
Adenosine A2Areceptors in neuronal outgrowth: a target for nerve regeneration?
2016-12-02 07:05FilipaF.Ribeiro,AnaM.Sebastio
中国神经再生研究(英文版) 2016年5期
PERSPECTIVE
Adenosine A2Areceptors in neuronal outgrowth: a target for nerve regeneration?
Axonal and dendritic outgrowth are fundamental processes in the development of the nervous system. During this period, neurons change their morphology from a simple bipolar shape into a mature complex shape. Neurons develop dendrites and extend long or short axons that travel through a complex path until reaching target cells and form functional and accurate neuronal circuits. Throughout the way, dendrites and axons interact with several extracellular guidance cues (Figure 1), among which are adhesive molecules, either from the extracellular matrix or the surface of nearby cells, as well as diffusible chemotropic molecules that are secreted to the extracellular milieu (Maeda, 2015). Axonal interaction with extracellular guidance cues is possible because on the distal tip of the axon there is a specialized, highly dynamic and motile structure, called the growth cone. Dendrites also possess similar structures. Growth cones contain several different receptors on its membrane surface, as well as cell adhesion molecules that interact with those signalling cues present in the extracellular space, resulting in the activation of signalling cascades and consequent regulation of the cytoskeleton structure (Lowery and Van Vactor, 2009). Signalling cues can function as either attractants or repellents, promoting or inhibiting neurite growth, regulating neurite pathfinding, and neuronal arborisation. These different functions depend not only on the cue, but mostly on the receptor repertoire that is present and activated at each moment, as well as on the intracellular signalling cascades available to be activated in the growth cones (Kaplan et al., 2014; Maeda, 2015). Activation of these receptors will induce a rearrangement of the cytoskeleton thus promoting changes in neuronal morphology. As examples of diffusible chemotropic cues are morphogens and growth/neurotrophic factors such as brain-derived neurotrophic factor (BDNF). BDNF is widely expressed in the adult brain, and particularly highly expressed in the developing brain. This neurotrophin is released by neurons, microglia and astrocytes, and is involved not only in nervous system development, including the regulation of neuronal outgrowth, such as in dendritic and axonal growth and arborisation, but also in synaptic plasticity (Huang and Reichardt, 2001) and neuronal regeneration following peripheral and central nervous system injury (Huang and Reichardt, 2001; Ribeiro et al., 2015b). Synaptic actions of BDNF are modulated by adenosine, a purine nucleoside that is ubiquitously present in the brain and modulates several actions of other molecules, including synaptic activity (Sebastião and Ribeiro, 2009). During brain development, adenosine is highly expressed in the brain (Ribeiro et al., 2015b). Adenosine is released by neuronal cells during neuronal activity, being its intracellular concentration under the tight control of the activity of adenosine kinase. In the adult brain, this enzyme is mostly expressed in astrocytes, which plays a preponderant role in the levels of extracellular adenosine (Diógenes et al., 2014) since adenosine is released or taken up through equilibrative transporters located in the plasma membrane. Moreover, adenosine is also extracellularly generated from the hydrolysis of released adenine nucleotides. There are two high affinity adenosine receptors in the brain, the inhibitory A1receptor (A1R) and the excitatory A2Areceptor (A2AR). These receptors are G-protein coupled receptors and are present in the whole brain, although their density varies in different brain regions (Sebastião and Ribeiro, 2009). The more widely known neuromodulatory actions of adenosine