/1矩阵数据进行续断属植物各样品间的遗传距离和相似系数分析,结果见表3。10份续断材料两两之间的遗传相似度相对较高,变化范围为0.6460[DL(W)与SZ]~0.8956[DL(H)与DL(WS)],平均遗传相似度为0.764944;遗传距离在0.1044[DL(H)与DL(WS)]~0.3540[DL(W)与SZ]之间,平均遗传距离为0.235056。其中相似性最大的为2种大理续断DL(H)与DL(WS),相似系数为0.8956,说明这两份续断亲缘关系很近,大理海东中药材种植基地续断种植成功。DL(W)-大理续断(云南丽江文笔山)与SZ-深紫续断(重庆南山金佛山古佛洞药池坝)相似性最小为0.6460,遗传距离则相反。4个不同产地的大理续断遗传距离在0.1044[DL(H)与DL(WS)]~0.1788[DL(W)与DL(H)]之间,平均遗传距离为0.1540。2个不同产地的川续断间的遗传距离为0.1487。根据NTSYS计算得出的遗传距离,用该软件进行聚类得到树状聚类图(图1)。
聚类图表明,当遗传相似系数为0.68时,10个续断样品分成了2个类群:DL-大理续断、ES-恩施续断、C(J)-川续断和RB-日本续断聚为一大类(Group);DT-大头续断和SZ-深紫续断聚为另一大类。当相似系数为0.79时,又分成了4个亚类群(Subgroup):第1亚类包括DL(L)、DL(W)、DL(H)、DL(WS)、C(D)和RB,4个品系的大理续断聚在一起,表明它们之间遗传亲缘关系比较近,基因交流比较频繁,但它们与RB(日本续断)和C(D)(川续断)也聚在一起,表明大理续断和日本续断以及川续断遗传亲缘关系比较近,有一定的基因交流;第2亚类包括ES(恩施续断)与C(J)(川续断),表明恩施续断与川续断亲缘关系较近;DT(大头续断)和SZ(深紫续断)各聚成第3和4大亚类,这表明大头续断和深紫续断两者亲缘关系也比较远,与大理续断、恩施续断以及日本续断亲缘关系也比较远,基因交流比较少。

表3 10份续断属植物间的遗传距离(D)(右上角)和相似系数(I)(左下角)
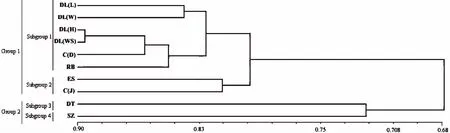
图1 10份续断属植物的UPGMA聚类图Fig.1 Dendrogram for ten accessions of Dipsacus by ISSR data using UPGMA method
3 讨 论
目前,国内多用形态学研究的方法对续断属植物进行了分类。但同一种植物在不同的环境条件下可能会发育出不同的形态或生理特征;形态接近,也可能是不同的基因型。分子标记则不受环境的影响,从植物的基因组出发,探讨其遗传多样性及遗传亲缘关系。艾铁民等对报道我国续断属植物共有18种(2栽培种),其中可供药用的有9种(包括2个栽培种)[4]。艾强等对四川贵州产地的川续断进行SRAP遗传分析,表明川续断居群的遗传多样性水平丰富,遗传变异主要存在居群内,居群遗传多样性主要受地理位置(海拔)和气候因素影响[15]。
本研究采用SRAP分子标记技术对来自湖北西部、重庆及云南大理和丽江等地的10份续断属植物的亲缘关系进行分析。结果表明,采用筛选得到的多态性较高的10对SRAP引物组合对10份续断样本进行PCR扩增,共扩出566条谱带,其中多态性条带为362条,平均多态性条 带百分率为64 %,结果表明该物种具有丰富的遗传变异。UPGMA聚类分析表明10个续断属植物分为2个大类和4个亚类,大头续断和深紫续断有一定的基因交流,有一定的亲缘关系,聚为一大类,与其它续断属植物间遗传差异较大;日本续断、恩施续断、大理续断和川续断的基因交流比较频繁,遗传亲缘关系较近,聚为另一大类,这与形态学分类基本一致[7]。供试的4个大理续断品系遗传亲缘关系较近,栽培种[DL(H)]与其他3个野生种遗传距离很近(0.1044~0.1788),表明大理续断的人工栽培比较成功,未对其遗传特性造成改变。笔者之前曾利用ISSR标记对同样的材料进行了遗传多样性及亲缘关系分析[16],与本研究结果基本一致,ISSR和SRAP标记对续断材料遗传多样性的分析结果呈极显著相关,相关性系数r=0.909(P<0.01),说明采用这2种分子标记对续断种质材料的遗传多样性分析具有较高的一致性和可信度。但SRAP标记的多态性小,遗传距离也有一定差异,表现在日本续断、恩施续断、川续断及大理续断遗传进化上,出现这种差异的原因可能是由于2种标记选取不同的引物及不同的扩增原理引起。ISSR标记是对两侧具有反向排列的SSR的一段DNA序列进行扩增,而SRAP标记则对内含子外显子区域进行特异扩增造成的,而且引物数量越多,多态性位点数也会随之增加,最终会使遗传距离发生变化[17-20]。冯学峰等[7]在对17种续断属植物的花粉的研究中发现大理续断和川续断的花粉形态相似,表明两者遗传亲缘关系比较近。目前也有人认为在续断属中没有大理续断和川续断的区别,它们都应该统称为川续断[21-22]。在本研究中结果也发现,川续断和大理续断同分为一个小类群,这从一定程度上支持了该观点,支持大理续断并入川续断,但仍需进行进一步研究如基因序列的测定来确定此观点的正确性。
[1]中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,1986:45-46.
