
Ophiognomonia castaneae协同板栗褐缘叶枯病病原菌致病作用的研究
2016-12-17 21:59马洪兵刘国兴龚赛张秀停王庆华姜
山东农业科学 2016年11期
马洪兵+刘国兴+龚赛+张秀停+王庆华+姜淑霞


摘要:为明确板栗新病害褐缘叶枯病病原菌Phomopsis castaneae-mollissimae(Y1)与其共生菌板栗蛇孢日规壳Ophiognomonia castaneae(Y6)的关系,采用在发病不同时期组织分离、定期采样观察、致病性测定及对峙培养等方法进行研究。结果表明,发病初期在褐缘叶枯病病健交界处只分离到Y1,随病斑扩展Y6的分离频率逐渐增加,发病后期Y6的分离频率略高于Y1;Y6的无性型和有性型子实体均生长在病斑上;离体叶片接种Y1易产生病斑,接种Y6很少产生病斑,两菌混合接种比单独接种产生的病斑症状明显且发病速度快,表现出Y1致病力强,Y6致病力弱,Y6具有协同Y1致病的作用;Y6和Y1的营养体表现出明显的完全的亲和力。
关键词:Ophiognomonia castaneae;板栗褐缘叶枯病;协同致病
中图分类号:S432.4+4 文献标识号:A 文章编号:1001-4942(2016)11-0109-05
Abstract The methods such as tissue isolation in different stages, regular sampling observation, pathogenicity identification and confronting incubation were adopted to find out the relation between Phomopsis castaneae-mollissimae(Y1), the pathogeny of a new chestnut disease brown margin leaf blight,and Ophiognomonia castaneae (Y6) , a symbiotic bacteria of Y1. The results showed that at the early stage of brown margin leaf blight, only Y1 was isolated at the junction of diseased and healthy tissue. Along with the extension of lesion spots, the isolation frequency of Y6 increased gradually. At the late stage, the isolation frequency of Y6 exceeded that of Y1 slightly. The anamorph and teleomorph fruit bodies of Y6 both occurred in the lesion spots. Inoculating the 2 fungi to detached leaves respectively, Y1 produced lesion spots easily and Y6 rarely. However, the lesion spot symptom occurred more obviously and the disease developing speed was faster when the detached leaves were inoculated by Y6 combined with Y1 than that inoculated by Y1 or Y6 solo. It showed that Y1 owned the powerful pathogenicity, while Y6 owned the weak pathogenicity, and Y6 and Y1 had the coordinated pathogenic effect. The vegetative body of Y6 and Y1 had the obvious and complete compatibility.
