
多巴胺受体基因多态性及其与鸡繁殖性状的相关性
2016-12-22 09:19:15朱丽莉徐龙鑫伍革民陶宇航唐继高顾永江
西南农业学报 2016年7期
朱丽莉,徐龙鑫,伍革民*,陶宇航,韩 雪,李 亮,唐继高,顾永江
(1.贵州省畜牧兽医研究所,贵州 贵阳 550005;2.贵州省凯里市畜牧兽医局,贵州 凯里 556000)
多巴胺受体基因多态性及其与鸡繁殖性状的相关性
朱丽莉1,徐龙鑫1,伍革民1*,陶宇航1,韩 雪1,李 亮1,唐继高1,顾永江2
(1.贵州省畜牧兽医研究所,贵州 贵阳 550005;2.贵州省凯里市畜牧兽医局,贵州 凯里 556000)
为探明多巴胺(Dopamine,DA)受体基因与鸡繁殖性状的关联性,找出可作为鸡繁殖性状的重要标记,选用贵州省地方鸡种荔波瑶山鸡,采用PCR测序法对5种DA受体基因(DRD1~DRD5)的多态位点进行检测,并进行与繁殖性状的关联分析。结果表明:DRD1基因有1073(G→A)1个多态位点,分别与300日龄产蛋量、首次就巢日龄呈极显著相关(P<0.01);DRD2基因上有15个多态位点,分别与繁殖性状存在显著或极显著相关,其中14个位点分布于各内含子上,仅有4260(G→C)1个位点位于第6外显子,该位点与开产日龄、300日龄产蛋量和首次就巢日龄呈显著相关(P<0.05);DRD3基因11720(C→G)与300日龄产蛋量存在显著相关(P<0.05),该位点CC型个体在试验期间无就巢现象;DRD4基因5365(C→T)与开产日龄存在显著相关(P<0.05),5546(G→A)与首次就巢日龄和首次就巢持续时间存在显著相关(P<0.05)。以上多态位点可作为与相关繁殖性状有显著或极显著影响指标的辅助选择标记。
多巴胺受体基因;基因多态位点;荔波瑶山鸡;繁殖性状
地方鸡品种资源因具有耐粗饲、抗逆性强、适应性强、肉质鲜美细嫩等特点,在我国优质鸡育种中发挥着重要作用,但又受限于地方鸡的重要经济性能偏低,尤其是开产日龄、产蛋量、就巢性等繁殖性状偏低,影响其养殖效益。繁殖性状是多基因控制的复杂性状,且遗传力较低,采用传统育种方法提高这些性状进展缓慢,因此可利用分子标记辅助选择。有研究表明,许多候选基因都与繁殖性状密切关系,如NPY基因、PRL基因、FSH基因、VIPR基因等[1-4],多巴胺(Dopamine,DA)也是这一性状中的重要因子。
