
不同抗寒性白菜型冬油菜杂交后代保护性酶活性与越冬率分析
2016-12-17 03:02王凯音孙万仓刘自刚郭仁迪
西南农业学报 2016年11期
王凯音,孙万仓,刘自刚,方 彦,杨 刚,马 骊,郭仁迪,
方 圆,侯献飞,刘林波,钱 武, 刘海卿, 袁金海,王治江
(甘肃省油菜工程技术研究中心,甘肃省干旱生境作物重点实验室/甘肃省作物遗传改良与种质创新重点实验室,甘肃农业大学农学院,甘肃 兰州 730070)
不同抗寒性白菜型冬油菜杂交后代保护性酶活性与越冬率分析
王凯音,孙万仓*,刘自刚,方 彦,杨 刚,马 骊,郭仁迪,
方 圆,侯献飞,刘林波,钱 武, 刘海卿, 袁金海,王治江
(甘肃省油菜工程技术研究中心,甘肃省干旱生境作物重点实验室/甘肃省作物遗传改良与种质创新重点实验室,甘肃农业大学农学院,甘肃 兰州 730070)
以超强抗寒性冬油菜品种陇油7号分别与抗寒性品种延油2号和耐寒性品种天油2号的杂交后代(F1、F2、BC1)为研究对象,对各世代越冬率进行统计并结合亲本及杂交后代的CAT、POD、SOD酶活性变化研究,分析它们之间的抗寒性关系。结果表明:不同抗寒性品种杂交后代的越冬率均介于2个亲本之间,但不同世代越冬率存在较大差异;母本的抗寒性越强,在F1代中,后代的越冬率越高,F1代的自交后代(F2)越冬率比F1代的低;在BC1中,后代的越冬率变化范围较大,既有低于F2代自交后代的杂交组合(天2×陇7)×天2,也有高于F1代的杂交组合(陇7×天2)×陇7,以抗寒性强的品种作轮回亲本可以使回交一代的越冬率明显升高。低温胁迫后,2个群体不同世代的保护酶活性均增加,在群体1中,F1代CAT平均值从降温后比降温前升高了,F2从8.41 U·g-1到15.72 U·g-1,BC1从10.62 U·g-1到19.20 U·g-1。对越冬率与保护性酶进行回归分析,回归方程为Y=1.208+2.698X1+1.154X2+0.163X3,可得出越冬率与CAT酶、POD酶、SOD酶活性的关系呈极显著正相关,说明保护性酶活性越强,越冬率越高,抗寒性也就越好。