Keywords Ophiognomonia castaneae;Phomopsis castaneae-mollissimae;Coordinated pathogenic effect
板栗褐缘叶枯病是近几年在我国板栗产区新发生的一种严重的叶部病害,其病原菌为板栗拟茎点霉菌Phomopsis castaneae-mollissimae(代号Y1,下同)[1-3]。在对板栗褐缘叶枯病病原菌分离的过程中发现,病斑上着生着另一真菌——Ophiognomonia castaneae(代号Y6,下同),该菌在病健交界处分离到的频率仅次于致病菌,且发病部位产生大量Y6的无性子实体(分生孢子盘),很容易被误认作褐缘叶枯病病原菌。产生Y6子实体的病斑症状、病斑大小与褐缘叶枯病病斑并无明显区别,交叉混杂在一起,且二者发生的时间较为一致,但将Y6单独在室内和林间接种,并不产生板栗褐缘叶枯病的症状。
真菌对营养的需求不同或不同生长阶段对营养的需求不同,造成了部分病原菌活动及致病机理的复杂性。根据科赫假定的原则,一种病害的致病菌只有一种,但现今的科学研究表明,有些病害的致病菌并非是单一病原菌[4,5],同一属内不同种类的真菌交互致病的现象较为常见,不同科属真菌协同侵染的研究报道却较少。本试验研究了Y6和Y1对寄主的侵染关系,以期为板栗褐缘叶枯病的发病规律、预防和治理提供理论依据。
1 材料与方法
1.1 供试菌株
板栗褐缘叶枯病病害标本采自山东省泰安市徂徕山林场光华寺林区。经组织分离、纯化培养得到Y1和Y6菌株,4℃冰箱保存备用。
1.2 试验方法
1.2.1 不同时期Y6与Y1分离频率 于2012、2013年7月下旬-10月中旬采集带病斑的叶片,在病健交界处切2 mm叶片组织,经冲洗、消毒,斜插到PDA平板培养基上,置于25℃培养箱培养。每次分离100个点,根据菌落形态差异,统计48 h分离到的两种菌在总分离物中的比例。
1.2.2 Y6子实体着生位置与板栗褐缘叶枯病病斑的关系 于2012年和2013年9-12月在林间采集病叶,体视镜下观察Y6无性型分生孢子盘在病叶上的分布特点,次年5-6月采集发病区上年落叶,观察有性型子囊壳的分布特点。
1.2.3 Y6 和Y1致病性测定 采用离体叶片接种法。于2013年8-9月份采集新鲜无病斑叶片,在 15 cm培养皿中铺6层灭菌纱布,无菌水浸湿。将一叶片放入皿中,用消毒昆虫针刺伤叶片,分别接种直径5 mm 的Y6、Y1及Y6和Y1各1/2的合并菌饼。每叶接4个点,2片叶为一组,对照组接空白培养基。置于昼30℃14 h、夜25℃10 h人工气候箱,湿度80%,48 h后去除菌饼。于接种后7、14 d观察病斑产生情况。
1.2.4 Y6与Y1的营养体亲和性 在PDA平板培养基上,一侧接种Y6菌饼,25℃下培养7 d后,再在另一侧接种Y1菌饼,菌饼间距1.5 cm,设5个重复。25℃条件下培养,10 d后观察对峙培养结果。
2 结果与分析
2.1 不同时期分离Y6与Y1的频率
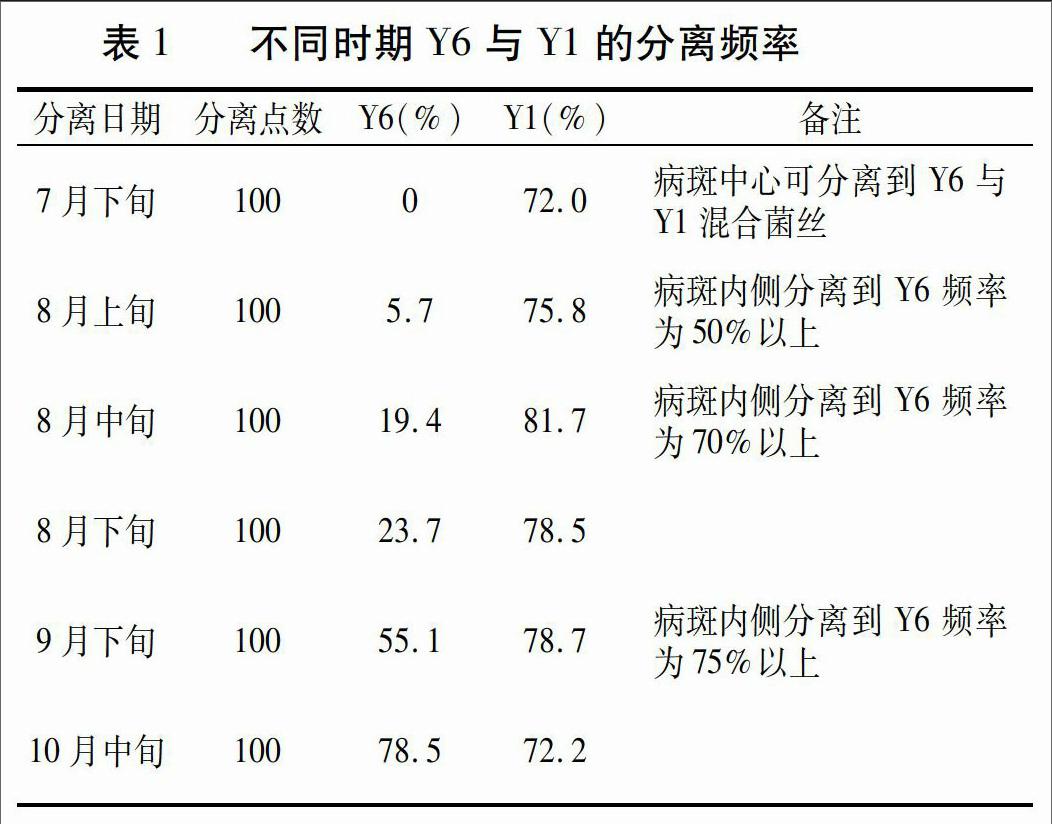
Y6与Y1两菌落形态有明显差异。Y6菌落形态具多样性,多表现为菌落较厚,毡状,边沿形状不规则,气生菌丝稀疏白色,生长缓慢(4周左右菌落直径3~4 cm),初分离得到的菌落易产生浅黄色、半固体粘稠的多糖类分泌物。Y1菌落的分离物先产生一层无色、透明、湿润的基生菌丝, 3~4 d后长出浓密气生菌丝,菌丝白色,生长速度快,后期为淡灰色或褐灰色,多数在平板易产生 3~4个界限明显的环痕[1]。不同时期Y6与Y1的分离频率见表1。
