
马尾松生物量影响因素的通径分析
2016-12-19 23:06黄志宏陈振雄
中南林业科技大学学报 2016年8期
朱 佳 ,黄志宏 ,陈振雄 ,孙 华
(1. 中南林业科技大学 a.生命科学与技术学院; b.南方林业生态应用技术国家工程实验室;c.林业遥感信息工程研究中心,湖南 长沙 410004;2. 国家林业局 中南林业调查规划设计院, 湖南 长沙 410014)
马尾松生物量影响因素的通径分析
朱 佳1a,1b,黄志宏1a,1b,陈振雄2,孙 华1c
(1. 中南林业科技大学 a.生命科学与技术学院; b.南方林业生态应用技术国家工程实验室;c.林业遥感信息工程研究中心,湖南 长沙 410004;2. 国家林业局 中南林业调查规划设计院, 湖南 长沙 410014)
林木生物量受多重因素的综合影响,同一树种在不同区域内水热因子的变化也可能会影响树木各器官生物量的分配关系。选择南方150棵马尾松为研究对象,采集样木所在的空间位置(经度、纬度)、年均温度、年均降水、胸径、树高等相关因子,测定树干、树皮、树枝、树叶、树根各器官生物量。运用方差分析、通径分析等方法,分析北亚热带、中亚热带、南亚热带内马尾松各器官生物量与影响因素的关系,利用变异系数探讨了不同气候带马尾松生物量对水热变化敏感程度的器官差异和区域差异。研究结果表明:(1)马尾松各器官生物量对温度升高的敏感程度表现为根最高,叶最低,按大小顺序为根>枝>皮>干>叶;各器官生物量对降水增加的敏感程度表现为皮最高,干最低,按大小顺序为皮>枝>叶>根>干。(2)马尾松生物量对温度和降水变化的敏感程度的区域差异明显,空间上呈现出北亚热带>中亚热带>南亚热带,从北往南敏感程度逐渐减小。(3)胸径和树高因子均与各器官生物量存在显著相关关系(P< 0.05)。胸径对各器官生物量的直接通径系数接近于1(P < 0.001),树高通过胸径影响生物量的间接通径系数接近于1。
生物量;通径分析;变异系数;马尾松
森林生物量是衡量森林生产力的重要标志,也是评估研究森林生态系统固碳的基础数据[1-2]。近年来森林生物量的估算成为林业科研领域的热点问题[3]。森林生物量的测定方法主要包括3大类:生物量实测方法、模型估算方法以及遥感反演方法[4-5]。生物量实测方法破坏性强,目前已经很少采用。基于遥感的方法是区域及全球范围碳储量估计的常用手段,通过分析遥感影像各波段的光谱反射率,植被指数及衍生因子与地面实测值之间的关系,建立相应的反演模型来实现大范围内的碳储量的模拟,同时也要意识到光学传感器数据对于高密度林分生物量的饱和问题[6]。基于模型的估算方法仍旧是区域范围森林生物量监测与评估的重要手段。建立通用生物量模型是进行区域或全国范围生物量估测的基础[7],全世界已经建立的生物量(包括总量和各分量)模型超过2 300个,概括起来包括线性和非线性两种形式,由于生物量建模对象的不同,方程的结构形式存在差异[8]。