冬油菜;白菜型;越冬率;保护酶活性;抗寒性;杂交后代
油菜是我国第一大油料作物,随着冬油菜北移融入北方地区的种植制度,有关油菜抗寒性研究逐渐为人们关注,目前,对白菜型冬油菜抗寒性的研究主要集中在冬油菜品种的适应性[1]、生长发育特性[2]、产量及经济特性方面[3-4]。陈姣荣[5]对白菜型冬油菜在北方地区的越冬率、生育期及产量进行比较,以分析其适应性。结果表明,不同冬油菜品种越冬率存在较大差异,孙万仓[2]对北方旱寒区北移冬油菜生长发育特性研究结果表明,冬油菜北移后越冬率降低;生育期延长。由于生长在相对恶劣的气候生态条件下,越冬期漫长而极端低温低,北移冬油菜栽培品种必须具备优异的抗寒性,以确保安全越冬。李明玉[6]研究发现,低温锻炼能激活POD、SOD和CAT的活性,增强作物对冷害的抗性。蒲媛媛[7]研究表明在低温胁迫条件下,抗寒性强的品种体内保护酶类活性较高是增强其抗寒性的原因之一。品种的抗寒性是植物体内一系列生理生化变化的结果,低温逆境中CAT、POD、SOD活性水平与植物的抗寒性强弱具有十分密切的关系,并可以作为植物抗寒性检测的生理指标。而目前对不同抗寒性白菜型冬油菜杂交后代保护性酶活性与越冬率分析的研究鲜见报道。本实验以超强抗寒性冬油菜品种陇油7号与抗寒性冬油菜品种延油2号和耐寒性冬油菜天油2号的杂交后代为研究材料,对不同抗寒性白菜型冬油菜杂交后代保护性酶活性与越冬率的关系进行分析,为北方地区冬油菜抗寒育种提供根据。
1 材料与方法
1.1 试验材料
陇油7号为超强抗寒性品种,由甘肃农业大学育成;延油2号为抗寒性品种,由延安农科所提供;天油2号为耐寒性品种,由天水农科所提供。各世代材料种植情况列于表1。
1.2 试验设计