DA是中枢神经系统中主要的神经递质,对人及其他动物神经系统、内分泌和繁殖起着重要的调节作用[5-7],具有强烈抑制促性腺激素释放以及刺激促生长激素分泌的作用。作为肽类激素,DA需通过其受体介导才能发挥生物学效应。目前,鉴定出的DA受体基因有5种,按生理功能类型分为两大类:D1和D5被统称为D1样受体,无内含子;D2、D3和D4被统称为D2样受体。有研究表明,鸟类DA通过兴奋多巴胺D2受体(DRD2)抑制垂体中VIP 和PRL 的分泌,从而对鸟类的繁殖性状起调控作用[8-9];多巴胺D1受体(DRD1)和DRD2基因某些多态位点与禽类的生产性能呈显著或极显著相关[10-11]。而关于多巴胺D3、D4、D5受体与禽类生产性能的相关研究鲜见报道。为探明这5种DA受体基因与鸡繁殖性状的关联性,笔者选用贵州省地方鸡种荔波瑶山鸡进行该研究,以期找出可作为鸡繁殖性状的重要标记。
1 材料与方法
1.1 荔波瑶山鸡
试验鸡为120日龄荔波瑶山鸡母鸡,60只,选自贵阳市乌当区百宜乡洛坝村种鸡场,均采用常规饲养管理,个体笼养,群体一致性较好,性能较为稳定。翅下静脉采血1.0~1.5 mL,加EDTA-Na2抗凝,-20 ℃保存。
1.2 性状测定
120 日龄母鸡转入产蛋舍单只笼养至300 日龄,测定开产日龄、开产体重、就巢性等繁殖性状,测定方法按照NY/T 823-2004[12]进行。
1.3 基因组DNA提取及引物设计
采用TIANamp Blood DNA Kit试剂盒提取血样DNA。根据GenBank中鸡的DA受体基因序列合成18对引物(由天根生化科技有限公司合成),检测5种DA受体基因在荔波瑶山鸡60只个体中的基因型遗传变异情况。检测的位点、引物序列、扩增长度以及检测方法见表1。
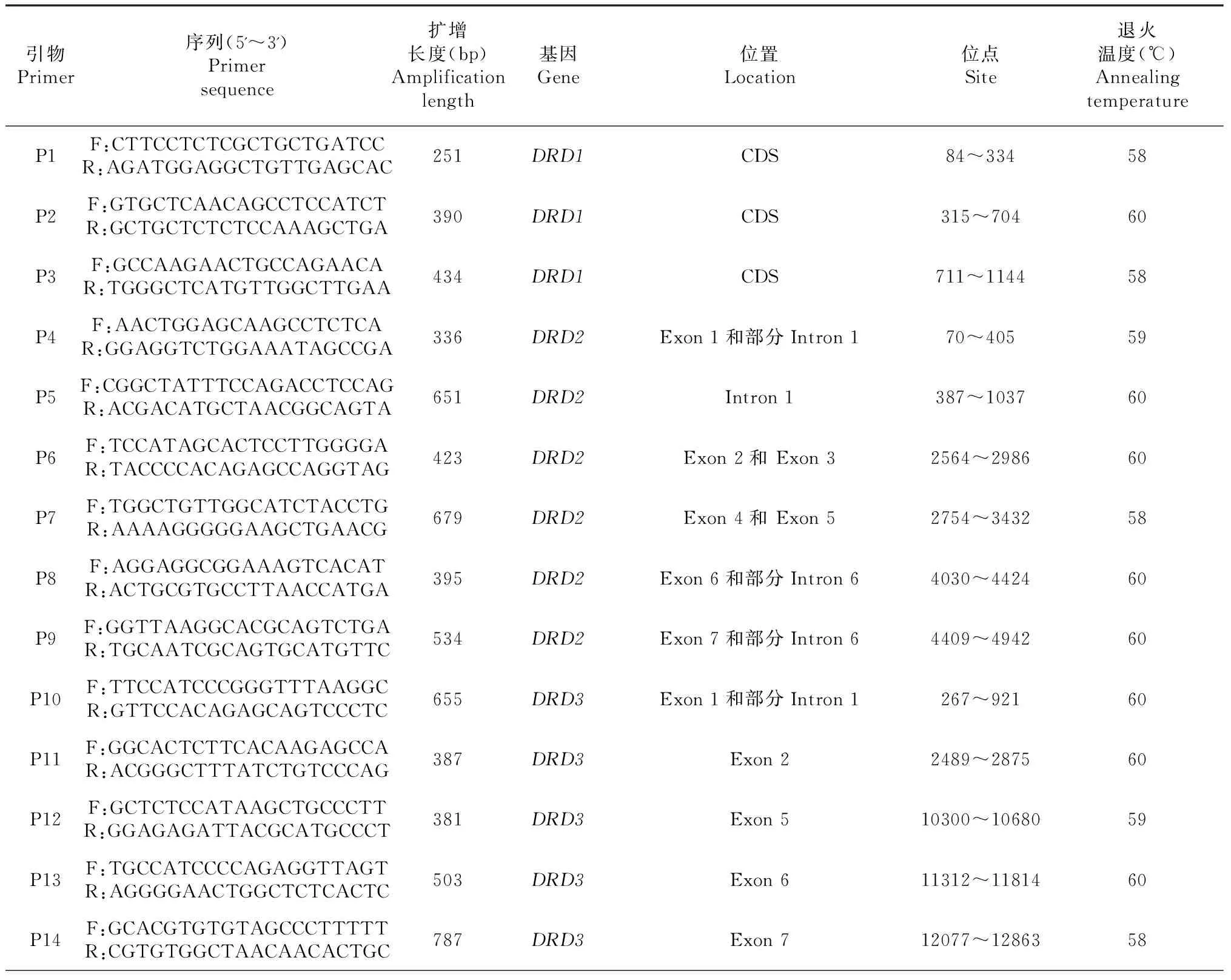
表1 荔波瑶山鸡基因型检测的18对引物信息
续表1 Continued table 1
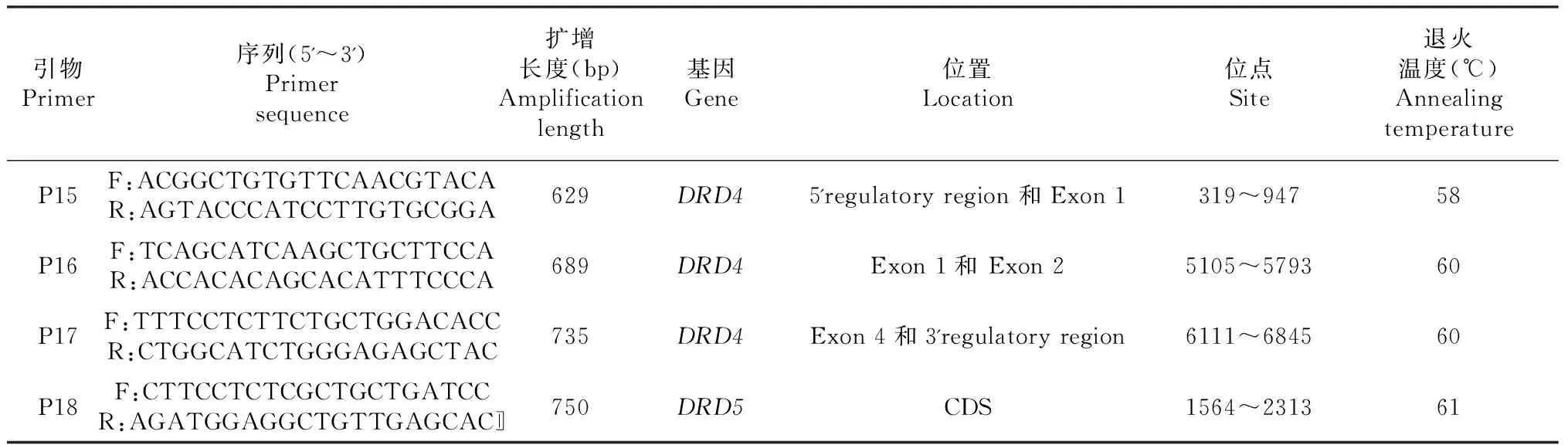
引物Primer序列(5'~3')Primersequence扩增长度(bp)Amplificationlength基因Gene位置Location位点Site退火温度(℃)AnnealingtemperatureP15F:ACGGCTGTGTTCAACGTACAR:AGTACCCATCCTTGTGCGGA629DRD45'regulatoryregion和Exon1319~94758P16F:TCAGCATCAAGCTGCTTCCAR:ACCACACAGCACATTTCCCA689DRD4Exon1和Exon25105~579360P17F:TTTCCTCTTCTGCTGGACACCR:CTGGCATCTGGGAGAGCTAC735DRD4Exon4和3'regulatoryregion6111~684560P18F:CTTCCTCTCGCTGCTGATCCR:AGATGGAGGCTGTTGAGCAC〛750DRD5CDS1564~231361