林木生物量大小及其各器官分配格局除了与林分密度、林分起源、胸径、树高等生物因素相关之外,还受到环境因素(气候、地形、土壤条件)的影响[9-11]。不同地域生态环境的差异,同一树种不同起源、不同区域内温度或降水的变化会改变树木的生长策略,进而影响树木各器官生物量的分配。当前,林分生物量的地理分布特征及与气候的关系研究较多[12-15],但温度、降水等因素与单个树种各器官生物量之间关系的定量研究较少[16-18]。本研究以南方马尾松为研究对象,收集了林木位置(经度和纬度)、年均温度、年均降水、胸径、树高因子,及树木的干、皮、枝、叶、根的各器官生物量指标。运用方差分析、通径分析等方法,分析不同气候带内马尾松各器官生物量与影响因素的关系,利用变异系数探讨了不同气候带马尾松生物量对水热变化敏感程度的器官差异和区域差异,为揭示区域尺度下马尾松生物量对水热的响应和反馈机制提供参考依据。
1 材料与方法
1.1 数据来源
研究所涉及的150棵马尾松立木生物量数据来自2009年国家森林资源连续清查生物量调查建模项目。样木采集区域包括江苏、浙江、安徽、福建、江西、湖南、广东、广西、贵州等9省区。样木数量按照各省的森林资源比重进行分配,全部样木按2、4、6、8、12、16、20、26、32、38 cm 以上共10个径阶均匀分布,每个径阶的样木按树高级均匀分布,样木在大尺度范围具有广泛的代表性[8]。
所有样木胸径、树高等测树因子在野外调查时伐倒后进行测量,记录每木的位置,干材、干皮、树枝、树叶、树根的鲜重(只采集了54株树的全部根系)。对野外采集的150棵样木,按照技术规程要求抽取样品带回实验室烘干,根据烘干后样品的质量推算各器官生物量和样木总生物量。
通过样木位置提取其所在的气候带(见图1),所有样木都落在北亚热带、中亚热带和南亚热带3个气候带内。温度和降水数据从中国气象科学数据共享服务网下载(http://cdc.nmic.cn/home.do),运用地理信息系统软件生成年度降水和年均温度图,再以150棵马尾松最大的年龄为基准,计算出整个生长期内的年均温度(MAT)和年均降水(MAP),以作为分析马尾松生物量的气候影响因子。
1.2 数据分析
位置信息、年均温度和年均降水的提取。利用GIS软件提取每株样木对应的经纬度坐标信息。然后根据林木年龄,获取整个生长期内年度降水和气温数据,在GIS软件中进行均值计算,得到每株样木对应年龄的年均降水和年均温度。
