
江南油杉球果种子特性种源变异分析
2016-12-21 01:33王润辉胡德活郑会全朱积余
中南林业科技大学学报 2016年2期
王润辉 ,胡德活,郑会全,晏 姝 ,蒋 燚 ,朱积余
(1. 广东省林业科学研究院,广东 广州 510520;2. 广西壮族自治区林业科学研究院,广西 南宁 530002)
江南油杉球果种子特性种源变异分析
王润辉1,胡德活1,郑会全1,晏 姝1,蒋 燚2,朱积余2
(1. 广东省林业科学研究院,广东 广州 510520;2. 广西壮族自治区林业科学研究院,广西 南宁 530002)
江南油杉是我国珍贵用材树种,其资源分布日益减少。为了更有效的评价江南油杉种质资源的遗传多样性,选择广东、江西境内的五个种源地开展球果和种子形态特征差异性的研究。方差分析结果表明,球果和种子的形态性状差异在种源间均达极显著水平,多重比较进一步证明种源间差异的真实存在;球果和种子形态性状间的相关分析显示,球果短径与三个种子性状间均存在极显著地表型和遗传相关。主成分分析和聚类分析等多元统计分析揭示了五个种源的相似性情况,与行政区划不同,广东的南雄种源与江西的大余种源和崇义种源间的相似性大,这可能与三地的地理位置相邻空间隔离较小有关。
球果;种子;遗传变异;江南油杉
江南油杉Keteleeria cyclolepisFlous.是我国特有的珍贵针叶用材树种,主要分布于福建、浙江、江西、广东、广西、湖南、贵州、云南等省区[1-2],是松科油杉属的常绿大乔木。天然或次生林分中生长的江南油杉,高可达40 m,胸径达100 cm以上,有很好的用材价值;同时其枝叶茂密浓绿、树形优美,具有良好的观赏价值,于是也成为优良的园林绿化树种。江南油杉含有胶质,广东粤北南雄等地区早年常用其树根、树叶和树枝蒸煮胶液用于造纸,说明其有多种用途,潜在的利用价值有待深入开发。由于江南油杉树形高大,易受人为砍伐破坏,导致江南油杉的自然分布数量有限,天然群落日渐稀少,资源已经处于濒危状态,宜加大力度开展繁殖和保护研究。2000年福建省将其列为重点保护树种[3],目的在于通过人为措施来挽救和保护该树种。广东、江西都是江南油杉的自然分布区,但由于遭到一定程度的破坏其分布地区和数量极为有限,据调查两省只在少数较为边远山区有江南油杉的分布。