1.4 PCR扩增及测序
PCR反应体系:DNA模板1 μl,上下游引物各1 μl,2×TaqPCR MasterMix 试剂10 μl,加ddH2O至20 μl。PCR反应参数:95 ℃预变性6 min;95 ℃变性30 s,特定温度(表1)退火30 s,72 ℃延伸45 s,35个循环;最后72 ℃延伸10 min。PCR产物用1 %琼脂糖凝胶电泳检测,委托诺赛基因组研究中心有限公司测序,即利用30只个体组成的混合DNA池作DNA模板,进行PCR扩增并测序,分析序列多态性;针对存在多态位点的引物,逐一对60个样本进行单独测序。
1.5 统计分析
使用DNAstar软件对测序结果进行校正,进入基因库网站采用BLAST分析序列SNPs,采用DNAMAN软件分析SNPs变异导致蛋白质编码变异的情况。以EXCEL软件统计60只鸡DA受体基因分型,利用SPSS 18.0软件采用单因子方差分析方法分析基因型与繁殖性状的相关性。
2 结果与分析
2.1 多态位点和遗传多样性
经检测,18对引物共检测出28个多态位点(表2),其中DRD5基因的引物未检出多态位点;各多态位点的基因型频率在0.06~0.93(表3),等位基因频率在0.03~0.95,多肽信息含量在0.12~0.58。DRD1基因位置1073导致该蛋白质第355位脯氨酸发生同义突变。DRD2基因18个多态位点仅有4260位于第6外显子,导致该蛋白质第304位氨基酸发生错义突变(赖氨酸K→天门冬酰胺N)(AAG→AAC),其余17个位点均分布于各内含子上。DRD3基因位置11720导致该蛋白质第333位氨基酸发生错义突变(丝氨酸S→半胱氨酸C)(UCU→UGU)。DRD4基因位置5365导致该蛋白质187位的异亮氨酸发生同义突变(AUC→AUU),位置5546导致该蛋白质第248位氨基酸发生错义突变(甘氨酸G→丝氨酸S)(GGC→AGC)。

表2 荔波瑶山鸡DA受体基因的多态位点
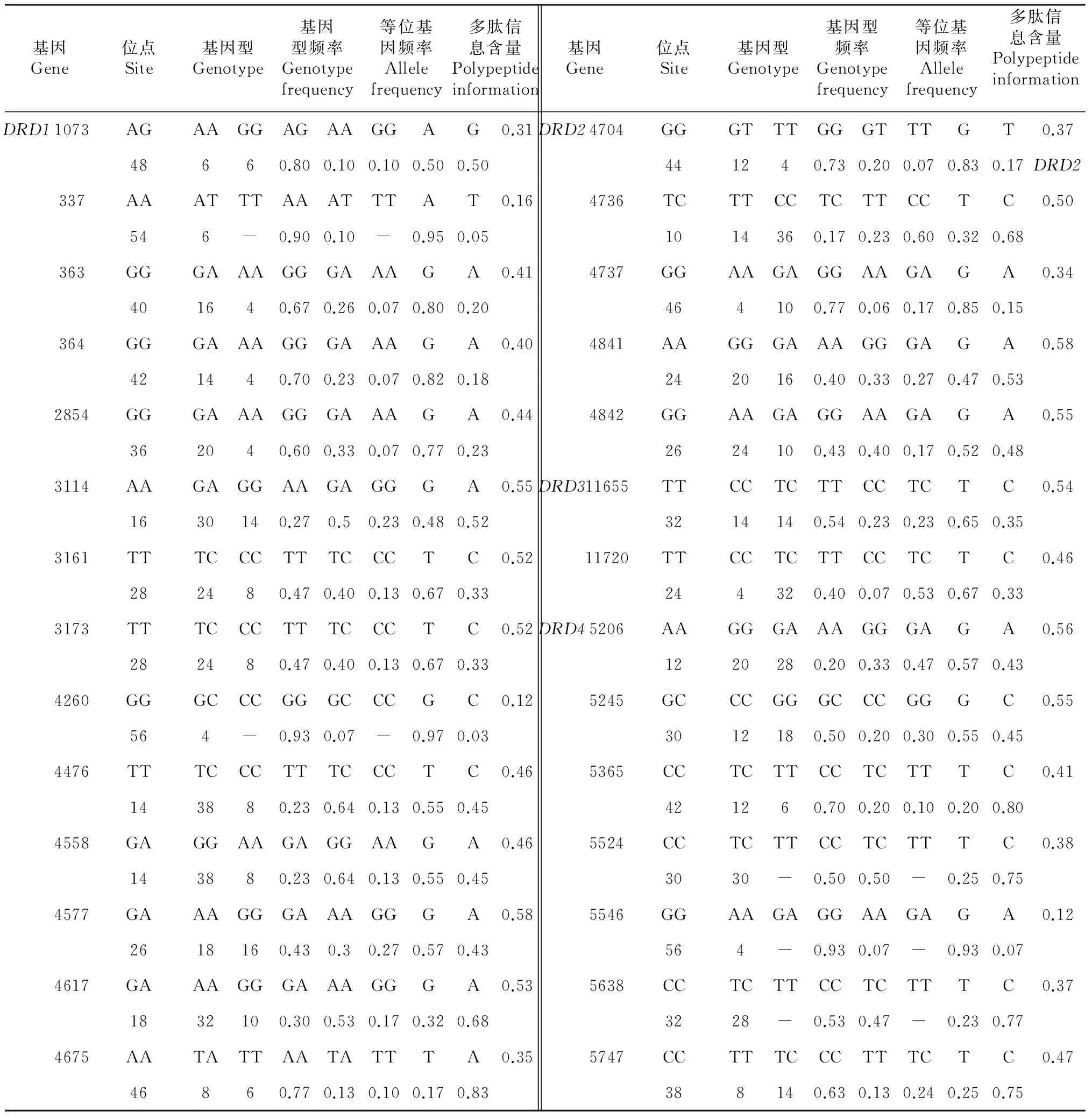
表3 DA受体基因的遗传多态性
2.2 多态位点与繁殖性状的相关性
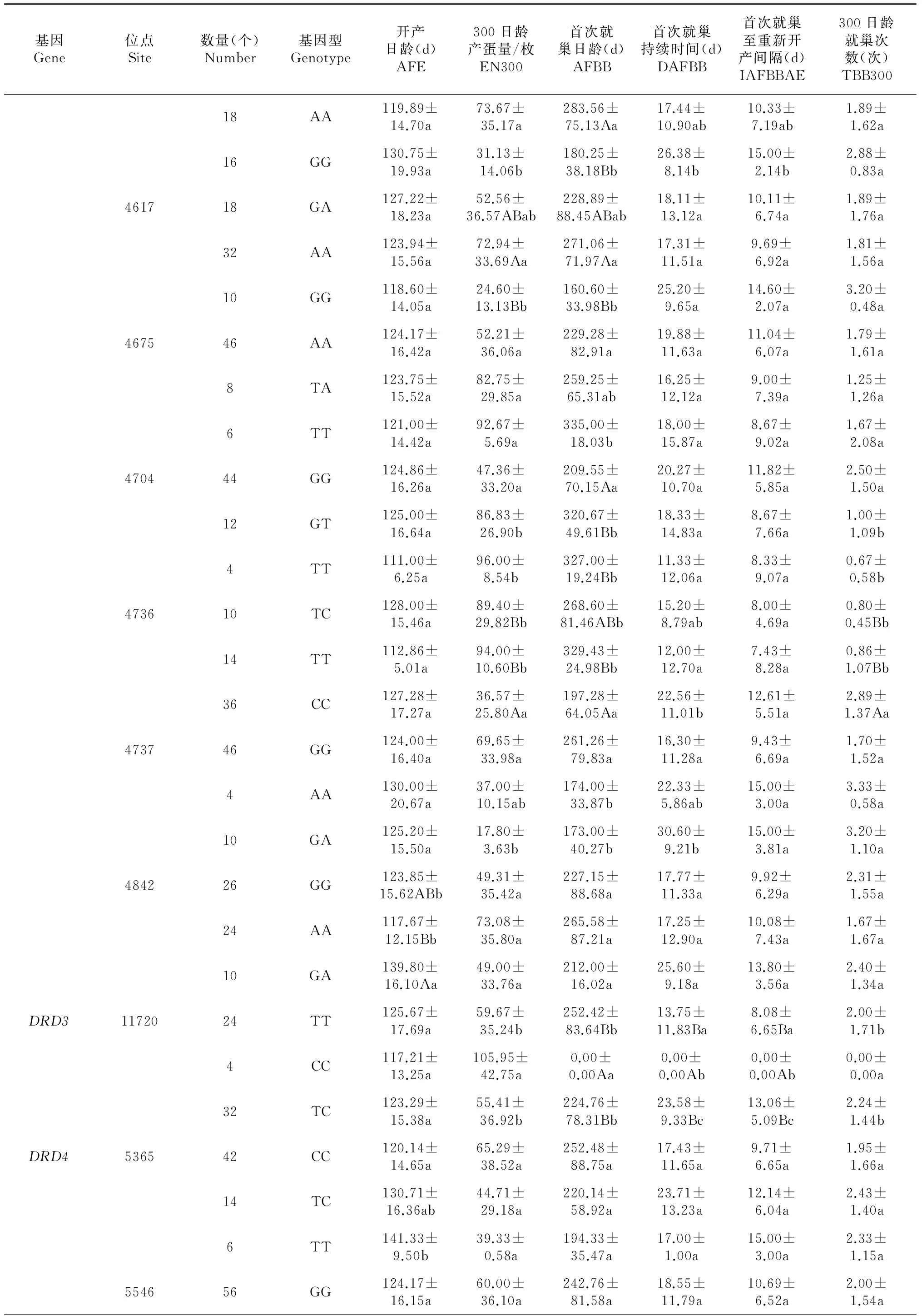
2.2.1DRD1基因 该基因300日龄产蛋量AA型与GG型间差异极显著;首次就巢日龄AA型与AG型间差异显著,AA型与GG型间差异极显著。