采用单因素方差分析法(One-way ANOVA)对3个气候带内样木位置(经度、纬度)、年均温度、年均降水、胸径、树高因子、马尾松各器官生物量等进行差异显著性检验。显著性水平为0.05。
运用通径分析(Path analysis)研究各器官生物量与影响因素的因果关系[19]。基于温度和降水对生物量的直接和间接作用,运用变异系数(公式1,公式2)比较分析马尾松对水热变化响应的器官变异特征和气候带间区域差异。数据分析和图形制作均在R软件[20]中完成。
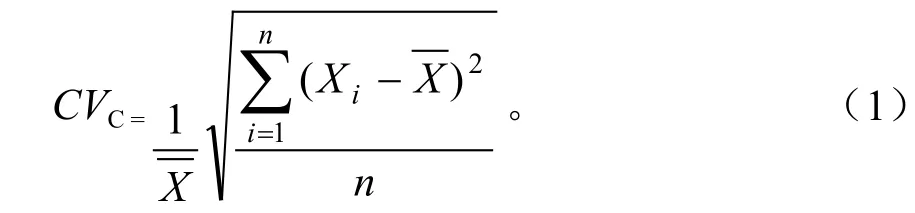
式(1)中,CVC为气候带内变异系数,n为气候带个数,Xi为第i个气候带的直接或间接通径系数,为气候带直接或间接通径系数的均值。
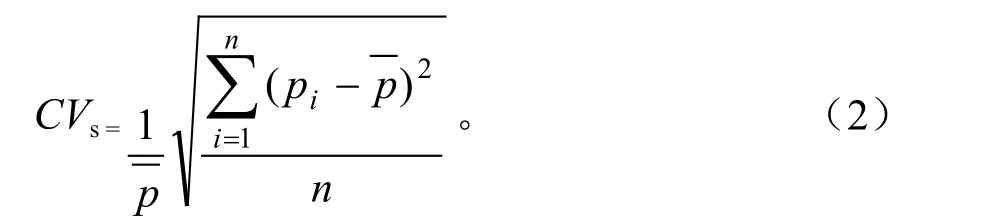

图1 马尾松样木分布Fig. 1 Location of Pinus massoniana distribution
式(2)中,CVs为气候带间变异系数,n为器官的种类个数,Pi为i第种器官的直接或间接通径系数,为器官直接或间接通径系数的均值。
2 结果与分析
2.1 生物量及其影响因素的方差分析
单因素方差分析是对影响因素在不同气候带中的差异显著性水平的总体判断。通过多重比较能进一步比较影响因素两两之间的差异情况。单因素方差分析结果见表1。
由表1可知,除经度的差异不明显外,纬度、年均温度和年均降水的F值明显大于其他因素,树高、胸径的P值也是极显著和很显著。方差分析表明,3种不同气候带下纬度、年均温度、年均降水差异极显著,胸径、树高及树干、皮、枝生物量等指标的差异也显著,说明不同气候带的生境塑造了马尾松生态特性的差异。
生物量影响因素及各器官生物量的多重比较结果如表2所示,年均温度和年均降水在3种气候带水平下两两之间差异极显著,说明温度和降水作为主要气候因子,其水平地带性差异显著,能有效的解释各器官生物量对水热变化响应的敏感程度的区域差异。胸径、树高在中亚热带与北亚热带,中亚热带与南亚热带均有显著差异,而在北亚热带与南亚热带没有显著差异。3种气候带的枝、叶、根生物量均值无明显差异,树干、树皮有一定差异。生物量影响因素多重比较结果表明不同气候带对干材生物量影响的差异,中亚热带地区较其他气候带更加显著,这与胸径、树高
的情况相一致。而叶、根生物量的多重比较结果与胸径、树高的差异结果并不一致。这一现象说明中亚热带地区的胸径、树高与树干、皮、枝生物量具有很强的相关性,而叶、根生物量可能更易受地带性规律的影响。
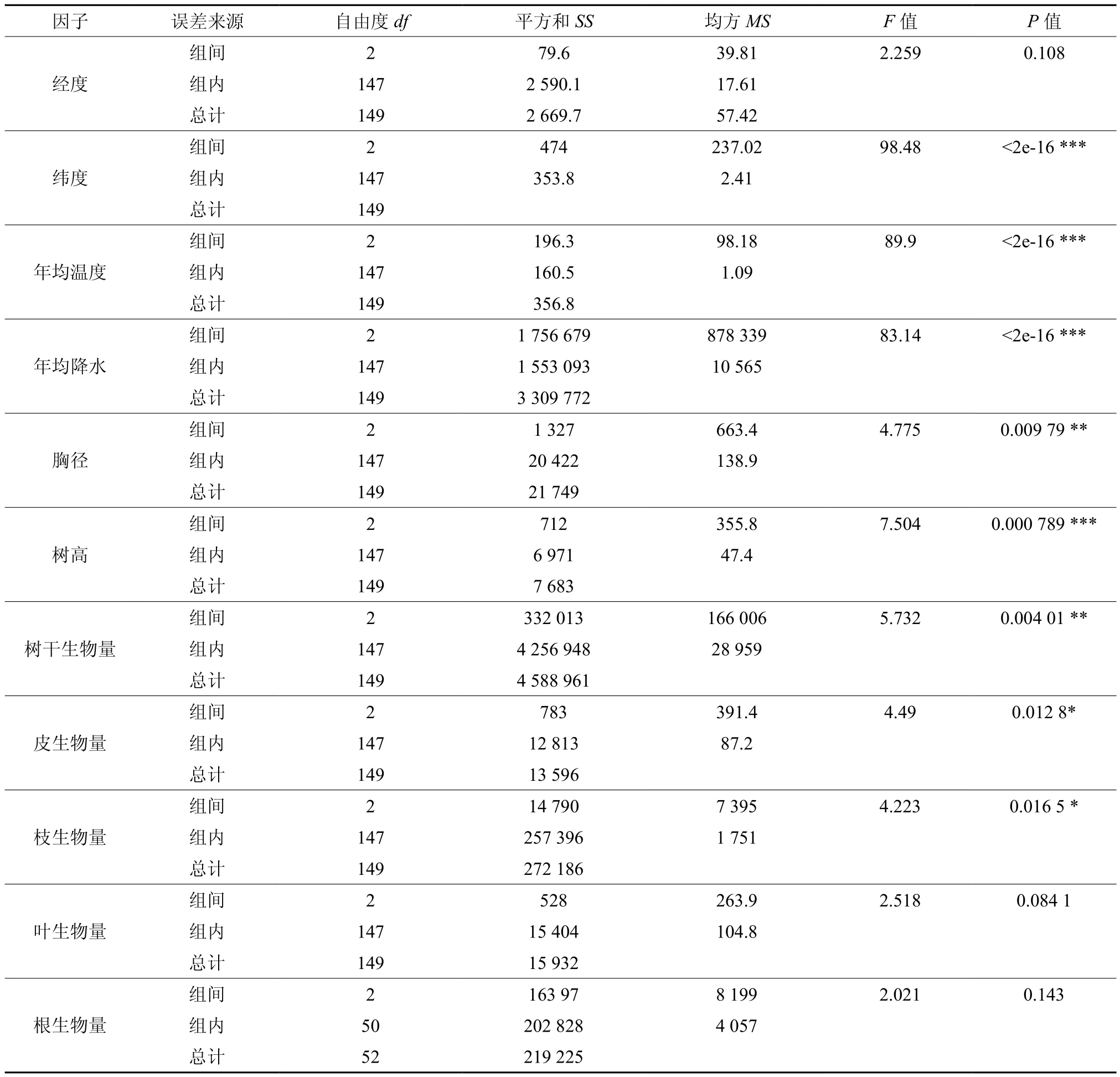