试验材料均于2013年8月20日播种于兰州市上川基地。试验地前茬为小麦(Triticumaestivu),施农家肥60 000 kg·hm-2、磷酸二铵300 kg·hm-2。田间管理按大田常规生产进行。分别于常温(2013年 10 月 5日,11~26 ℃)和低温胁迫后(2013 年 11 月21日,-3~11 ℃)按单株取样,取同一位点生长健壮的功能叶片,测定其生理生化指标,每个单株重复3 次。
1.3 测定指标与方法
1.3.1 生理生化特性测定 过氧化氢酶(CAT)采用紫外吸收法,以 1 min 内A240减少0.1的酶量为 1 个酶活单位;过氧化物酶(POD)活性测定采用愈创木酚比色法,以每分钟内A470变化 0.01 为1个过氧化物酶活单位;超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光还原法,1个酶活单位定义为将 NBT 的还原抑制到对照的一半时所需的酶量。上述 3 个指标均参照文献[8]进行。

表1 试验材料
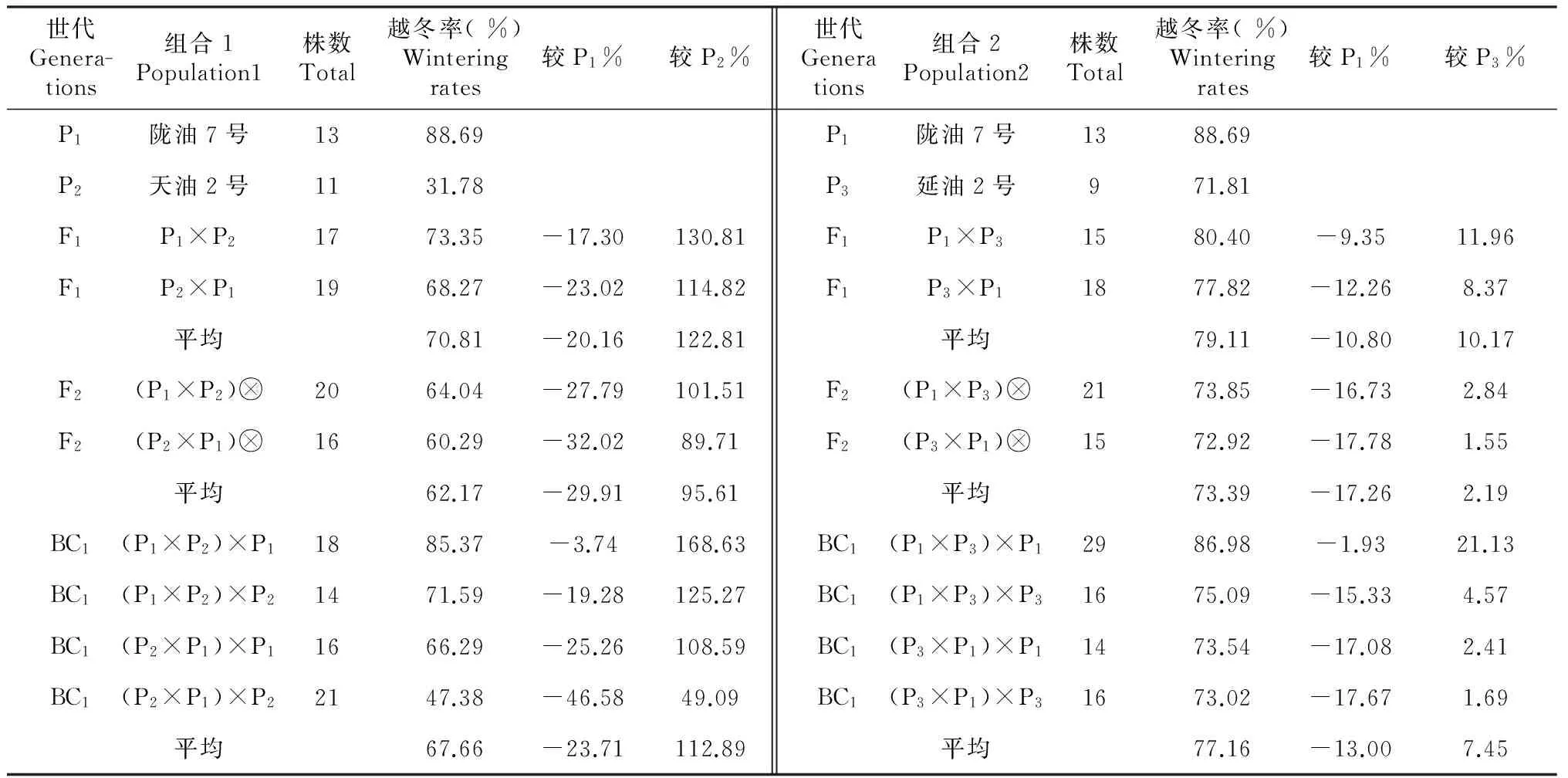
表2 不同世代越冬率
1.3.2 越冬率统计 冬前统计群体株数,第2年返青后统计群体存活株数。
越冬率=第2年返青后群体存活株数/冬前统计群体株数×100 %
1.3.3 数据处理 试验数据采用 Excel 2010整理,回归分析采用SPSS19.0[9]软件处理。
2 结果与分析
2.1 白菜型冬油菜不同抗寒性品种杂交后代越冬率变化
如表2所示,陇油7号、延油2号、天油2号越冬率分别为88.69 %、 71.81 %、31.78 %。3个亲本越冬率与抗寒性一致。
由杂交后代越冬率与亲本越冬率的比较得出:不同抗寒性品种杂交后代的越冬率均介于2个亲本之间;但不同世代越冬率存在较大差异,越冬率平均值比较F1代(70.81 %)>BC1代(67.66 %)>F2代(62.17 %),F2代越冬率较F1代降低;同一世代间由于母本与轮回亲本的不同,越冬率也不同,F1代,陇7×天2的越冬率为73.35 %,天2×陇7越冬率为68.27 %,F2代,(陇7×天2)自交的越冬率为64.04 %,(天2×陇7)自交越冬率为60.29 %,BC1代,(陇7×天2)×陇7的越冬率为85.37 %,(陇7×天2)×天2的越冬率为71.59 %;从而表明,做母本的抗寒性越强,后代的抗寒性越大,做轮回亲本的抗寒性越强,后代的抗寒性也就越强,由于白菜型油菜的高度不亲和性,使得自交后代的抗寒性明显降低。
2.2 白菜型冬油菜不同抗寒性品种杂交保护性酶活性变化
2.2.1 白菜型冬油菜不同抗寒性品种杂交后代过氧化氢酶(CAT)活性变化 CAT是生物体内重要的H2O2清除酶之一[10]。从表3可以看到,低温胁迫后不同抗寒性品种杂交后代 CAT 酶活性均显著高于胁迫前。低温胁迫后,超强抗寒亲本陇油7号为 27.50 U·g-1,抗寒亲本延油2号为18.34 U·g-1,耐寒亲本天油2号为15.45 U·g-1,逐渐减小,与胁迫前的变化情况相同,即随着亲本抗寒性的减弱,CAT 酶活性也逐渐减小。低温胁迫后不同抗寒性品种杂交后代的CAT 酶活性与亲本相比较介于两个亲本之间,但不同世代存在差异,在F1代中,陇7×天2为20.40 U·g-1,天2×陇7为18.36 U·g-1,明显高于F2代(陇7×天2)自交为15.80 U·g-1,(天2×陇7)自交为15.64 U·g-1,BC1中,同一世代间差异较大,酶活性最高的(陇7×延2)×陇7为27.07 U·g-1,与亲本相比较,只比超强抗寒性亲本的陇7低1.56 %,比另一强抗寒性的亲本延2高出47.60 %,酶活性最低的(天2×陇7)×天2为15.49 U·g-1,比陇7低43.67 %,比天2高7.20 %,由此说明自交使后代的酶活性降低,在回交一代中抗寒性越强的品种作为母本和轮回亲本,后代的酶活性越高,越接近抗寒性强的亲本。