2.2.2DRD2基因 该基因有18个多态位点,其中开产日龄基因型间差异显著或极显著的位点有3个,分别是364、4260和4842;300日龄产蛋量基因型间差异显著或极显著的位点有11个,分别是337、364、3173、4260、4476、4558、4577、4617、4704、4736和4737;首次就巢日龄基因型间差异显著或极显著的位点有12个,分别是337、364、3173、4260、4476、4558、4577、4617、4675、4704、4736和4737;首次就巢持续时间基因型间差异显著或极显著的位点有5个,分别是363、4476、4577、4736和4737;首次就巢至重新开产间隔基因型间差异显著或极显著的位点有4个,分别是363、364、4476和4577;300日龄就巢次数基因型间差异显著或极显著的位点有7个,分别是364、2854、3173、4476、4558、4704和4736。
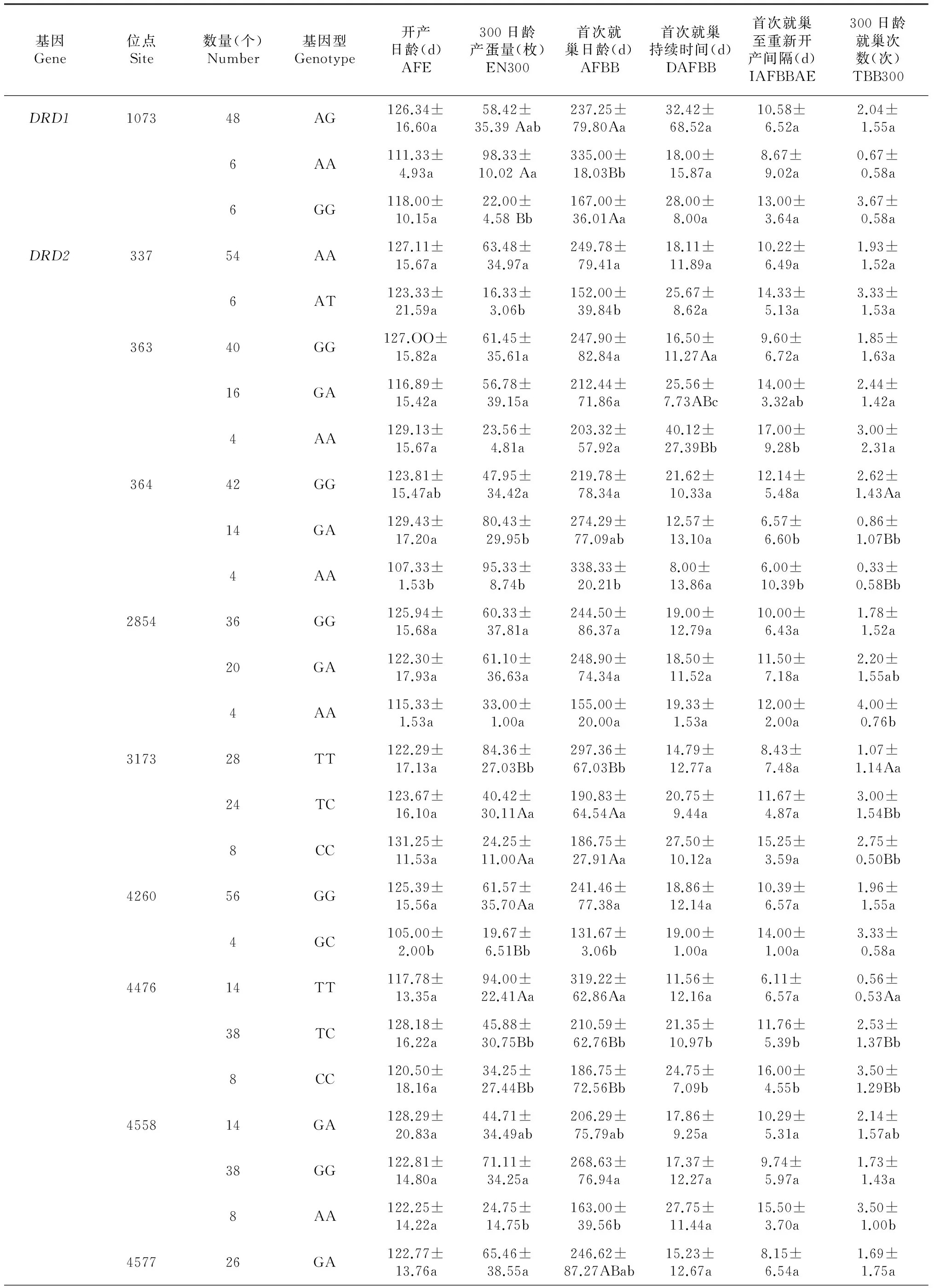
表4 DRD基因多态位点与繁殖性状的相关性
续表4 Continued table 4
基因Gene位点Site数量(个)Number基因型Genotype开产日龄(d)AFE300日龄产蛋量/枚EN300首次就巢日龄(d)AFBB首次就巢持续时间(d)DAFBB首次就巢至重新开产间隔(d)IAFBBAE300日龄就巢次数(次)TBB30018AA119.89±14.70a73.67±35.17a283.56±75.13Aa17.44±10.90ab10.33±7.19ab1.89±1.62a16GG130.75±19.93a31.13±14.06b180.25±38.18Bb26.38±8.14b15.00±2.14b2.88±0.83a461718GA127.22±18.23a52.56±36.57ABab228.89±88.45ABab18.11±13.12a10.11±6.74a1.89±1.76a32AA123.94±15.56a72.94±33.69Aa271.06±71.97Aa17.31±11.51a9.69±6.92a1.81±1.56a10GG118.60±14.05a24.60±13.13Bb160.60±33.98Bb25.20±9.65a14.60±2.07a3.20±0.48a467546AA124.17±16.42a52.21±36.06a229.28±82.91a19.88±11.63a11.04±6.07a1.79±1.61a8TA123.75±15.52a82.75±29.85a259.25±65.31ab16.25±12.12a9.00±7.39a1.25±1.26a6TT121.00±14.42a92.67±5