植物的表型性状是环境与遗传因素共同作用的结果,表型多样性是遗传多样性研究的重要内容,也是遗传多样性与环境多样性的综合体现[4-5]。顾万春[6]提出表型多样性研究主要通过科学有效地采样,合理的数学统计方法,采用总体形态性状、繁殖器官性状、生理生化性状等较稳定的性状,来揭示群体的遗传规律、变异大小,以客观评价其遗传多样性。根据球果、种子等繁殖器官较为稳定的性状,臧润国等[7]、王小平等[8]、孙玉玲等[9]、王旭军[10]、梁晓静[11]和、Gil等[12]分别研究了天山云杉、白皮松、秦岭冷杉、红榉、肉桂和加纳利松Pinus canariensis等群体的表型多样性,指出了这些珍贵树种群体形态变异规律,为这些树种的保护和利用提出了合理的意见和建议。
为了有效的保护和利用广东、江西两省的江南油杉种质资源,本研究在两省调查收集种源、家系遗传材料,开展不同种源和家系的球果和种子特性的表型多样性及遗传变异规律分析。旨在挖掘表型数据中隐藏的信息,为该树种的繁殖、保护和利用提供表型变异的理论依据。
1 材料与方法
1.1 材料来源
在查阅文献的基础上,2012年分别在广东、江西两省的江南油杉天然分布地区开展分系采种,详见下表1。
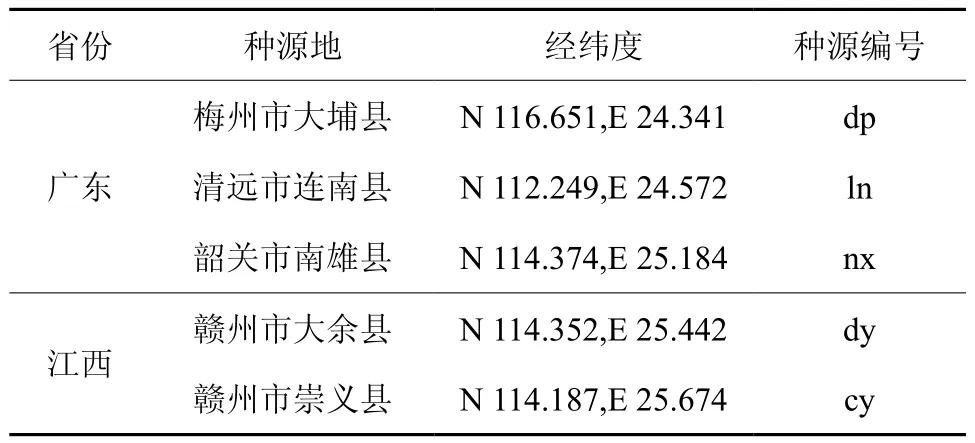
表1 种源家系材料Table 1 Material of provenances
1.2 数据收集
参照臧润国等[7]和孙玉玲等[9]的方法测定球果、种子特征数据。球果特性数据在收集球果后进行测定,其中球果长径直接测定球果纵轴上球果的长度,球果短径为球果中部横向的最大直径;两性状重复测定3次,每次重复测定5个球果数据(共测15个球果)。在球果晾晒干种子自然脱落后,测定种子特性数据,其中,种翅长度为种翅纵轴上种翅的最大长度,种翅宽度为种翅中部横轴上的长度;重复测定3次,每次重复测定5个种子数据(共测15个种子);千粒重由100粒种子的重量计算得出,重复测定3次。