are the control of synaptic function by either interfering with neurotransmitter release or neurotransmitter action at synapses. By using cortical neurons in culture we recently explored a much less known adenosine action, its neurotrophic potential. Both A2ARs and BDNF TrkB receptors are co-expressed during the development of primary cortical neurons (Ribeiro et al., 2015a). Considering the crosstalk between A2ARs and TrkB receptors and the influence of BDNF upon neuronal outgrowth, we hypothesized that A2ARs could have a role upon neuronal outgrowth. Remarkably, we could find out (Ribeiro et al., 2015a) that activation of A2AR promotes the elongation of the axon, as well as dendritic arborisation (Figure 2), being the effect of axonal elongation independent of endogenous BDNF actions. On the other hand, A2AR activation enhances BDNF-dependent dendritic branching (Ribeiro et al., 2015a), which means that adenosine, through A2AR activation, is able to act in a synergistic way with BDNF to control its neurotrophic actions. Potentiation of BDNF actions may involve the cAMP/PKA signalling pathway (Diógenes et al., 2004) and facilitation of TrkB signalling through transactivation of TrkB receptors and/or TrkB translocation to lipid rafts (Assaife-Lopes et al., 2014) or even the stimulation of BDNF mRNA and protein expression, as well as secretion of BDNF protein to the extracellular space, which in turn will act on TrkB receptors and thus promote changes in neuronal morphology (Ribeiro et al., 2015b). There are pathologies, as depression, where a decrease in neuronal arborisation occurs and that are associated to a decrease in BDNF levels and/or to a decrease in the levels of the signalling receptor for BDNF, the TrkB receptor. Indeed, several antidepressants promote BDNF expression and thus neuronal branching. One may thus hypothesize that A2AR selective agonists, by promoting BDNF action on neuronal morphology, may prove a good strategy to attenuate depression.
The ability of adenosine, through A2AR activation, to promote axonal elongation, is dependent on the activation of several signalling cascades known to be involved in the regulation of microtubule dynamics during axonal growth, such as phosphoinositide 3-kinase (PI3K), mitogen-activated protein kinases (MAPK) and phospholipase C (PLC) (Ribeiro et al., 2015a). Remarkably, these actions of adenosine are independent of BDNF (Ribeiro et al., 2015a). Adenosine may thus act as an extracellular guidance cue, directing the migration of axonal growth cones or even fine-tuning the actions of other cues. Indeed, A2ARs are positively coupled to adenylate cyclase and cyclic AMP which has an important role in modulating the growth cone turning responses in the presence of guidance cues (Ribeiro et al., 2015b).
The cytoskeleton of the growth cone is highly dynamic, making possible the exploration of the surrounding environment and axon guidance towards the appropriate targets. It is mainly formed by microtubules coming along the axon and splaying in the centre of the growth cone, as well as by actin fibres present in filopodia and lamellipodia structures in the peripheral region of growth cones (Lowery and Van Vactor, 2009). Rearrangements of the cytoskeleton comprise changes in actin and microtubule assembly and dynamics. In fact, dynamic microtubules in the growth cones may function as detectors of extracellular conditions, thus favouring process elongation or retractiondepending on the extracellular environment. Although several mediators are known to influence cytoskeleton rearrangements, the mechanisms behind the translation of the signalling cues into changes of the cytoskeletal machinery are largely unknown.

Figure 1 Migration of an axonal growth cone during brain development.
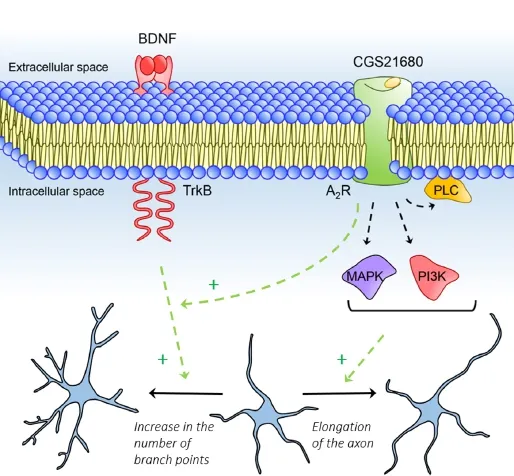
Figure 2 Adenosine A2Areceptor activation increases dendritic arborisation and axon extension.