.69a335.00±18.03b18.00±15.87a8.67±9.02a1.67±2.08a470444GG124.86±16.26a47.36±33.20a209.55±70.15Aa20.27±10.70a11.82±5.85a2.50±1.50a12GT125.00±16.64a86.83±26.90b320.67±49.61Bb18.33±14.83a8.67±7.66a1.00±1.09b4TT111.00±6.25a96.00±8.54b327.00±19.24Bb11.33±12.06a8.33±9.07a0.67±0.58b473610TC128.00±15.46a89.40±29.82Bb268.60±81.46ABb15.20±8.79ab8.00±4.69a0.80±0.45Bb14TT112.86±5.01a94.00±10.60Bb329.43±24.98Bb12.00±12.70a7.43±8.28a0.86±1.07Bb36CC127.28±17.27a36.57±25.80Aa197.28±64.05Aa22.56±11.01b12.61±5.51a2.89±1.37Aa473746GG124.00±16.40a69.65±33.98a261.26±79.83a16.30±11.28a9.43±6.69a1.70±1.52a4AA130.00±20.67a37.00±10.15ab174.00±33.87b22.33±5.86ab15.00±3.00a3.33±0.58a10GA125.20±15.50a17.80±3.63b173.00±40.27b30.60±9.21b15.00±3.81a3.20±1.10a484226GG123.85±15.62ABb49.31±35.42a227.15±88.68a17.77±11.33a9.92±6.29a2.31±1.55a24AA117.67±12.15Bb73.08±35.80a265.58±87.21a17.25±12.90a10.08±7.43a1.67±1.67a10GA139.80±16.10Aa49.00±33.76a212.00±16.02a25.60±9.18a13.80±3.56a2.40±1.34aDRD31172024TT125.67±17.69a59.67±35.24b252.42±83.64Bb13.75±11.83Ba8.08±6.65Ba2.00±1.71b4CC117.21±13.25a105.95±42.75a0.00±0.00Aa0.00±0.00Ab0.00±0.00Ab0.00±0.00a32TC123.29±15.38a55.41±36.92b224.76±78.31Bb23.58±9.33Bc13.06±5.09Bc2.24±1.44bDRD4536542CC120.14±14.65a65.29±38.52a252.48±88.75a17.43±11.65a9.71±6.65a1.95±1.66a14TC130.71±16.36ab44.71±29.18a220.14±58.92a23.71±13.23a12.14±6.04a2.43±1.40a6TT141.33±9.50b39.33±0.58a194.33±35.47a17.00±1.00a15.00±3.00a2.33±1.15a554656GG124.17±16.15a60.00±36.10a242.76±81.58a18.55±11.79a10.69±6.52a2.00±1.54a
续表4 Continued table 4

基因Gene位点Site数量(个)Number基因型Genotype开产日龄(d)AFE300日龄产蛋量/枚EN300首次就巢日龄(d)AFBB首次就巢持续时间(d)DAFBB首次就巢至重新开产间隔(d)IAFBBAE300日龄就巢次数(次)TBB3004AA122.67±3.79a19.00±3.46a165.00±38.74b33.00±4.36b9.33±2.52a4.00±1.00a554738CC120.11±13.50a65.79±38.06a251.84±87.28a17.37±10.88a9.74±6.36a1.95±1.61a6TT145.33±11.59b38.33±2.08a193.33±34.15a22.67±9.87a15.33±3.06a2.33±1.15a12TC129.83±17.74ab44.50±31.96a225.00±62.99a24.67±14.22a12.17±6.62a2.33±1.51a
注:同基因同位点不同基因型间同列不同大写字母表示差异极显著(P<0.01),不同小写字母表示差异显著(P<0.05),)。与所有性状均不显著的位点在本表中未列出。 Note: Different capital and lowercase letters in the same column indicate significance of difference atP<0.01 andP<0.05 level respectively. The sites insignificantly related to all traits are not listed in the table.