2.2.2 白菜型冬油菜不同抗寒性品种杂交后代过氧化物酶(POD)活性变化 POD是植物活性氧代谢中活性氧清除系统的一种重要酶类[11]。从表4 中看到,低温胁迫后不同抗寒性品种亲本的 POD 酶活性均升高,陇油7号胁迫前62.23 U·g-1,低温胁迫后升至89.02 U·g-1,延油2号和天油2号分别从45.27 和 35.67 U·g-1增高到60.25 和 48.33 U·g-1,无论低温胁迫前后F1代酶活性的平均值都高于F2代的,降温后F1代中,酶活性最高的陇7×延2为69.08 U·g-1明显高于酶活性最低的天2×陇7(64.67 U·g-1),BC1世代间的酶活性差异较大,在62.35 ~75.3 U·g-1范围内变化。由此说明,F1代的酶活性变化范围较BC1代小,相对较稳定。

表3 不同世代 CAT 活性变异
2.2.3 白菜型冬油菜不同抗寒性品种杂交超氧化物歧化酶(SOD)活性变化 SOD与过氧化物酶、过氧化氢等酶协同作用防御活性氧或过氧化物自由基对细胞膜系统的伤害[12]。在表5中,由胁迫后/胁迫前的比值可看出,低温胁迫后不同抗寒性品种的超氧化物歧化酶(SOD)活性明显升高,亲本的酶活性差异与其抗寒性表现差异相同,BC1代中,不同抗寒性亲本的不同组合酶活性不同,以强抗寒性亲本陇7作为母本和轮回亲本[(陇7×天2)×陇7]的酶活性为645.25 U·g-1远远高于以弱抗寒性的亲本天2作为母本和轮回亲本[(天2×陇7)×天2]的酶活性为612.03 U·g-1。
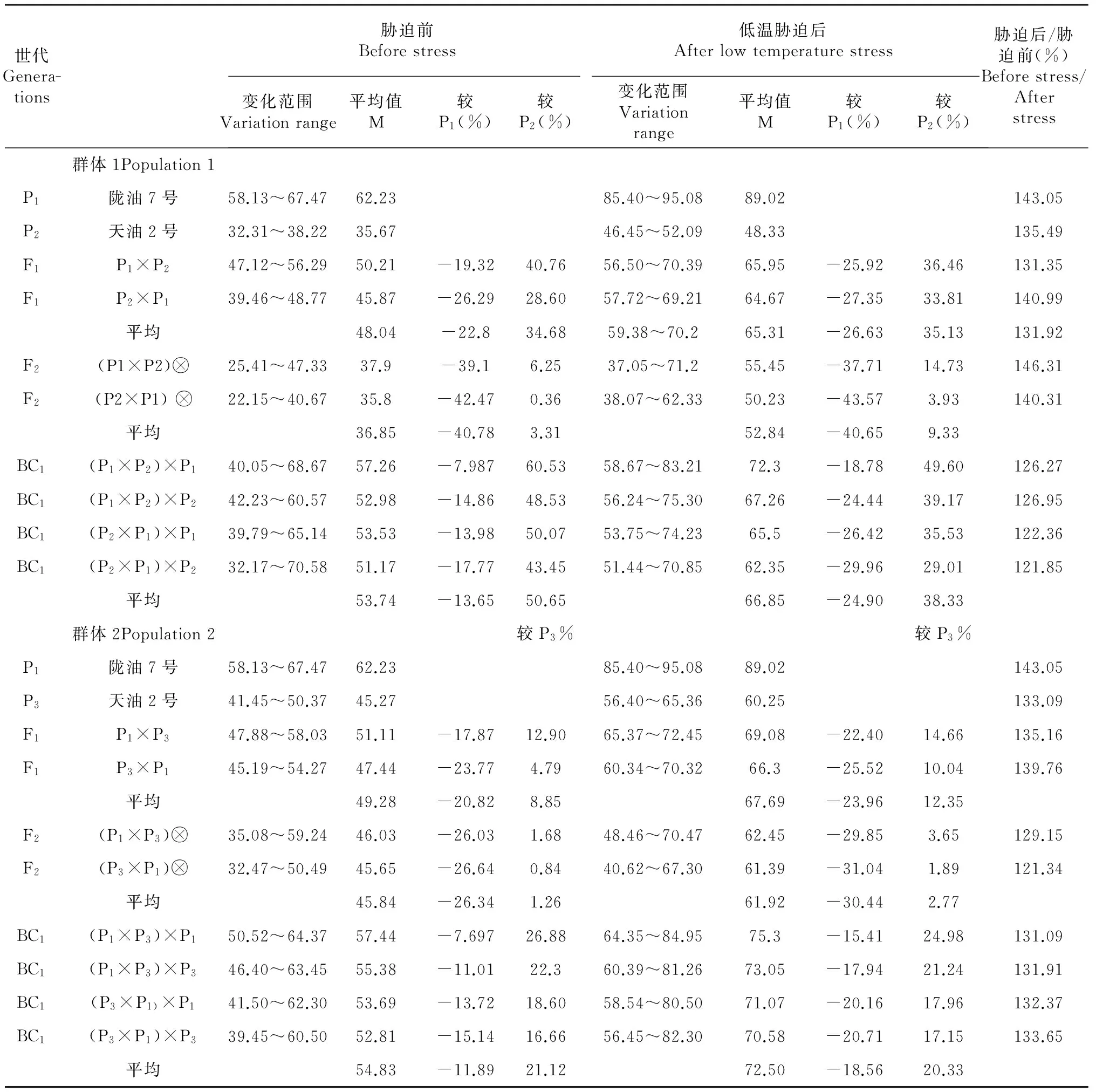
表4 不同世代 POD 活性变异
2.3 越冬率与保护性酶的回归分析
对越冬率与保护酶低温胁迫前后的差值进行回归分析,得出回归方程Y=1.208+2.698X1+1.154X2+0.163X3,其中Y为越冬率,X1为CAT活性,X2为POD活性,X3为SOD活性。经检验,回归方程表示X1、X2、X3、Y的关系达极显著(方差分析表Sig.=0.000),X1对Y影响显著(系数表Sig.=0.026≤0.05)。X1、X2、X3对Y的偏回归系数分别为2.698、1.154、0.163与越冬率呈正相关,复相关系数R=0.835。