7月下旬,病区少量叶片正面初见小病斑,病健交界处组织分离培养,分离到大量Y1;8月上旬,病斑面积开始扩展,病健交界处Y6和Y1的分离频率为5.7%和75.8%,Y1的数量占绝对优势,但在病健交界处至病斑中心Y6分离频率可达高到50%以上(多数为Y1和Y6的混合菌丝);8月中旬病斑数量明显增长,病害进入快速增长期;8月下旬至9中、下旬病害进入发病高峰,病斑面积和数量都在不断扩大,部分感病严重的叶片病斑开始连接成片,发病部位焦枯,Y6菌株的分离频率逐渐增加;10月中旬,Y6菌株的分离频率超过Y1,且时间越晚Y6菌株的分离频率越大。
2.2 Y6子实体着生位置与板栗褐缘叶枯病病斑的关系
10月底开始,板栗病叶落叶前焦枯的叶片及落叶后干枯的叶片,其叶背面沿病斑边沿分布一圈密集深褐色的圆形小点,即Y6分生孢子盘,初期只着生在病斑边沿,形成环形,后期病斑中部也开始形成,随后所有病斑上均可见到Y6无性型子实体。病斑边沿处分生孢子盘较大,饱满,密集分布,环形明显;病斑内分生孢子盘相对稀疏,较小,分布较均匀。非病斑处及叶正面病斑处未发现分生孢子盘,如图1A 。
病叶脱落后,最早在当年11月下旬,湿度较大或有水浸湿的叶片背面病斑处长出Y6未成熟子囊壳,至次年5~6月,子囊壳大量产生,从成熟子囊壳顶端生长出黑色长喙,密集分布在叶背面病斑上,其在叶片上的分布规律相似于无性型子实体,如图1B。
2.3 Y6 和Y1的致病性
8月份致病性测定结果表明,接种Y6菌7 d后,大多不产生病斑,偶尔有很小的(<0.1 cm)病斑产生,14 d后致病率14.1%,病斑一般小于0.2 cm;接种Y1菌7 d后,大多接种点产生小于1 cm病斑,14 d后致病率81.3%,但病斑扩展较小,多在1 cm左右;同时接种Y6和Y1两菌7 d后,接种点大多产生大于1 cm的病斑,14 d后全部接种点都产生病斑,病斑直径一般2~3 cm。CK组14 d后仍不产生病斑(见图2、表2)。
9月份致病性测定表明,接种Y6菌与8月份致病结果近似,但14 d后,致病率提高到26.6%;接种Y1菌7 d后,接种点大部产生病斑,病斑迅速扩展,形成2~3 cm不规则病斑,少数接种点病斑连接成片至整个叶片,14 d后,病斑扩展至全叶并焦枯;接种Y6和Y1合并菌7 d后, 接种点产生较大病斑,且多连接成片,病斑处叶片开始焦枯,14 d后,病斑扩展至整片叶,并产生严重的焦枯现象;CK组14 d后仍不产生病斑(见表2)。
2.4 Y6与Y1营养体亲和性
结果显示,接种后Y1菌丝迅速生长到Y6菌丝边沿,后出现短暂的停滞现象,菌丝整齐,后迅速覆盖到Y6菌落上。菌落交界处均表现为菌丝变致密,无色素产生,形成一条由致密的气生菌丝集结的隆起带,隆起带偏于菌落生长较慢的Y6菌落一侧,如图3所示。表明,Y6与Y1的营养体表现出完全的亲和性,这与两菌在板栗褐缘叶枯病上共同侵染、形成混合病斑的现象是一致的。
3 讨论与结论
不同时期Y6与Y1的分离频率表明,前期病健交界处以Y1为主,其分离频率呈现先增长后减少的趋势,Y1先侵染使叶片产生病斑,当叶片病斑扩展至2~3 mm时,在病斑中心可分离到Y6。病害发生的中、后期Y6的比例逐渐增大,最终在数量上超过Y1,成为病健交界处的优势菌株。Y6的有性型和无性型子实体均生长在Y1引起的病斑上,而在无Y1病斑的叶片上未发现有Y6子实体。可以推断:①Y1具有强的致病性和寄生性,Y6具有弱的致病性和较强的腐生性;② Y6需要在Y1侵染形成的病斑上产生繁殖结构繁衍后代;③ Y6通过增强Y1致病力与Y1协同致病,通过加强对叶片的危害来扩大其栖息场所从而更好地生长和繁衍。