表1 马尾松生物量及影响因素的方差分析†Table 1 ANOVA of organs biomass and influencing factors for Pinus massoniana

表2 各气候带的生物量影响因素及各器官生物量的比较(平均值±标准差)†Table 2 Comparison of organs biomass and influencing factors for three kinds of climate zone
2.2 生物量影响因素的通径分析
通径分析是标准化的多元线性回归分析,将相关系数拆分为自变量对因变量的直接作用,和通过其他变量对因变量的间接作用,此多元分析方法有利于更准确的解释多个自变量与因变量之间的本质关系[21-22]。经度和纬度对马尾松生物量的直接作用如图2所示,北亚热带明显高于中亚热带和南亚热带;北亚热带的经度和纬度对干生物量的直接作用最小,对根生物量的直接作用方向也与其他器官有明显差异;北亚的纬度对干、枝、叶生物量的直接作用为负。

图2 影响因素的直接作用Fig. 2 Direct effects of influencing factors
年均温度和年均降水对各器官生物量的直接作用是北亚热带最高,中亚和南亚热带明显降低且差别不大。年均温度的直接作用在中亚热带出现一个低值(根除外)。图2中,北亚热带的年均温度和年均降水对干、枝、根的直接和间接作用的绝对值均较大,方向相反。北亚热带年均温度的直接作用绝对值大小的顺序为根>枝>干>叶>皮,年均降水的直接作用绝对值大小的顺序为根>枝>叶>干>皮。北亚热带年均温度通过纬度对枝的间接作用(2.95)超过了其直接作用(0.9);通过经度、纬度对叶的间接作用(-2.16,2.31)超过了其直接作用(0.38);通过经度、纬度对根的间接作用(3.89,-3.16)超过了其直接作用(-2.05)。北亚热带地区年均温度对枝、叶、根生物量通过经度或纬度的间接作用超过其直接作用,说明该地区年均温度与地理位置共同作用影响了枝、叶、根的生物量。另外,北亚热带年均温度对枝、根生物量的直接作用,及通过经度、纬度的间接作用均明显大于干、叶。年均降水由北往南,对干生物量的直接作用大小逐渐降低。年均降水对根和枝的直接作用,及通过纬度对根和枝的间接作用均明显大于其他器官。
表3和表4是胸径和树高对各器官生物量的直接作用和通过其他因素的间接作用。胸径与各器官生物量的直接作用均很大,通过其他因素的间接作用很小(见表3)。树高与各器官生物量的直接作用很小,通过胸径的间接作用却很大(见表4),表明间接作用改变了树高与生物量的相关系数。
2.3 敏感因素的差异分析