3 讨 论
越冬率是反应抗寒性最为直观可靠的指标。本研究通过对白菜型冬油菜不同抗寒性品种杂交后代越冬率分析,发现随着3个亲本陇油7号、延油2号、天油2号的抗寒性依次减弱,它们的越冬率也逐渐减小,杂交后代(F1、F2、BC1)的越冬率都介于两个亲本之间,在F1代中,作母本的品种抗寒性越强后代的抗寒性越强。F2代自交后代抗寒性明显比F1代差,由于白菜型冬油菜高度不亲和,导致自交后代的抗寒性降低。BC1中,以抗寒性越强的亲本作母本和轮回亲本,后代的抗寒性越强,出现了抗寒性很接近超强抗寒性亲本的后代,说明以超强抗寒性亲本作为轮回亲本进行回交可有效提高杂交后代的抗寒性。
CAT、POD和 SOD统称保护酶, 在冷胁迫下清除活性氧 ,防止膜脂过氧化及抗低温伤害具有重要作用[10-12],植物体中保护酶活性的强弱直接关系到抵御低温伤害的能力[13]。在本实验中,降温后,CAT 、POD和 SOD 3个保护酶活性均增强,说明低温胁迫后,白菜型冬油菜通过提高自身的保护酶活性来抵御低温带来的伤害。杂交后代的酶活性都低于抗寒性强的亲本,高于抗寒性弱的亲本,但不同世代存在差异,F2自交后代的酶活性比F1代低,在BC1中,世代间的差异较大,既有酶活性高于F1代以抗寒性强的品种为轮回亲本的杂交后代也有低于F2代以抗寒性弱的品种为轮回亲本的杂交后代,从而说明应该以抗寒性强的品种为轮回亲本用正确的回交方式来提高杂交后代的酶活性,以提高它的抗寒性。
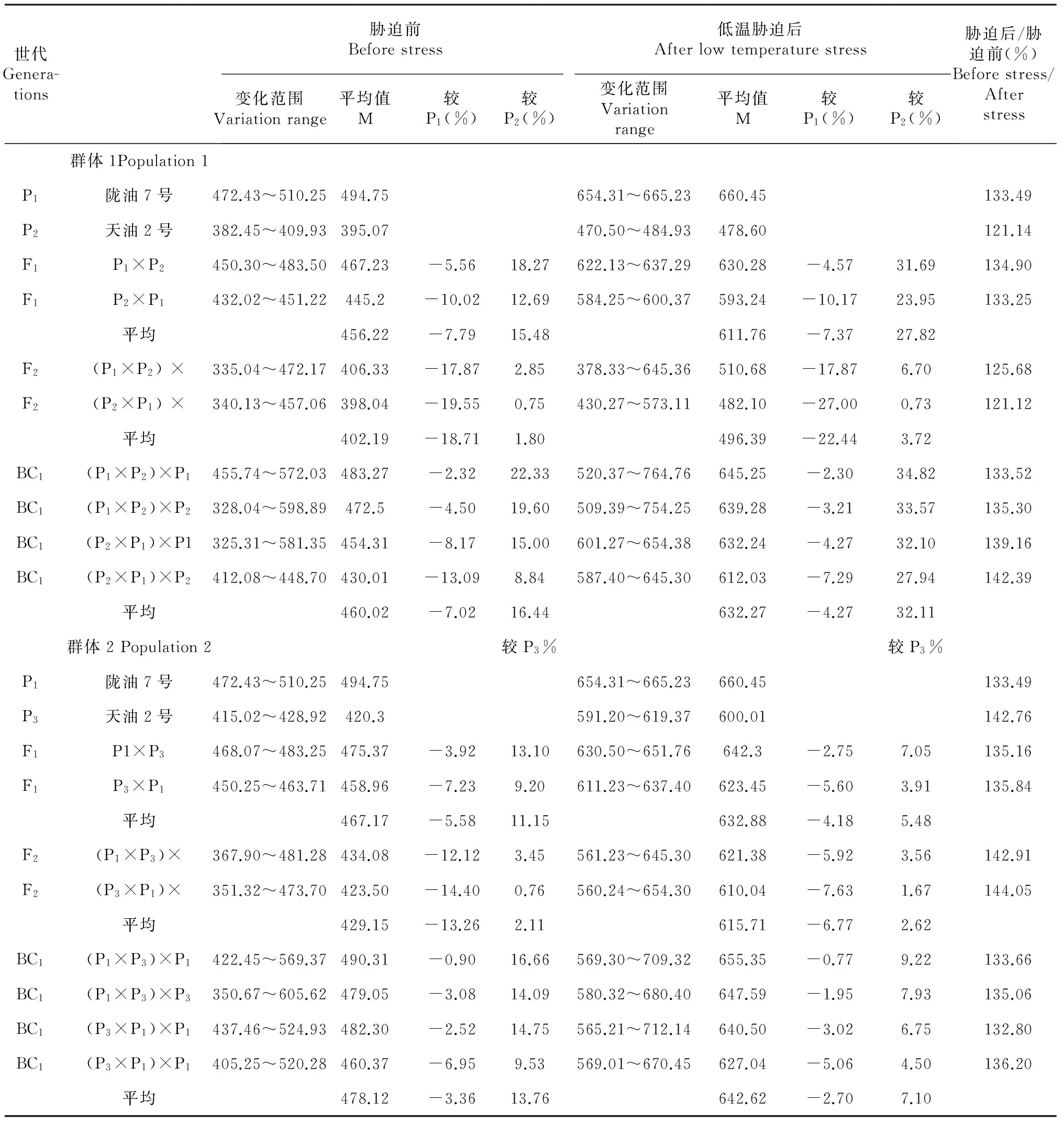
表5 不同世代 SOD活性变异
对越冬率与保护酶进行回归分析,可得出越冬率与CAT酶、POD酶、SOD酶的关系极显著,说明保护性酶活性越强,越冬率越高,抗寒性也就越好。
用抗寒性强的品种作母本可有效的提高杂交一代的保护酶活性及越冬率,自交使后代的保护酶活性及越冬率降低,以抗寒性强的亲本作为轮回亲本进行回交的后代的酶活性和越冬率显著升高,因此,以抗寒性强的品种作为母本及配合正确的杂交方式可有效提高后代的保护酶活性和越冬率,抗寒性更好。