Microtubules are tubular structures composed of heterodimers of α- and β-tubulin. Axonal elongation is achieved through microtubule assembly of tubulin heterodimers to the microtubule plus ends. During the process of axon growth, microtubules undergo cycles of polymerization (growth) and depolymerisation (shortening) at microtubule plus ends in the growth cone, a process called dynamic instability (Lowery and Van Vactor, 2009). By transfecting rat primary cultured cortical neurons with a construct of end-binding protein 3 (EB3), we were able to label growing microtubule plus ends and to find that A2AR activation increases the microtubule growth speed (Ribeiro et al., 2015a). Moreover, the axon has regions with different microtubule stability, whose regulation allows the axon to grow, turn and stabilize its structure. Microtubule dynamics and stability are affected by several post-translational modifications of tubulin. For example, post-translational tyrosinated α-tubulin is normally associated with dynamic microtubules, whereas detyrosinated and acetylated α-tubulin is associated with stable microtubules (Fukushima et al., 2009). Activation of A2ARs decrease the ratio acetylated α-tubulin/tyrosinated α-tubulin, which suggests an increase in microtubule dynamics that is central for microtubule extension and thus axonal growth (Ribeiro et al., 2015a). This is also consistent with reports showing that Gα subunit directly regulates the dynamic instability of microtubules required for neurite outgrowth, in a process independent of cAMP/PKA (Ribeiro et al., 2015b). Although we demonstrated that A2ARs promote microtubule instability, how cytoskeletal dynamics is regulated by A2AR activation in a context of neurite outgrowth is still unclear.Whether this action of A2ARs results from a direct interference of A2AR signalling cascades upon microtubules or through the action of other membrane signalling molecules also remains unknown. Nevertheless, as herein reviewed, evidences start to emerge that demonstrate the importance of adenosine, through A2AR activation, in processes of neuronal differentiation and neurite outgrowth, thus suggesting that A2AR play a trophic influence during neuronal maturation.
Signalling cues and cytoskeleton dynamics at the growth cones are not only important for neuronal outgrowth during development, but are also essential for neuronal regeneration. A2ARs may thus constitute a target for the development of future strategies for neuronal regeneration. The use of selective A2AR agonists would be suitable to regenerate circuits after a brain injury, in which it is necessary to guide new axons and to re-establish the dendritic and axonal arbour. Moreover, it could be also promising in several pathologies that encompass neuronal atrophy, including psychiatric diseases such as depression. However, care has to be taken in situations where A2ARs have their levels enhanced. With ageing and in conditions of psychiatric or neurologic diseases such as chronic stress, Parkinson's disease and Huntington's disease, the density of A2ARs may be increased in some brain areas and its activation can be deleterious (Sebastião and Ribeiro, 2009). An example is the stress-induced atrophy of dendritic arborisation, which can be reverted by A2AR blockade (Batalha et al., 2012).
Altogether, these observations highlight a key role of A2AR in neuronal arborisation, where a fine tuning action of A2AR may occur as a function of neuronal development and activity. By shaping A2AR activity it may be possible to promote neuronal morphological changes with impact in the function of neuronal circuits. Further studies are however required to better understand the consequences and mechanisms underlying the A2AR-mediated neurotrophic actions in the different brain areas and under different pathophysiologic conditions.
This work was supported by Fundacão para a Ciência e a Tecnologia (FCT) Grants∶ PTDC/SAU-NEU/64126/2006, FCOMP-01-0124-FEDER-017455 (HMSP-ICT/0020/2010), EXPL/BIMMEC/0009/2013, Portugal. Filipa F Ribeiro was in receipt of a fellowship (SFRH/BD/74662/2010) from FCT.
Filipa F. Ribeiro, Ana M. Sebastião*
Instituto de Farmacologia e Neurociências, Faculdade de Medicina, Universidade de Lisboa, Lisbon, Portugal; Instituto de Medicina Molecular, Faculdade de Medicina, Universidade de Lisboa, Lisbon, Portugal
*Correspondence to: Ana M. Sebastião, Ph.D., anaseb@medicina.ulisboa.pt.