2.2.3DRD3基因 试验期间该基因CC型个体未就巢。TT型和TC型间开产日龄、300日龄产蛋量、首次就巢日龄间差异均不显著,首次就巢持续时间、首次就巢至重新开产间隔、300日龄就巢次数间差异显著。
2.2.4DRD4基因 该基因5365位点CC型与TT型间开产日龄差异显著;5546位点GG型与AA型首次就巢日龄、首次就巢持续时间差异均显著;5747位点CC型与TT型开产日龄间差异极显著。
3 结论与讨论
(1)鸡DRD1基因定位于13号染色体上,无内含子。有关DRD1基因与家禽生长性能的相关性研究表明,由C+765T、C+1011T与G+1065A位点组成的单倍型与鸡77日龄冠高显著相关[11];鸭DRD1基因CDS区序有5个SNPs位点与繁殖性状、脂肪代谢显著相关[13];鸭DRD1基因CDS区有2个SNPs位点与活重、体斜长和胸深等呈显著相关14]。研究表明,荔波瑶山鸡DRD1基因位置1073(G→A)与鸡300日龄产蛋量、首次就巢日龄均呈显著相关,其中AA型个体的300日龄平均产蛋量比GG型多76枚 (P<0.01),AA型个体的首次就巢日龄分别比AG型和GG型晚98 d (P<0.05)和168 d(P<0.01)。
(2)鸡DRD2定位于24号染色体上,有7个外显子。有研究发现,DRD2基因5’调控区上的突变位点及不同基因型与宁都三黄鸡就巢率显著相关,还与开产日龄、300 日龄总产蛋数、91 日龄体重和冠高等性状也呈显著或极显著相关[10-11];DRD2基因GG型个体与淮南麻黄鸡开产日龄和28 周龄产蛋数呈显著相关[15];DRD2基因位置SNP 962(C→T)与鸡38 周龄产蛋量和平均蛋重呈显著或极显著相关[16]。研究表明,荔波瑶山鸡DRD2基因的18个变异位点,有15个变异位点与鸡繁殖性状存在显著或极显著相关,其中位点4260(G→C)位于第6外显子,该位点与开产日龄、300日龄产蛋量和首次就巢日龄呈显著相关(P<0.05),GG型与GC型间差异显著(P<0.05);其余14个位点均分布于各个内含子1、3、4和6上,与不同性状存在显著或极显著相关。内含子含有基因表达的调控元件,能起到类似增强子或其他顺式调控元件的功能,从而对基因表达有着重要的调控作用[17-18];内含子的功能有利于物种的进化选择[19]。荔波瑶山鸡14个内含子是否调控其繁殖性能主效基因的表达,进而影响鸡的繁殖性能有待进一步研究验证。
(3)鸡DRD3定位于1号染色体上,有7个外显子。DRD3受体常表达于突触后神经元,在情感、运动和认知感觉的信息加工过程中发挥重要作用。张宝等[20]研究表明,敲除DRD3基因并不影响小鼠的繁殖能力。但关于鸡DRD3基因多态性、表达调控及其与经济性状的关联性和进化等方面的研究甚少。研究表明,荔波瑶山11720(C→G)变异位点的CC型个体在试验期间未表现出就巢性状,其300日龄产蛋量显著高于TC和TT型(P<0.05)。
(4)鸡DRD4定位于5号染色体上,有4个外显子。Van ToI H H M等[21]研究表明,DRD4基因的多态现象与猎奇人格有关,从而引发了一些不同寻常的性格倾向。范彩云等[22]首次克隆马DRD4基因发现其存在于6个品种马之间的多态位点。但关于鸡的DRD4基因多态性、表达调控及其与经济性状的关联和进化等方面的研究甚少。研究表明,荔波瑶山鸡DRD4基因有7个变异位点,其中5365(C→T)、5546(G→A)和5747(C→T)位点与繁殖性状显著或极显著相关。5365位点CC型的开产日龄显著早于TT型(P<0.05);5546位点GG型的首次就巢日龄和首次就巢持续时间与AA型间差异显著(P<0.05);5747位点CC型的开产日龄与TT型间差异极显著(P<0.01)。
(5)综上所述,贵州地方鸡荔波瑶山鸡DA受体基因上有28个多态位点,其中DRD1上有1个多态位点分别对母鸡300日龄产蛋量、首次就巢日龄影响显著或极显著;DRD2上有15个多态位点对母鸡的繁殖性状影响显著或极显著;DRD3基因有1个多态位点对母鸡300日龄产蛋量、首次就巢日龄、首次就巢持续时间、首次就巢至重新开产间隔和300日龄就巢次数影响显著或极显著;DRD4基因有3个多态位点分别对母鸡开产日龄、首次就巢日龄和首次就巢持续时间影响显著或极显著。该结果为利用分子标记辅助选择提高荔波瑶山鸡的繁殖性能奠定了一定基础。
[1]Dunnic I C,Miao Y W,Morris A,et a1.A study of association between genetic markers in candidate genes and reproductive traits in one generation of a commercial broiler breeder hen population[J].Heredity,2004,92(2):128-134.
[2]Cui J X,Xu H L,Liang Y,et a1.Association of polymorphisms in the promoter region of chicken prolactin with egg production[J].Poult Sci,2006,85(1):26-31.
[3]施振旦,郭日红,邢光东.催乳素调控禽类繁殖活动的研究现状与展望[J].中国家禽,2012,34(4):1-4.
[4]周 敏,梁菲菲,饶友生,等.VIPR-1基因12个多态位点与鸡早期产蛋性状的相关性[J].畜牧兽医学报,2008,39(9):1147-1152.
[5]Margaret R S,Michelle A D,Patricia P,et al.Variant alleles of the D2 do pamine receptor gene and obesity[J]. Nutri Res,2000,20(3):371-380.
[6]Kampman O,Anttila S,Illi A,et al. Dopamine receptor D2-141C Insertion/Deletion polymorphism in a Finnish population with schizophrenia[J].Psychiatry Res,2003,121(1):89-92.
[7]Hansen K A,Zhang Y,Colver R,et al.The dopamine receptor D2 genotype is associated with hyper prolactinemia[J]. Fertil Steril,2005,84 (3):711-718.
[8]Youngren O M,Pitts G R,Phillips R E,et al. The stimulatory and inhibitory effects of dopamine on prolactin secretion in the turkey[J]. Gen Comp Endocrinol,1995,98(1):111-117.
[9]Youngren O M,Pitts G R,Phillips R E,et al. Dopaminergic control of prolactin secretion in the turkey[J].Gen Comp Endocrinol,1996,104(2):225-230.
[10]徐海平.多巴胺受体2基因对家鸡就巢性状的遗传效应[D].广州:华南农业大学,2008.