1.3 统计分析方法
运用SAS 9.1开展数据分析,统计模型:yijkl=μ+pi+fj(pi)+bk+bk*fj(pi)+eijkl,其中,yijkl为在k区组内的第i种源第j家系第l个体观察值,μ为群体均值,pi为第i种源效应,fj(pi)为第i种源内的第j家系效应,bk为第k区组效应,eijkl为在k区组内的第i种源第j家系第l个体误差。
方差分析使用Proc GLM过程,多重比较采用Duncan方法;性状表型相关采用Proc Corr过程[13];性状遗传相关系数估算根据公式其中covGXY为性状X和Y的遗传协方差,为性状X和Y的遗传方差,运用Proc Mixed过程估算[16-17]。
运用主成分分析及聚类分析等多元统计方法进行因子统计分析[18],其中,主成分分析采用SAS PROC PRINCOMP过程;聚类分析运用SAS PROC CLUSTER和SAS PROC TREE过程[19]。
2 结果与分析
2.1 球果种子性状的差异分析
球果、种子性状特性数据在种源间存在很大的差异(图1)。各性状方差分析结果见表2,球果长径性状的种源效应、种源内的家系效应均达到极显著水平(P值<.0001),区组效应达到极显著水平,但种源内家系与区组的互作效应差异不显著;球果短径性状方差分析结果与球果长径性状相同,只是其种源内家系与区组的互作效应差异同样达到极显著水平。种翅长度性状的种源效应达到极显著水平(P=0.001 4)、种源内的家系效应差异显著水平(P=0.017 1),区组效应、种源内家系与区组的互作效应差异均不显著;种翅宽度性状方差分析结果与种翅长度性状相同,只是其种源内家系效应的差异也达到极显著水平。千粒重性状方差分析结果,由于没有开展多次测定,没有区组和互作效应,其种源、种源内家系效应的差异均达极显著水平(P<.0001)。这些说明球果、种子特性在种源、种源内家系的差异是真实存在的,不同种源间、不同家系间的球果和种子的差异较大。