[1]朱惠霞, 孙万仓, 邓 斌, 等. 白菜型冬油菜品种的抗寒性及其生理生化特性[J]. 西北农业学报, 2007, 16(4):34-38.
[2]孙万仓, 武军艳, 方 彦, 等. 北方旱寒区北移冬油菜生长发育特性[J]. 作物学报, 2010, 36(12):2124-2134.
[3]魏文慧, 孙万仓, 郭秀娟, 等. 氮磷钾肥对西北寒旱区冬油菜越冬率、产量及经济性状的影响[J]. 西北农业学报, 2009, 18(2):122-125, 130.
[4]方 彦, 孙万仓, 武军艳, 等. 叶面喷施硼肥对西北旱寒区冬油菜保护酶、产量及经济性状的影响[J]. 草业科学, 2012, 29(9):1446-1450.
[5]陈姣荣, 孙万仓, 方 彦, 等. 白菜型冬油菜在北方寒旱区的适应性分析[J]. 干旱地区农业研究, 2012, 30(6):17-22, 31.
[6]李明玉, 曹辰兴, 于喜艳. 低温锻炼对冷胁迫下黄瓜幼苗保护性酶的影响[J]. 西北农业学报, 2006, 15(1):160-164.
[7]蒲媛媛, 孙万仓. 白菜型冬油菜抗寒性与生理生化特性关系[J]. 分子植物育种, 2010, 8(2):335-339.
[8]徐向宏. 试验设计与 Design-Expert. SPSS应用[M]. 北京:科学出版社, 2010:69-71.
[9]刘鸿先, 曾韶西, 李 平, 等. 零上低温对不同耐冷力的亚热带植物过氧化物酶与酯酶同工酶的影响[J]. 植物生理学报, 1981, 7(4):337-344.
[10]李秀菊, 丁钟荣. 抗寒锻炼对不同小麦品种质膜及其保护酶的影响[J]. 河南职技师院学报, 1991, 19(2):1-5.
[11]刘鸿先, 曾韶西, 王以柔, 等. 低温对不同耐寒力黄瓜幼苗子叶各细胞器官中SOD的影响[J]. 植物生理学报, 1985, 11(1):48-57.