[11]徐海平,周 敏,罗成龙,等.DRD1及DRD2基因多态性与鸡冠高和体重的相关性[J].中国农业大学学报,2010,15(2):64-70.
[12]NY/T 823-2004 家禽生产性能名词术语和度量统计方法[M].北京:中国农业出版社,2006.
[13]王 翠,李世军,李 闯,等.鸭多巴胺受体1(DRD1)基因克隆与多态性检测[A]∥中国畜牧兽医协会第十五次全国家禽学术讨论会论文集[C].广州:广东省农业科学院畜牧研究所,2011.
[14]潘兰兵,张依裕,林家栋,等.鸭多巴胺受体D1基因CDS区多态性与生长性状的关联性分析[J].中国畜牧兽医,2014,41(1):155-158.
[15]刘 伟,李绍全,詹 凯,等.淮南麻黄鸡FSH-β和DRD2基因多态性与早期产蛋性能关系分析[J].中国家禽,2013,35(5):9-12.
[16]朱丽莉,陶宇航,顾永芬,等.荔波瑶山鸡DRD2基因多态与早期产蛋性能的关系[J].黑龙江畜牧兽医,2015(7):94-96.
[17]Haigh C L,Wright J A,Brown D R. Regulation of prionprotein expression by noncoding regions of the Prap gene[J].J Mol Biol,2007368(4):915-927.
[18]Li K,Cai R,Dai B B,et a1.SATBI regulates SPARC expression in K562 cell line through binding to a specific sequence in the third intron[J].Biochem Biophys Res Commun,2007,356(1):6-12.
[19]Deutsch M,Long M. Intron-exon structures of eukaryotic model organisms[J].Nucleic Acids Res,1999,27(15):3219-3228.
[20]张 宝,魏曙光,张洪波,等.DRD3基因敲除鼠饲养繁殖及基因型鉴定[J].安徽农业科学,2013,41(26):10657-10658.
[21]Van ToI H H M,Wu C M,Guan H C,et a1.Multiple dopamine D4 receptor variants in the human population[J].Nature,1992,358:149-152.
[22]范彩云,芒 来,程建波,等.6个品种马DRD4基因克隆与序列比较分析[J].畜牧兽医学报,2007,38(8):866-871.
(责任编辑 冯 卫)
Polymorphism of Dopamine Receptor Gene and Correlation between Dopamine Receptor Gene and Chicken Reproductive Trait
ZHU Li-li1, XU Long-xin1, WU Ge-min1*,TAO Yu-hang1,HAN Xue1,LI Liang1,TANG Ji-gao1,GU Yong-jiang2
(1.Guizhou Institute of Animal Husbandry and Veterinary, Guizhou Guiyang 550005,China; 2.Kaili Bureau of Animal Husbandry and Veterinary, Guizhou Kaili 556000, China)
Libo Yaoshan Chicken, a Guizhou local chicken variety, was used to detect polymorphic site of five DA receptor genes (DRD1-DRD5) by PCR sequencing and the relationship between these DA receptor genes and chicken reproductive traits was analyzed to discuss the relationship between Dopamine (DA) receptor gene and chicken reproductive traits and find out the important markers for chicken reproductive traits. Results: 1073 (G→A) ofDRD1 gene is related to egg number of 300 days (EN300) and age at first broodiness behavior (AFBB) very significantly. More than 15 polymorphic sites ofDRD2 gene are related to chicken reproductive traits significantly or very significantly. 14 sites of 15 sites distribute to different introns. The site of 4260 (G→C) is located in exon-6, which is related to age of first egg (AFE), EN300 and AFBB significantly. 11720 (C→G) ofDRD3 gene is related to EN300 significantly. The individuals with CC type and 11720 (C→G) are no broodiness behavior. 5365 (C→T) ofDRD4 gene is related to AFBB significantly and 5546 (G→A) ofDRD4 gene is related to AFBB and duration of age at first broodiness behaviour (DAFBB) significantly. The above polymorphic sites are used as the assistant selection markers to influence the related reproductive traits significantly or very significantly.
Dopamine receptor; Polymorphic site; Libo Yaoshan chicken; Reproductive trait
1001-4829(2016)07-1735-08
10.16213/j.cnki.scjas.2016.07.042
2016-01-13
贵州省联合基金项目“瑶山鸡ONECUTI基因的CDS区克隆、差异表达及与生产性状关联分析”[黔科合LH字(2015)7067];贵州省科技基金项目“瑶山鸡就巢性关键基因筛选”[黔科合J字(2014)2102];贵州省农业动植物育种项目“瑶山鸡新品系的选育”[黔农育专字(2014)008];贵州省农业科学院2014年博士基金“瑶山鸡STAT5变异检测及与产蛋性状的关联分析 ”
朱丽莉(1984-),女,助理研究员,硕士,从事家禽遗传育种研究,E-mail:zhuerms@163.com,*为通讯作者。
S831
A
猜你喜欢
今日农业(2022年15期)2022-09-20 06:54:16
Asian Herpetological Research(2021年4期)2021-12-30 08:24:20
歌海(2021年3期)2021-07-25 02:30:48
公民与法治(2020年18期)2020-10-28 08:48:18
北方音乐(2017年16期)2017-10-21 02:07:42
夜郎文学(2017年2期)2017-09-26 06:29:38
乡村地理(2017年4期)2017-09-18 02:54:20
歌海(2016年6期)2017-01-10 01:35:40
兽医导刊(2016年12期)2016-05-17 03:51:55
饲料博览(2015年12期)2015-04-04 04:28:36