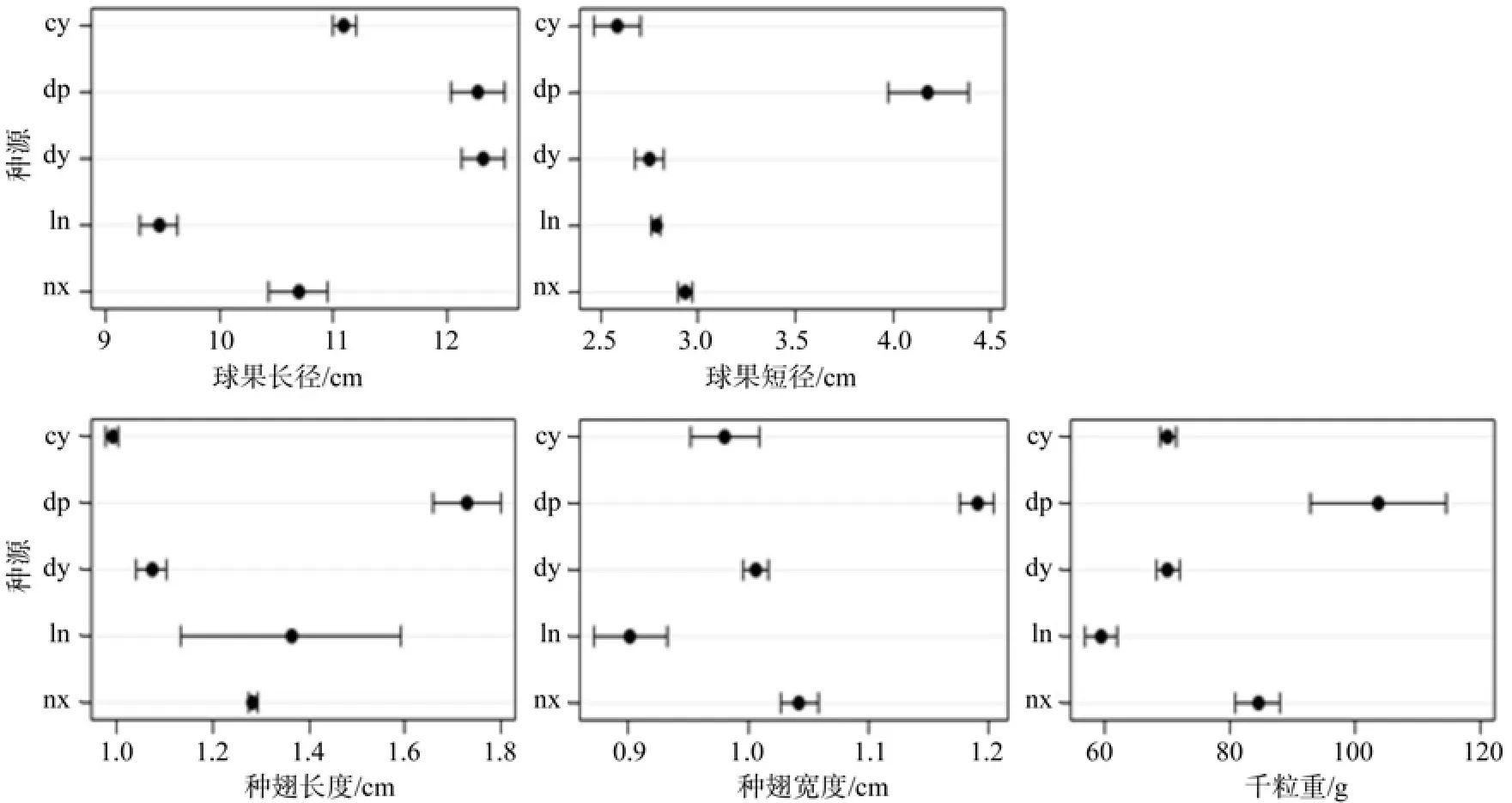
图1 江南油杉球果种子性状描述统计( 平均值±标准误)Fig. 1 Character statistics of the seeds of Keteleeria cyclolepis
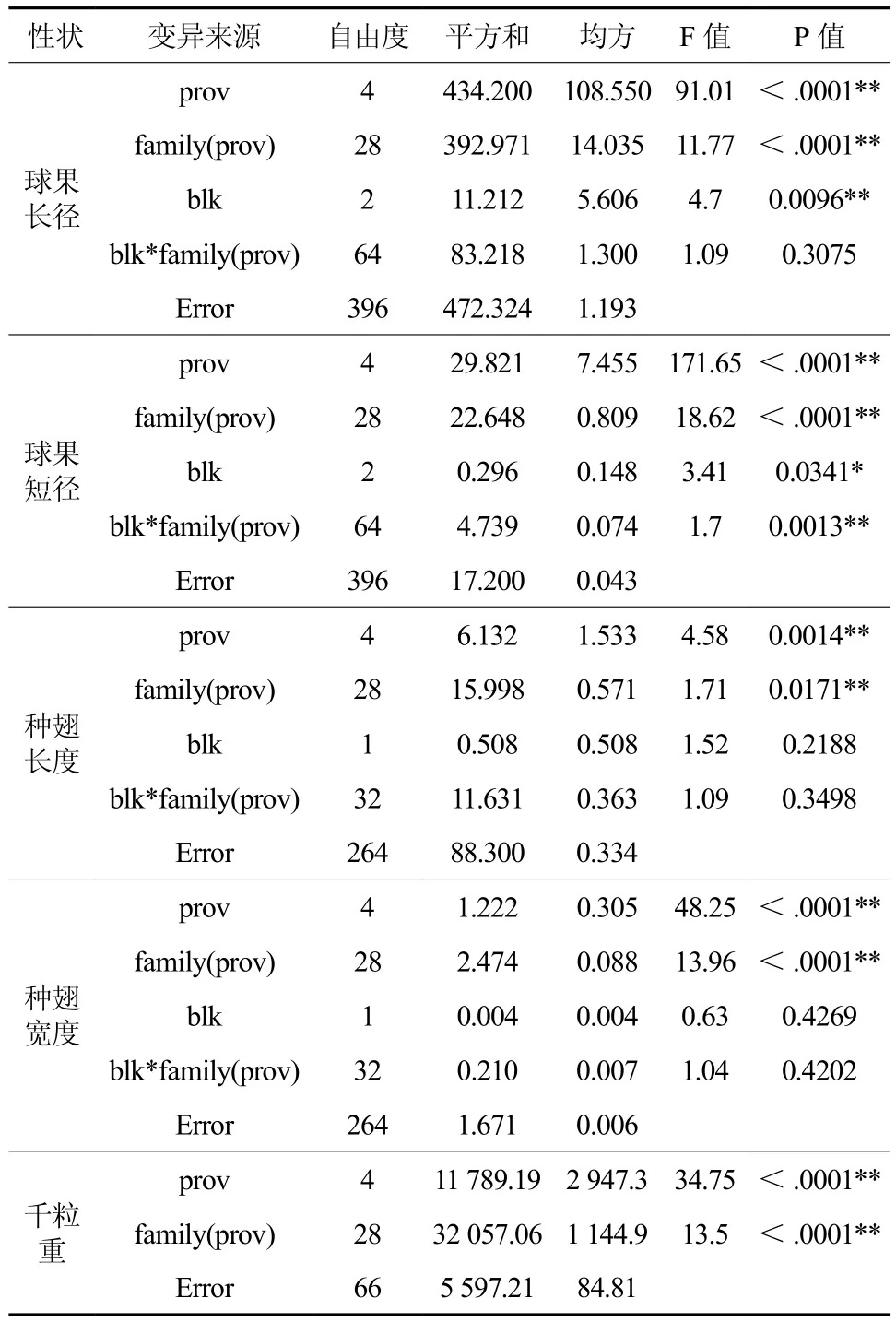
表2 方差分析结果†Table 2 Results of ANOVA
2.2 球果种子性状多重比较分析
球果、种子性状在种源间的Duncan多重比较结果如表3所示。球果长径大小顺序为:dy>dp>cy>nz>ln,球果短径大小顺序为:dp>nx>ln>dy>cy。这表明dy种源的球果短径较小、但纵长较大,为细长型球果;dp种源球果短径较大、纵长也较大,为粗大型球果;cy崇义种源球果短径最小、纵长较一般,为较细小球果;nx南雄种源球果短径最大、纵长较小,为粗短型球果;ln连南种源球果短径一般、纵长最小,也是较细小球果。
种翅长度、种翅宽度和千粒重都是dp大埔种源最大,属于大粒种子;ln连南种源的种翅长度较大,但其种翅宽度和千粒重都是最小,属于细长型种子;nx南雄种源的种翅长度一般,但种翅宽度和千粒重都较大,属于宽短型种子;dy大余种源和cy崇义种源种翅长度、种翅宽度和千粒重都较小,属于较细小种子。
2.3 相关分析
球果长径、球果短径、种翅长度、种翅宽度和千粒重等五个性状的表型、遗传相关系数如表4所示。五个性状的相关系数都为正,表明它们的存在正向的相关性,呈现球果性状越大种子性状也越大的趋势。其中,球果长径与种翅宽度间的表型相关系数为0.735,遗传相关系数为0.749,均达到显著水平,说明球果长径较大种翅宽度也将较大;球果短径与种翅长度、种翅宽度和千粒重的表型相关系数分布为0.876、0.878、0.795,遗传相关系数分别为0.944、0.890、0.861,均达到极显著水平,说明球果短径越大,种子的种翅长度、种翅宽度和千粒重将越大。