[2]国家药典委员会. 中国药典[M]. 一部.北京:科学出版社,2010.
[3]陈虎彪, 艾铁明. 中国续断属药用植物资源[J]. 中国中药杂志, 1997, 22(11):649-652.
[4]艾铁民, 杨连菊, 冯学锋. 中国续断属新分类群[J]. 植物研究, 1995, 15(1):38-42.
[5]艾铁明. 中药续断原植物及续断属修订研究[J]. 中国医药学报, 1992, 7(2):22-24.
[6]Caputo P, Cozzolino S, Moretti A. Molecular phylogenetics ofDipsacaceaereveals parallel trends in seed dispersal syndromes[J]. Plant Syst.Evol., 2004, 246:163-175.
[7]冯学峰, 艾铁明, 徐海宁. 续断属花粉形态研究[J]. 中国中药杂志, 2000, 25(7):394-401.
[8]Li G, Quiros C F. Sequence-related amplified polymorphism(SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging inBrassica[J]. Theor Appl Gene, 2001, 103:455-461.
[9]Ferriol M, Picó B, Nuez F. Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers[J]. Theor Appl Genet, 2003, 107:271-282.
[10]Li G, Gao M, Yang B, et al. Gene for gene alignment between the Brassica and Arobidopsis genomes by direct transcriptome mapping[J]. Theor Appl Genet, 2003, 107:168-180.
[11]Sun Z, Wang Z, Tu J, et al. An ultradense genetic recombination map forBrassicanapus, consisting of 13551 SRAP markers[J]. Theor Appl Genet, 2007, 114:1305-1317.
[12]Doyle J J, Doyle J L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J]. Phytochem Bull, 1987,19:11-15.
[13]Li G, Quiros C F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction:its application to mapping and gene tagging inBrassica[J]. Theoretical and Applied Genetics, 2001, 103:455-461.
[14]陈明亮, 张冬林, 李志辉, 等.PAGE染色和条带回收方法的改进[J]. 中国林叶科技大学学报, 2007, 27 (6):163-165.
[15]艾 强, 周 涛, 江维克, 等. 川续断种质资源遗传多样性的SRAP 分析[J]. 中草药, 2013, 43 (15):2155-2159.
[16]李晓玲, 杨 进, 张 雄, 等. 10 份续断属植物亲缘关系的ISSR分析[J]. 植物研究, 2015, 35(3): 450-456.
[17]Li A, Ge S. Genetic variation and clonal diversity ofPsammochloavillosadetected by ISSR marker[J].Annals of Botany, 2001, 87(5):585-590.
[18]Pan D, Yu M W, Guo Y C, et al. Sequence-related amplified polymorphism(SRAP) of wild emmer wheat (Triticumdicoccoides) in Israel and its ecological association[J]. Biochemical Systematics and Ecology, 2010, 38(1): 1-11.
[19]郭凌飞. 应用ISSR和SRAP标记研究澳洲坚果的遗传多样性[D]. 海口: 海南大学, 2008.
[20]沈程文, 黄建安, 赵世浩, 等. 利用SRAP和ISSR标记分析广东茶树种质资源的遗传多样性[J]. 核农学报, 2010, 24 (5): 948-955.
[21]朱成兰. 云南常用中草药彩色图谱[M]. 海口: 海南出版社, 2003.
[22]太光聪, 朱继富, 郑启宏. 大理续断的特征特性及高产栽培技术[J]. 现代农业科技, 2011, 10:139-140.
(责任编辑 李山云)
Genetic Relations Analysis inDipsacusPlant Based on SRAP
LI Xiao-ling1,2, CHENG Sui-han1,2, YANG Jin1*
(1.College of Life Science and Pharmacy, Three Gorges University, Hubei Yichang 443002, China; 2.Engineering Research Center of Eco-environment in Three Gorges Reservoir, Ministry of Education, China Three Gorges University, Hubei Yichang 443002, China)
10 SRAP primer pairs were used to analyze the genetic relationship of 10Dipsacusmaterials. The results showed that 10 primer pairs produced 566 bands with average 56.6 bands by each primer, of which 362 bands were polymorphisms with average 36.2 bands, accounting for 64 %, and the length were 150-2000 bp, which indicated thatDipsacusplant had abundant genetic variation. According to the UPGMA cluster analysis, the ten pieces ofDipsacuscould be gathered into 2 big groups or 4 subgroups, DT and SZ were distance from any other accessions, and the genetic relationship between them was very close; DL, ES, RB, C(J) and C(D) had close relationships between them, and C(J) and C(D) belonged to the same subgroup, which supported thatD.daliensisT.M.Ai. Belonged toD.asperoidesC.Y.Cheng et T.M.Ai .
Dipsacusplant; SRAP; Genetic relation
1001-4829(2016)07-1554-05
10.16213/j.cnki.scjas.2016.07.010
2015-04-28
湖北省重点实验室开放基金“天然产物研究与利用”(2008NP09)
李晓玲(1973-),女,博士,副教授,主要从事三峡库区珍稀濒危植物保护的研究与利用,E-mail:lixiaolinggz@126.com,*为通讯作者。
S567.23
A
猜你喜欢
作物学报(2022年2期)2022-11-06 12:11:02
中国现代中药(2021年9期)2021-11-16 07:43:32
青年歌声(2021年2期)2021-03-05 09:02:08
音乐世界(2020年4期)2020-09-10 20:57:26
民族音乐(2018年5期)2018-11-17 08:20:00
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
广东农业科学(2017年5期)2017-08-29 10:37:26
Coco薇(2017年8期)2017-08-03 20:58:26
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
中国光学(2015年1期)2015-06-06 18:30:20