[12]王建华, 刘鸿先, 徐 同, 等. 超氧化物歧化酶(SOD) 在植物逆境和衰老生理中的作用[J]. 植物生理学通讯, 1989(1):1-7.
(责任编辑 李山云)
Analyses of Protective Enzyme Activity and Wintering Rates in Filial Generations of Different Cold Resistance Winter Rapeseed (BrassicarapaL.)
WANG Kai-yin, SUN Wan-cang*, LIU Zi-gang,FANG Yan,YANG Gang, MA Li, GUO Ren-di, FANG Yuan,HOU Xian-fei, LIU Lin-bo, QIAN Wu, LIU Hai-qing, YUAN Jin-hai, WANG Zhi-jiang
(Campestrisseed Engineering Research Center of Gansu Province,Gansu Provincial Key Laboratory of Arid Land Crop Sciences,Improvement and Key Laboratory of Crop Genetics and Germplasm Enhancement,Gansu Agricultural University,Gansu Lanzhou 730070,China)
Taken the filial generations (F1, F2, BC1) of the ulterstrong cold resistance variety Longyou 7 with cold resistance variety Yanyou 2 and cold tolerance variety Tianyou 2 as tested materials, their each generation wintering rates were estimated, combining with their parents and filial generations CAT, POD, SOD enzyme activity change, and their cold resistance relationship between them were analyzed. The results showed that wintering rates in filial generations of different cold resistance winter turnip rape between the two parent, but there were large differences between different generations; In F1generation, the stronger the cold resistance of female parent was, the higher the wintering rate of offspring was, and F2generation of selfing generations of wintering rate was lower than that of the F1generation; In BC1generations, the wintering rate range was larger, there were lower than that of F2generation of selfing generations of hybrid combinations (Tianyou 2 × Longyou 7) × Tian you 2 and higher than that of F1generation hybrid combination (Longyou 7 × Tianyou) × Long you 7, and the variety with ultrastrong cold resistance as recurrent parent could make backcross generation wintering rate increase significantly. After low temperature stress, the protective enzyme activity of different generations of two groups increased to some extent, in group 1, the F1generation CAT average from before cool to after cooling was increased, F2from 8.41 to 15.72 U·g-1, and BC1from 10.62 to 19.20 U·g-1. By regression analysis for wintering rates and protective enzyme, their regression equation wasY=1.208+2.698X1+1.154X2+0.163X3, and the relationship of wintering rates with CAT , POD and SOD were significantly positive, indicating that the stronger the protective enzyme activity was, the higher the rate was, and the better the cold resistance was.
Winter rapeseed;BrassicarapaL.; Wintering rate; Protective enzyme activity; Cold resistance; Filial generations
1001-4829(2016)11-2529-07
10.16213/j.cnki.scjas.2016.11.004
2015-11-10
国家现代农业产业技术体系“北方寒旱区冬油菜育种岗位科学家”(CARS-13);国家自然科学基金“白菜型油菜抗寒评价指标体系及数学模型构建”(31460356);973计划-油菜高产油量形成的分子生物学机制(2015CB150206);国家863计划专项“强优势油菜杂交种的创制与应用”(2011AA10A104);国家农业科技成果转化项目(2014G10000317);国家自然科学基金(31560397);甘肃省自然科学基金(145RJZG050);甘肃省高等学校科研项目(2014B-049)
王凯音(1988-),女,甘肃兰州人,在读硕士,主要从事油菜育种工作,E-mail:444593310@qq.com,*为通讯作者:孙万仓,教授,博士,主要从事油菜育种及十字花科种质资源研究,E-mail:18293121851@163.com。
S634.3
A
猜你喜欢
今日农业(2022年1期)2022-11-16
电脑迷(2022年10期)2022-11-08
中国糖料(2021年3期)2021-07-13
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
西藏农业科技(2018年4期)2018-04-25
新疆农垦科技(2016年2期)2016-08-21
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
园艺与种苗(2015年10期)2015-02-27