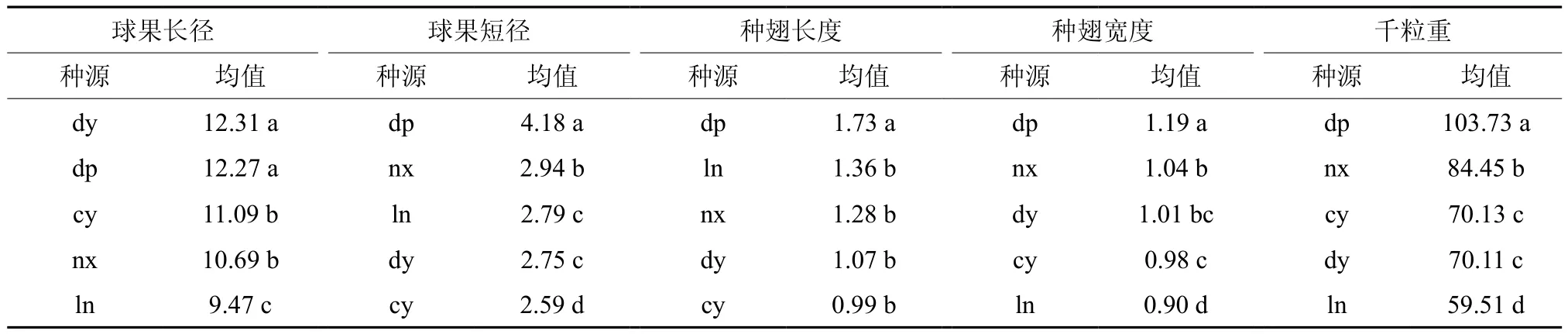
表3 Duncan多重比较结果Table 3 Results of Duncan multi-comparison
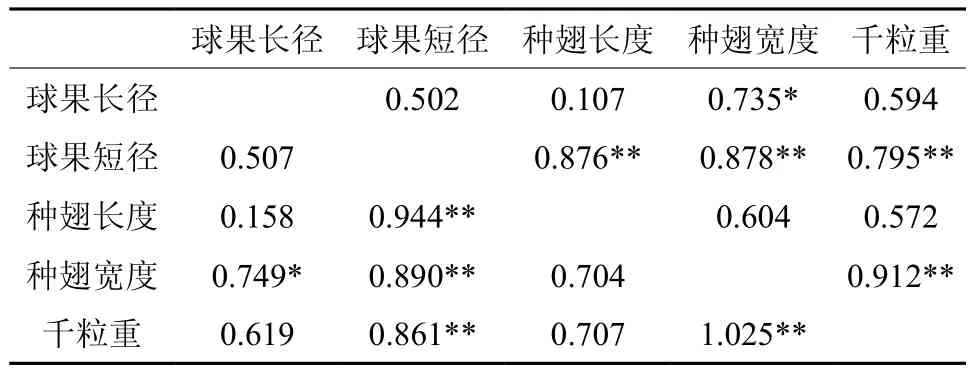
表4 性状表型及遗传相关†Table 4 Characters phenotypic and genetic correlation
2.4 主成分分析
为了综合反映球果、种子等的五个测定性状在不同种源间差异的信息,采用多元分析技术中的降维分析方法,对不同种源间的球果种子性状开展主成分分析,五个主成分信息如表5所示,第一主成分解析了总变异的77.1%,第二主成分解析了总变异的19.6%,前两个主成分累计解析了总变异的96.7%,说明用前两个主成分可较好地解析了五个测定性状反映的不同种源间差异的信息。前两个特征向量计算得出的不同种源的前两个主成分值见表6。
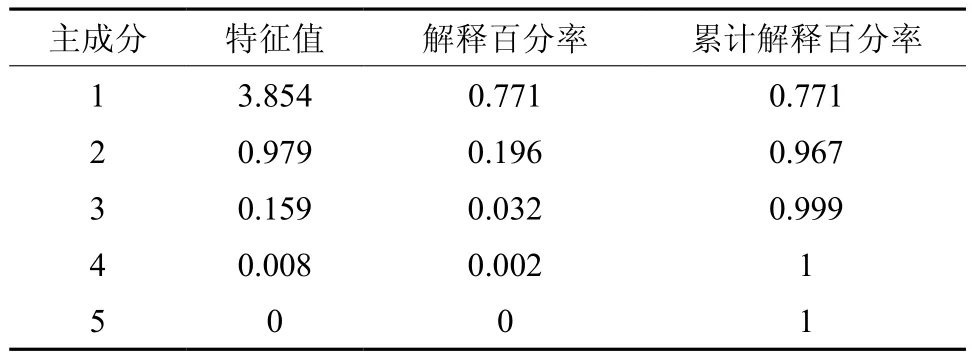
表5 主成分信息Table 5 Information of principal component analysis
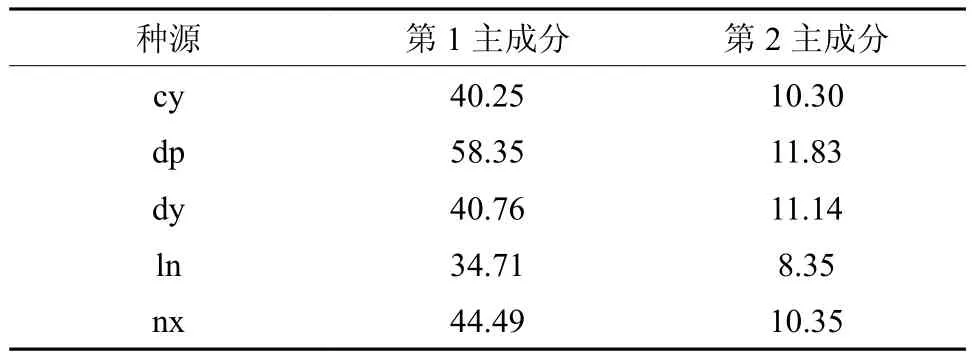
表6 种源主成分量值Table 6 Values of different PCA
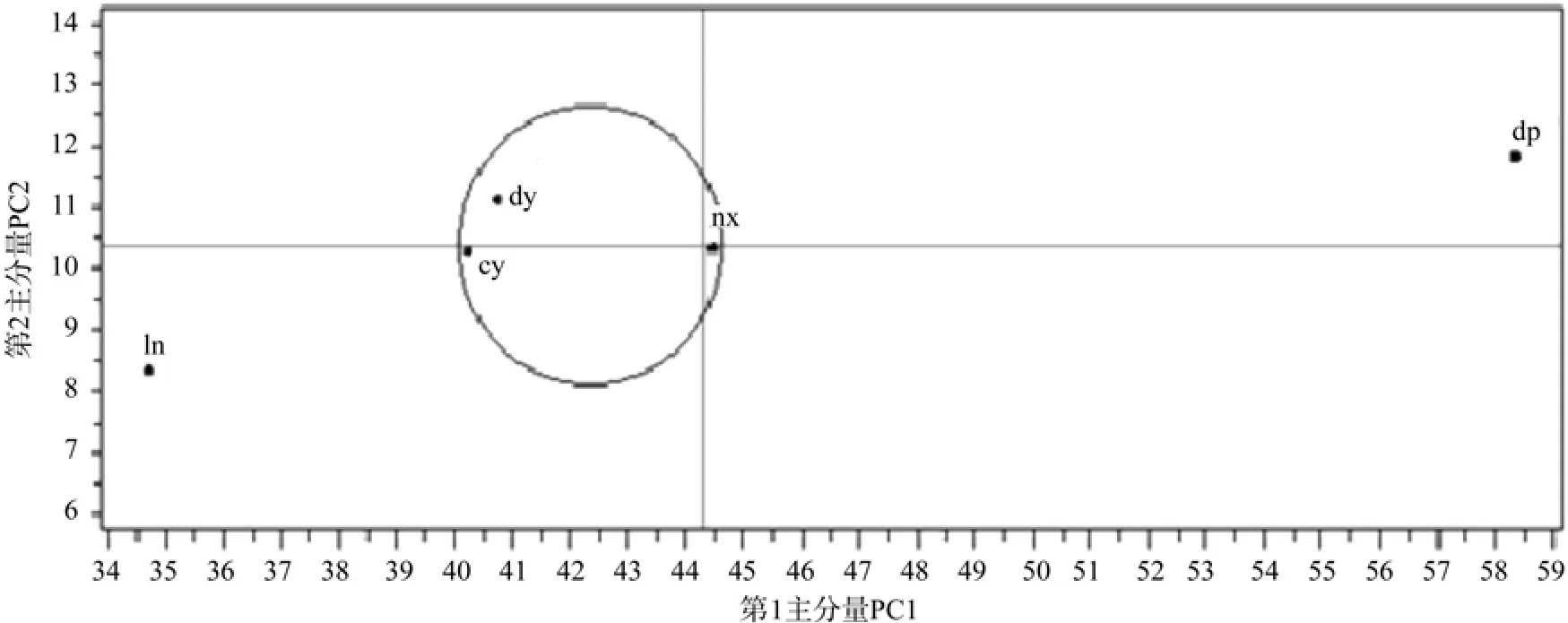
图2 种源两主成分的散点图(横、竖线为各种源主分量的均值,圆表示其中的种源相似性大)Fig. 2 Plot of two PCA of provenances (vertical and horizontal lines were the mean values, provenances in the circle were more similarity)