基于温度和降水对马尾松各器官生物量的通径系数,采用变异系数获取通径系数在不同气候带的波动程度。表5和表6列出了年均温度和年均降水的直接、间接作用及其在北、中、南亚热带的气候带间和带内变异系数。气候带间变异系数说明各器官生物量对年均温度和年均降水变化的敏感程度的器官差异。各器官生物量对温度变化的敏感程度的器官差异按大小顺序是根>枝>皮>干>叶,对降水变化的敏感程度的器官差异按大小顺序是皮>枝>叶>根>干。气候带内变异系数说明各器官生物量对年均温度和年均降水变化的敏感程度的区域差异,差异程度的大小顺序大致为北亚热带>中亚热带>南亚热带。
变异系数反映出各器官生物量对年均温度和年均降水变化的敏感程度的差异。亚热带地区马尾松器官对水热条件变化的响应,根对温度变化的敏感程度最大,叶最小;皮对降水变化的敏感程度最大,干最小(图3a)。各器官生物量对温度变化响应的区域变异程度是从北亚热带往南亚热带逐渐减小,地带性特征受温度的影响是逐渐减小;对降水变化的区域变异程度也大致是从北亚热带往南亚热带逐渐减小,只是间接作用的变异系数的大小顺序为南亚热带>北亚热带>中亚热带(图3b)。

表3 胸径对各器官生物量的影响Table 3 Effects of DBH on organs biomass

表4 树高对各器官生物量的影响Table 4 Effects of tree height on organs biomass
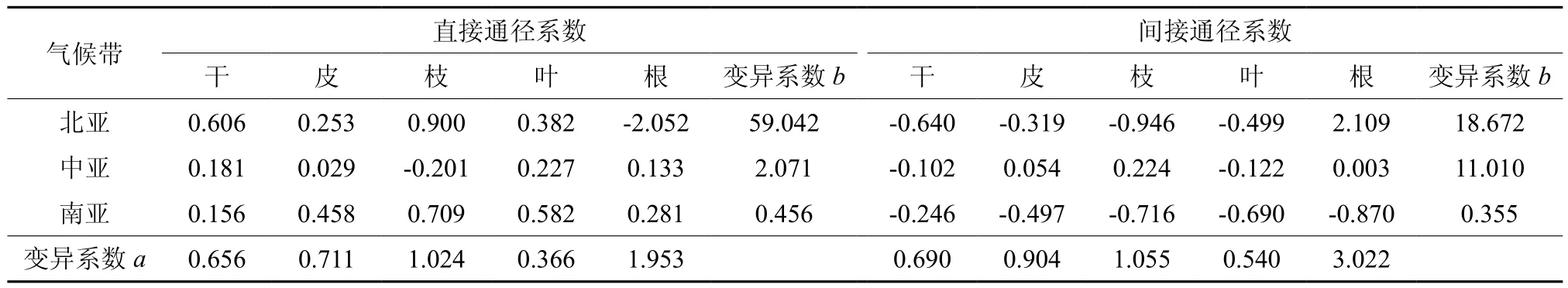
表5 变异系数根据年均温度的直接和间接作用的比较†Table 5 Comparison of coefficient of variation on direct effects and indirect effects of MAT
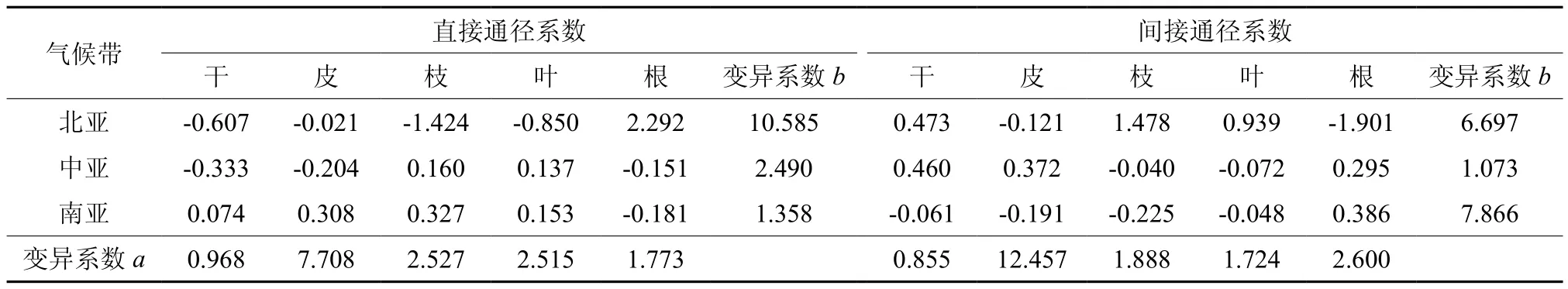
表6 变异系数根据年均降水的直接和间接作用的比较†Table 6 Comparison of coefficient of variation based on direct effects and indirect effects of MAP

图3 年均温度和年均降水的变异系数Fig. 3 Coefficient of variation of MAT and MAP
3 结论与讨论
论文以南方马尾松生物量数据为基础,提取林木位置、年均温度和降水等环境因子,采用方差分析方法分析不同气候带内马尾松生物量与其影响因素之间的规律;运用通径分析方法开展影响因素与各器官生物量的直接作用;结合变异系数,计算不同气候带内马尾松对水热变化响应的器官变异特征和气候带间区域差异。主要研究结果如下:
(1)温度和降水对马尾松生物量的影响存在地带性差异。从北亚热带、中亚热带和南亚热带3个气候带的生物量影响因素的方差分析结果来看,经度均值差异不显著,纬度、年均温度和年均降水的均值差异都很显著,且温度和降水因子呈现出从高纬向低纬递增的纬度地带性规律。另外,温度和降水对马尾松生物量的影响强度在3个气候带上存在差异,北亚热带明显高于中亚热带和南亚热带,而中亚热带和南亚热带差异不显著,这说明这3种气候带中,随着纬度的变化,影响马尾松生长的限制因子也存在差异,差异形成的原因,可能与水热因子通过与其他生态因子的间接作用有关,如通过太阳辐射、海拔、土壤等形成了影响植物生长的微环境、小气候。研究结果与程瑞梅[23]提出北亚热带的温度和降水是马尾松生长的限制因子结论一致。
(2)马尾松各器官生物量对温度和降水变化的敏感程度存在器官差异和区域差异。年均温度对各器官生物量影响的变异系数的差异表明,根对温度变化的敏感程度最大,而叶的敏感程度最小。树皮对降水变化的响应的敏感程度最大,而树干的敏感程度最小。各器官生物量对温度变化响应的区域变异程度是从北亚往南亚逐渐减小,地带性特征受温度的影响是逐渐减小;各器官生物量对降水变化的区域变异程度也大致是从北往南逐渐减小,只是间接作用的变异系数的大小顺序为南亚>北亚>中亚。比较年均降水与年均温度直接作用和间接作用的气候带内变异系数,温度的变异系数更大,说明温度比降水对器官生物量的影响在3种气候带之间波动更大,也从一定程度说明了马尾松各器官生长对温度变化的敏感程度更大。
(3)胸径是马尾松生物量模型的决定因素,树高是间接因素。研究结果一方面表明胸径作为影响生物量的决定因素,是生物量建模的关键因子。另一方面,通径分析结果显示树高主要是通过胸径对生物量产生了很强的间接作用。
总之,本研究通过各器官生物量与水热变化的关系的通径分析和变异系数的结果,讨论了马尾松与影响因素的关系,以及马尾松对水热变化的响应情况。森林植被的分布受多种因素的影响,除了年均温度、年均降水外,还与土壤、相对湿度等因素有关。这些因素与森林生物量的关系还有待进一步研究。
[1] Heather K, Brendan M, Sandra B, et al. Estimating carbon carrying capacity in natural forest ecosystems across heterogeneous landscapes: addressing sources of error [J].Global Change Biology, 2010, 16(11): 2971-2989.