Accepted: 2016-03-15
orcid: 0000-0001-9030-6115 (Ana M. Sebastião)
Assaife-Lopes N, Sousa VC, Pereira DB, Ribeiro JA, Sebastião AM (2014) Regulation of TrkB receptor translocation to lipid rafts by adenosine A2A receptors and its functional implications for BDNF-induced regulation of synaptic plasticity. Purinergic Signal 10:251-267
Batalha VL, Pego JM, Fontinha B, Costenla AR, Valadas J, Baqi Y, Radjainia H, Müller CE, Sebastião AM, Lopes LV (2012) Adenosine A2A receptor blockade reverts hippocampal stress-induced deficits and restores corticosterone circadian oscillation. Mol Psychiatry 18:320-331.
Diógenes MJ, Fernandes CC, Sebastiao AM, Ribeiro JA (2004) Activation of adenosine A2A receptorfacilitates brain-derived neurotrophic factor modulation of synaptic transmission in hippocampal slices. J Neurosci 24:2905-2913.
Diógenes MJ, Neves-Tomé R, Fucile S, Martinello K, Scianni M, Theofilas P, Lopatár J, Ribeiro JA, Maggi L, Frenguelli BG, Limatola C, Boison D, Sebastião AM (2014) Homeostatic Control of Synaptic Activity by Endogenous Adenosine is Mediated by Adenosine Kinase. Cereb Cortex 24:67-80.
Fukushima N, Furuta D, Hidaka Y, Moriyama R, Tsujiuchi T (2009) Post-translational modifications of tubulin in the nervous system. J Neurochem 109:683-93.
Huang EJ, Reichardt LF (2001) Neurotrophins: roles in neuronal development and function. Annu Rev Neurosci 24:677-736.
Kaplan A, Kent CB, Charron F, Fournier AE (2014) Switching responses: spatial and temporal regulators of axon guidance. Mol Neurobiol 49:1077-1086.
Lowery LA, Van Vactor D (2009) The trip of the tip: understanding the growth cone machinery. Nat Rev Mol Cell Biol 10:332-343.
Maeda N (2015) Proteoglycans and neuronal migration in the cerebral cortex during development and disease. Front Neurosci 9:98.
Ribeiro FF, Neves-Tomé R, Assaife-Lopes N, Santos T, Silva R, Brites D, Ribeiro JA, Sousa M, Sebastião AM (2015a) Axonal elongation and dendritic branching is enhanced by adenosine A2A receptors activation in cerebral cortical neurons. Brain Struct Funct doi:10.1007/s00429-015-1072-1.
Ribeiro FF, Xapelli S, Miranda-Lourenço C, Tanqueiro SR, Fonseca-Gomes J, Diógenes MJ, Ribeiro JA, Sebastião AM (2015b) Purine nucleosides in neuroregeneration and neuroprotection. Neuropharmacology doi:10.1016/ j.neuropharm.2015.11.006.
Sebastião AM, Ribeiro JA (2009) Adenosine receptors and the central nervous system. Handb Exp Pharmacol 193:471-534.
10.4103/1673-5374.182683 http∶//www.nrronline.org/
How to cite this article: Ribeiro FF, Sebastião AM (2016) Adenosine A2Areceptors in neuronal outgrowth: a target for nerve regeneration? Neural Regen Res 11(5):706-708.

- 中国神经再生研究(英文版)的其它文章
- Self-assembling peptide nanofibrous hydrogel as a promising strategy in nerve repair after traumatic injury in the nervous system
- Recovery of injured fornical crura following neurosurgical operation of a brain tumor: a case report
- Possible application of apolipoprotein E-containing lipoproteins and polyunsaturated fatty acids in neural regeneration
- Antibody-based neuronal and axonal delivery vectors for targeted ligand delivery
- Alzheimer's disease: the silver tsunami of the 21stcentury
- Clinical trial perspective for adult and juvenile Huntington's disease using genetically-engineered mesenchymal stem cells