根据不同种源主成分值,以第一主成分为横坐标、第二主成分为纵坐标,作不同种源的散点图如下图2。从图2中,可较明显地看出不同种源的差异性,dp大埔种源的两个主成分值均较大,dy大余种源、cy崇义种源和nx南雄种源两主成分值均处于中等水平,而ln连南种源的两个主成分值均较小。根据主成分值可初步判断dy大余种源、cy崇义种源和nx南雄种源的球果种子特性较为相似,与dp大埔种源和ln连南种源有较明显的差异。
2.5 聚类分析
为了更好地反映不同种源间的球果种子等特性的差异性,各测定性状按欧几里得距离算法,球果种子等性状的差异性在SAS PROC DISTANCE中计算,再用PROC CLUSTER和PROC TREE作聚类分析如图3所示。图3更为直观表现出不同种源球果种子特性差异程度,cy崇义种源和dy大余种源首先聚为一类,说明它们间的最相似,都为江西省的种源;与广东的nx南雄种源最相似,其次是ln连南种源,最后是dp大埔种源。
五个种源的地理分别位置图如下如下图4所示,尽管nx南雄种源、dy大余种源和cy崇义种源在行政区划上隶属于不同省份,但在地理位置上是相互连接的,位于南岭山脉和武夷山脉交界处。Ln连南种源位于粤北偏西方向,处于广东与湖南和广西交界处;dp大埔种源位于粤东,处于广东与福建交界处。广东的三个江南油杉种源地在空间上存在较大距离的隔离,而江西的dy大余种源和cy崇义种源与广东的nx南雄种源的空间隔离程度较小。
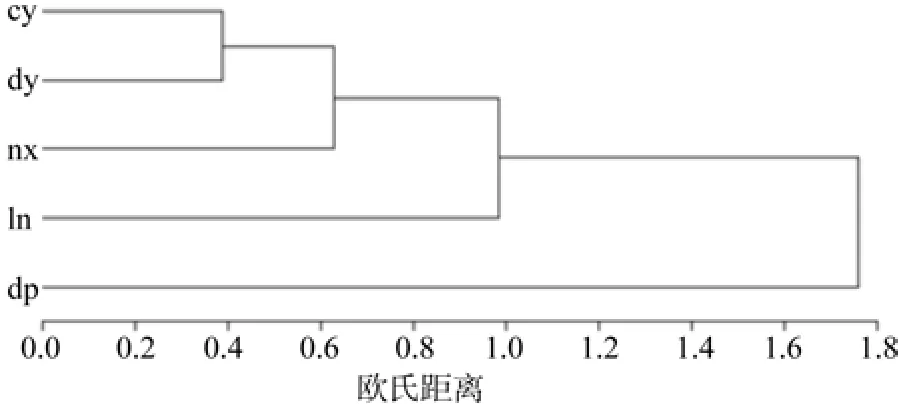
图3 种源聚类图Fig. 3 Cluster of provenances
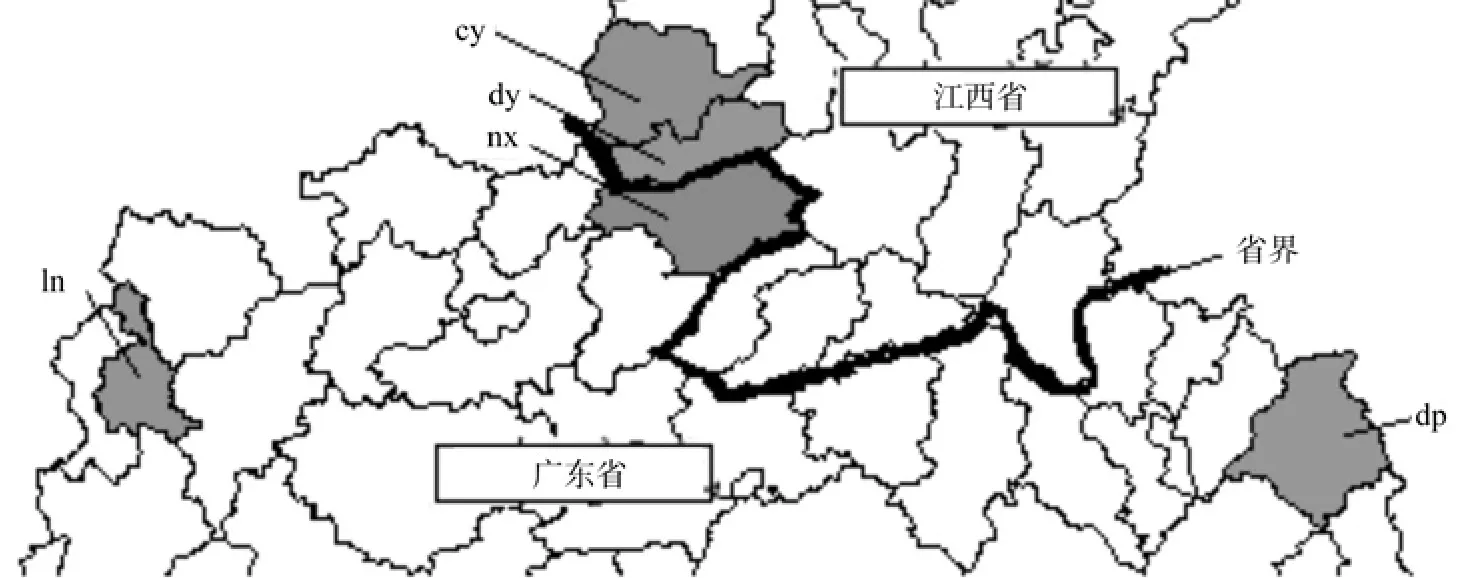
图4 种源地理位置图Fig. 4 Geographical location of different provenances
3 结论与讨论