[2] 郭 屹, 项文化, 刘 聪, 等. 湖南省马尾松林生物量动态特征及其对龄组结构变化的响应 [J] . 中南林业科技大学学报,2015, 35(7): 81-87.
[3] 闫 晶, 罗云建, 郑德福, 等.桉树生物量估算差异的源解析 [J] . 林业科学, 2014, 50(2): 92-98.
[4] Lu D. The Potential and Challenge of Remote Sensing-based Biomass Estimation [J]. International Journal of Remote Sensing,2006, 27(7): 1297-1328.
[5] Lu D, Chen Q, Wang G, et al. A survey of remote sensing-based aboveground biomass estimation methods in forest ecosystems [J].International Journal of Digital Earth, 2014(13): 1-43.
[6] Foody G M, Cutler M E, McMorrow J, et al. Mapping the biomass of Bornean tropical rain forest from remotely sensed data [J] . Global Ecology & Biogeography, 2001, 10(4):379-387.
[7] 王轶夫, 孙玉军. 马尾松生物量模型的对比研究 [J] . 中南林业科技大学学报, 2012, 32(10): 29-33.
[8] 曾伟生, 肖前辉, 胡 觉, 等. 中国南方马尾松立木生物量模型研建 [J] . 中南林业科技大学学报, 2010, 30(5): 50-56.
[9] Malhi Y, Wood D, Baker T, et al. The regional variation of aboveground live biomass in old-growth Amazonian forests [J].Global Change Biology, 2006, 12(7): 1107-1138.
[10] 符利勇, 曾伟生, 唐守正. 利用混合模型分析地域对国内马尾松生物量的影响 [J] . 生态学报, 2011, 31(19): 5797-5808.
[11] 李海奎, 宁金魁.基于树木起源、立地分级和龄组的单木生物量模型 [J] . 生态学报, 2012, 32(3): 740-757.
[12] 刘世荣, 徐德应, 王 兵. 气候变化对中国森林生产力的影响Ⅰ.中国森林现实生产力的特征及地理分布格局[J] . 林业科学研究, 1993, 6(6): 633-642.
[13] 秦建华, 姜志林. 杉木林生物量及其分配的变化规律 [J] . 生态学杂志, 1996, 15(1): 1-7.
[14] 王俊鸿. 不同产区杉木人工林生物生产力及其与环境因子的关系研究 [D] . 福州:福建农林大学, 2014.
[15] 刘迎春, 于贵瑞, 王秋凤, 等. 基于成熟林生物量整合分析中国森林碳容量和固碳潜力 [J] . 中国科学: 生命科学, 2015,45(2): 210-222.
[16] 彭少麟, 李跃林, 余 华, 等. 鼎湖山森林群落不同演替阶段优势种叶生态解剖特征研究 [J] . 热带亚热带植物学报, 2002,10(1): 1-8.
[17] 杜 虎, 曾馥平, 王克林, 等. 中国南方3种主要人工林生物量和生产力的动态变化 [J] . 生态学报, 2014, 34(10): 2712-2724.
[18] Reich P B, Luo Y, Bradford J B, et al. Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots[J]. Proceedings of the National Academy of Sciences, 2014,111(38): 13721-13726.
[19] Hata K, Kohri M, Morita S, et al. Complex Interrelationships Among Aboveground Biomass, Soil Chemical Properties,and Events Caused by Feral Goats and Their Eradication in a Grassland Ecosystem of an Island [J]. Ecosystems, 2014, 17(6):1082-1094.