8月份和9月份两次致病性测定结果有一定差异,分析认为,8月份板栗叶片生长旺盛,营养较为丰富,抗性较强,而此时自然界病原菌刚侵染叶片产生病斑,因此对叶片接种产生病斑较小,扩展较慢,叶片不易焦枯。9月份板栗树处于生殖生长后期,叶部营养物质减少,抗病力降低,易受病菌侵染而发病。离体叶片接种结果表明,Y1菌致病力强,病斑产生快,Y6致病力很弱,二者同时接种致病力大于单独接种Y1菌,表现出协同侵染的特点。Y6与Y1菌株的营养体亲和性强,间接证明二者可以融合生长在一起。有关协同致病、协同侵染或复合侵染的报道多见于真菌与昆虫[6]、真菌与线虫[7,8]、病毒与病毒[9-11]之间,而真菌与真菌的相关报道较少。2种及其以上病原物侵染寄主的方式有相继协同作用(sequential synergism)和相伴协同作用(concomitant synergism)两种[12],初步断定Y6与Y1的关系属于前者,侵染过程中Y1承担着初始病原(primary pathogens)的角色,而Y6则扮演了继入病原(secondary pathogens)的角色,但是两菌的协同侵染机制和过程等还需要进一步探索。
参 考 文 献:
[1] 姜淑霞, 马洪兵. 板栗上的拟茎点霉一新种[J]. 菌物学报, 2010, 29(4): 467-471.
[2] 姜淑霞, 刘传忠, 王庆华, 等. 板栗新病害褐缘叶枯病及病原鉴定[J].林业科学, 2011, 47(5): 177-180.
[3] 姜淑霞, 贾雯, 邵云华, 等. 板栗褐缘叶枯病病原菌生物学特性研究[J]. 山东农业科学, 2010 (4): 52-55.
[4] 曹茂开,汪美先. 科赫氏法则在现代医学中的新思考[J]. 医学与哲学(人文社会医学版),1991(12):8-10.
[5] 喻盛甫,胡先奇,王扬. 包含病原线虫的植物复合侵染病害[J]. 植物病理学报,1999,29(1):1-7.
[6] 宋立秋,魏利民,王振营,等. 亚洲玉米螟与串珠镰孢菌复合侵染对玉米产量损失的影响[J]. 植物保护学报,2009,36(6):487-490.
[7] Smith A L. The reaction of cotton varieties to Fusarium wilt and root-knot nematode[J]. Phytopathology, 1941,31:1099-1107.
[8] Porter D M, Powell N T. Influence of certain Meloidogyne species on Fusarium wilt development in flue-cured tobacco[J]. Phytopathology, 1967,57:282-285.
[9] 方琦,董家红,郑宽瑜,等. 番茄环纹斑点病毒与马铃薯Y病毒复合侵染烟草的细胞病理特征[J]. 植物学报,2014,49(6):704-709.
[10]青玲,牛颜冰,吴楚钊,等. 部分烟区烟草病毒病病原检测及复合侵染分析[J]. 烟草科技,2009(12):58-64.
[11]谢礼,吕明芳,王芳,等. 甘薯羽状斑驳病毒和甘薯褪绿矮化病毒复合侵染甘薯引起的细胞病理学研究[J]. 电子显微学报,2013,32(6):485-491.
[12]Powell N T. Internal synergisms among organisms inducing disease[J]. Plant Disease, 1979,4:112-133.
猜你喜欢
农民致富之友(2020年31期)2020-11-18
食品安全导刊(2020年6期)2020-06-15
中国瓜菜(2019年8期)2019-09-19
江苏农业科学(2017年9期)2017-07-15
湖北农业科学(2017年4期)2017-03-28
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
江苏农业科学(2015年9期)2015-10-20
中国质量万里行(2015年1期)2015-01-27
食品与生活(2014年8期)2014-10-20