由于天然分布区日益减少,江南油杉被列为珍贵树种加以保护利用。为更好地推广利用江南油杉种质资源,首先应对其遗传多样性开展研究。本研究从球果和种子的形态特征方面对不同种源的差异进行分析,揭示了江西的大余、崇义和广东的连南、南雄和大埔等五个种源地上的差异存在极显著差异,表明了在种源水平上江南油杉由于空间的隔离在球果和种子的形态特性方面上差别较大。种源间的多重比较分析进一步证明差别的证实存在。
多元统计分析中的主成分分析和聚类分析的结果都说明,与行政区划不同,nx南雄种源与dy大余种源和cy崇义种源的在球果和种子的特性上的相似性较大,而与ln连南种源和dp大埔种源的相似性较小。从五个种源地的地理分布位置来看,nx南雄种源、dy大余种源和cy崇义种源由于地理位置的邻近,可以推测其空间隔离而形成居群的时间相对较短,从而导致在种源水平上存在较大的相似性。这可能就是主成分分析和聚类分析中的上述三种源存在较大的相似性的原因。
另外,球果长径、球果短径、种翅长度、种翅宽度和千粒重等五个性状的表型、遗传相关分析显示球果短径与种翅长度、种翅宽度和千粒重的表型相关均达到极显著水平,说明球果短径越大,种子的种翅长度、种翅宽度和千粒重将越大。
[1]何国生. 福建江南油杉4种天然林群落物种结构特征[J]. 西南林业大学学报,2011,31(5):1-5.
[2]常翼原,张 斌,许 琪,等. 氮、磷、钾对江南油杉形态及生理变化的影响[J]. 中南林业科技大学学报,2015,35(5):46-50.
[3]翁 闲. 福建江南油杉天然种群分布规律研究[J]. 福建林业科技,2008,35(4):12-14.
[4]孙玉玲, 李庆梅, 杨敬元,等.秦岭冷杉球果与种子的形态变异[J]. 生态学报,2005, 25(1):176-181.
[4]Hornung C, Sosa V. Morphological variat ion in Puya(Bromeliaceae): an allometric study[J]. Plant Systematics and Evolution, 2006, 256: 35-53.