[20] R Core Team. R: A language and environment for statistical computing[M]. R Foundation for Statistical Computing, Vienna,Austria.2013. ISBN 3-900051-07-0, URL http://www.R-project.org/.
[21] Wootton J T. Predicting direct and indirect effects: an integrated approach using experiments and path analysis [J]. Ecology, 1994,75(1): 151-165.
[22] Duffy J E, Reynolds P L, Boström C, et al. Biodiversity mediates top-down control in eelgrass ecosystems: a global comparative experimental approach [J]. Ecology Letters, 2015 (8): 696-705.
[23] 程瑞梅, 封晓辉, 肖文发, 等. 北亚热带马尾松净生产力对气候变化的响应 [J] . 生态学报, 2011, 31(8): 2086-2095.
Biomass and factors of Pinus massoniana based on path analysis
ZHU Jia1a,1b, HUANG Zhi-hong1a,1b, CHEN Zhen-xiong2, SUN Hua1c
(1a. College of Life Science and Technology; 1b. National Engineering Laboratory for Applied Technology of Forestry & Ecology in South China; 1c. Research Center of Forestry Remote Sensing and Information Engineering, Central South University of Forestry and Technology, Changsha 410004, Hunan, China; 2. Central South Forest Inventory and Planning Institute of State Forestry Administration,Changsha 410014, Hunan, China)
Forest biomass is influenced by multiple factors. The growth of trees shift in strategy altered with change of temperature and precipitation even for the same tree species in different regions, which would remodel the biomass allocation patterns. 150 pieces of masson pine(Pinus massoniana)grown in south China were studied. And the related information of the sample trees were recorded including spatial location(longitude and latitude), mean annual temperature ( MAT ), mean annual precipitation ( MAP ), tree diameter at breast height(DBH)and tree height , tree barks, stems, branches, foliage and roots. The one-way ANOVA and path analysis methods were employed to investigate the relationship between organs biomass with the influencing factors. Coefficient of variation based on the results of path analysis was used to test biomass sensitive degree to change in temperature and precipitation of organs difference and regional difference. The results showed that the sensitivity of biomass to the increasing MAT varied with organs which was in a sequence as roots > branches > bark > stems > foliage. While for the sensitivity to the increasing MAP, it was arranged as barks > branches >foliage > roots > stems. The sensitivity of masson pine biomass to changes in MAT and MAP also was significant difference in regions which presented a pattern of northern subtropics > mid-subtropics > southern subtropics with a decreasing difference from north to south. Organs biomass is significantly correlated with DBH and tree height (P < 0.05). P. massoniana biomass was found to have significant correlations with DBH (P < 0.001), which had the greatest direct influence coefficients with values close to 1. Whereas, tree height that exerted influence on biomass through DBH had the greatest indirect influence coefficients close to 1.
biomass; path analysis; coefficient of variation; Pinus massoniana
S718.55
A
1673-923X(2016)08-0088-08
10.14067/j.cnki.1673-923x.2016.08.016
2015-11-09
湖南省自然科学基金创新研究群体(湘基金委字[2013]7号);湖南省创新平台与人才计划(科技人才)项目(2015RS4048);中国博士后科学基金项目(2014M562147)
朱 佳,硕士研究生
黄志宏,副教授,博士;E-mail:huanghugh2013@yahoo.com
朱 佳, 黄志宏, 陈振雄, 等. 马尾松生物量影响因素的通径分析[J]. 中南林业科技大学学报, 2016, 36(8): 88-95.
[本文编校:文凤鸣]
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
黑龙江气象(2021年2期)2021-11-05
农家科技中旬版(2020年10期)2020-07-12
农家科技中旬版(2020年2期)2020-03-18
数学学习与研究(2019年20期)2019-11-30
绿色科技(2019年5期)2019-11-29
亚热带农业研究(2019年2期)2019-09-05
课程教育研究(2018年4期)2018-03-16
星星·诗歌原创(2017年5期)2017-08-13
环境与发展(2017年4期)2017-07-19