[5]阎爱民, 陈文新. 苜蓿、草木樨、锦鸡儿根瘤菌的表型多样性分析[J]. 生物多样性, 1999, 7(2): 1-8.
[6]顾万春.统计遗传学[M],北京: 科学出版社, 2004.
[7]臧润国,刘贵峰, 巴哈尔古丽阿尤甫, 等,天山云杉球果大小性状的地理变异规律[J]. 林业科学,2009,45(2):27-32.
[8]王小平,刘晶岚,王九龄,等,白皮松种子及球果形态特征的地理变异[J]. 北京林业大学学报[J],1998,20(3):25-31.
[9]孙玉玲,李庆梅,杨敬元,等,秦岭冷杉球果与种子的形态变异[J]. 生态学报,2005, 25(1):176-181.
[10]王旭军,张日清,许忠坤,等. 红榉不同种源种子形态性状变异[J]. 中南林业科技大学学报,2015, 35(1): 1-7.
[11]梁晓静,韦晓娟,李开祥,等. 肉桂果实、种子形态特征的研究[J]. 中南林业科技大学学报,2012, 32(9): 65-68.
[12]Gil L, Climent J, Nanos N,et al.Cone morphology variation inPinus canariensisSm[J]. Plant Systematics and Evolution, 2002,235(1): 35-51.
[13]王润辉,胡德活,郑会全,等,杉木2.5代种子园开花物候遗传变异分析[J]. 西南林业大学学报[J]. 2013,33(4):25-29.
[14]Xiang B, Li B, Isik F. Time trend of genetic parameters in growth traits ofPinus taedaL.[J]. Silvae Genetica, 2003, 52(3-4): 114-121.
[15]Isik F, Goldfarb B, LeBude A,et al. Predicted genetic gains and testing ef fi ciency from two loblolly pine clonal trails[J]. Can J For Res, 2005, 35(7):1754-1766.
[16]王润辉,胡德活. SAS Proc Mixed在林木育种试验统计中应用简介[J]. 广东林业科技,2011, 27(3):50-54.
[17]王润辉,胡德活,郑会全,等,杉木无性系生长和材性变异及多性状指数选择[J]. 林业科学, 2012,48(3):45-50.
[18]温小荣,蒋丽秀,郑 勇,等. 因子分析在马尾松人工林蓄积相关因子评价中的应用[J].中南林业科技大学学报,2014,34(11): 32-36.
[19]Cron G V, Balkwill K, Knox E B. Multivariate analysis of morphological variation in Cineraria deltoidea (Asteraceae,Senecioneae)[J]. Botanical Journal of the Linnean Societ y, 2007,154(4): 497-521.
Analysese of genetic variation of fruit and seed on Keteleeria cyclolepis
WANG Run-hui1, HU De-huo1, ZHENG Hui-quan1, YAN Shu1, JIANG Yi2, ZHU Ji-yu2
(1. Guangdong Academy of Forestry, Guangzhou 510520, Guangdong, China;2. Guangxi Academy of Forestry, Nanning 530002, Guangxi, China)
Keteleeria cyclolepisFlous is the rare timber species in China, but its distribution becomes smaller and smaller. For the sake of evaluation of the diversity ofKeteleeria cyclolepis, fi ve provenances within Guangdong and Jiangxi provinces were applied to study the phenotypical variation of cone and seed. Results from ANOVA indicated that phenotypical differeces of both cone and seed reached highly signi fi cant level. Multiple comparison also approved that. Correlation between all the fi ve traits showed that cone diameter had signi fi cant correlation with seed wing length, seed wing width and Thousand-grain weight. The similarity between the fi ve provenances was revealed by principal components analysis and cluster analysis. Unlike administrative zoning, provenance of nanxiong (nx) from Guangdong province and provenances of Dayu (dy) and chongyi (cy) from Jiangxi province possessed more similarity, perhaps because of the smaller geographical isolation.
cones; seeds; genetic variation;Keteleeria cyclolepis
S791.15
A
1673-923X(2016)02-0019-06
10.14067/j.cnki.1673-923x.2016.02.004
http: //qks.csuft.edu.cn
2014-10-29
国家林业局林业行业公益性专项“江南油杉良种选育及栽培关键技术研究”(201304108)
王润辉,高级工程师
胡德活,研究员;E-mail:hudehuo@163.com
王润辉,胡德活,郑会全,等. 江南油杉球果种子特性种源变异分析[J].中南林业科技大学学报,2016, 36(2): 19-24.
[本文编校:吴 彬]
猜你喜欢
特产研究(2022年6期)2023-01-17
绿色科技(2021年21期)2021-11-26
源流(2021年1期)2021-07-28
辽宁林业科技(2021年1期)2021-03-16
农家科技中旬版(2020年4期)2020-07-06
岭南音乐(2019年6期)2019-12-31
艺术家(2019年4期)2019-04-20
中国森林病虫(2019年1期)2019-03-04
中国林副特产(2019年3期)2019-01-10
神州民俗(2018年9期)2018-11-21
