
COXⅡ蛋白质分子确定哺乳动物物种分歧时间及其演化关系
2016-12-01 09:25:28李可群
西北师范大学学报(自然科学版) 2016年1期
李可群
(同济大学化学系,上海 200092)
COXⅡ蛋白质分子确定哺乳动物物种分歧时间及其演化关系
李可群
(同济大学化学系,上海 200092)
使用本文作者提出的生物分子绝对进化速率计算公式和COXⅡ蛋白质分子,计算得到了针鼹等单孔类哺乳动物于2.32亿年前分异出,这一结果与文献依据化石记录得出的结果相符;计算还得到袋鼩、袋鼹、负鼠和袋鼬的物种分歧时间均为2.0亿年前,袋鼠的物种分歧时间为1.39亿年前,这与两大类群分属于多门齿亚目和双门齿亚目以及两者于早期分化的文献报道结果相符.所得计算结果进一步支持哺乳动物进化的“兽类假说”,即哺乳动物的进化路径是从单孔类至有袋类再至胎盘类.若关于胎盘类哺乳动物单系类群假说正确的话,胎盘类哺乳动物起源时间不应早于袋鼠的物种分歧时间,即1.39亿年前.另外,计算还得到人与其他人猿总科成员,黑猩猩、大猩猩、猩猩和长臂猿的物种分歧时间分别为453.61万年前、814.02万年前、1 245.02万年前和1 822.40万年前,旧世界猴的物种分歧时间为3 934.736万年前,这些结果除黑猩猩等因缺乏早期化石外,均与它们的化石记录或文献结果基本相符.
哺乳动物;灵长动物;物种分歧时间;分子钟;进化速率;COXⅡ蛋白质
哺乳动物作为一类进化程度最高的动物类群,重建其进化历史和系统发育关系一直是系统发育学的核心问题,也是进化生物学研究的重要内容之一[1].动物化石首次出现可以反映该物种起源的最晚时间,但不一定等同于该物种的实际起始时间.多年来,国内外学者从解剖学、考古学等方面进行了研究,但有许多动物的分类学地位和进化关系仍无定义,存在许多争议[2].而目前迅速发展的“分子钟”理论由于一般未能考虑各支系分子进化速率的差异,得到的物种分歧时间一般早于化石记录,如真兽类哺乳动物目一级的物种分歧时间平均比化石早50%~90%.目前基于分子数据的系统发育研究方法还不能很好地阐明哺乳动物系统发育树基部问题以及哺乳动物进化过程中所经历的快速辐射事件[1].三大现生哺乳动物类群(单孔类、有袋类和胎盘类)进化关系一直是分类学家争议的问题,分子生物学研究结果存在关于哺乳动物演化顺序的2种假说,即认为演化顺序为单孔类→有袋类→胎盘类的“兽类假说”和认为单孔类与有袋类亲源关系更近,它们一起与胎盘类具有共同祖先的“Marsuipionta假说”[1].为了进一步弄清哺乳动物的演化关系,本文作者依据其提出的生物分子绝对进化速率计算公式,使用线粒体COXⅡ蛋白质分子得到了一些主要哺乳动物的物种分歧时间,它们与文献基于动物实体化石得到的结果相符.本文计算结果进一步支持关于哺乳动物进化的“兽类假说”.此外,本文作者还对人与其他一些灵长类动物,即黑猩猩、大猩猩、猩猩、长臂猿以及旧世界猴的物种分歧时间进行了计算,结果与化石记录或现有文献结果基本相符.
1 主要哺乳动物物种分歧时间及演化关系
1.1 同源生物分子绝对进化速率的计算公式
在研究生物分子进化时,作者给出了分子绝对进化速率的计算公式[3]:
(1)
其中,k1,k2分别为两个同源序列的分子绝对进化速率;t为从共同祖先序列进化的时间,或称两个物种的分歧时间;p为序列比对所得的种间差异率;k1t,k2t分别为两个同源序列的平均突变概率(或称遗传距离);d1,d2分别为两个同源序列与共同祖先序列相比的差异位点数;2n0为被比较2个同源序列氨基酸或核苷酸位点总数;d为两个同源序列中相同位点氨基酸(或核甘酸)不相同的位点数.为了计算和表述的方便,除特别申明,本文分子绝对进化速率的单位采用氨基酸(或核苷酸)位点数/10亿年,进化时间(或物种分歧时间)的单位采用10亿年.
1.2Kimura经典计算体系和犬科动物绝对进化速率的确定
为了说明生物分子进化速率的恒定性,著名分子进化学家Kimura在《The neutral theory of molecular evolution》[4]一书中给出了8种脊椎动物(鲨鱼、鲤鱼、蝾螈、鸡、针鼹、袋鼠、狗和人)的进化体系(图1).
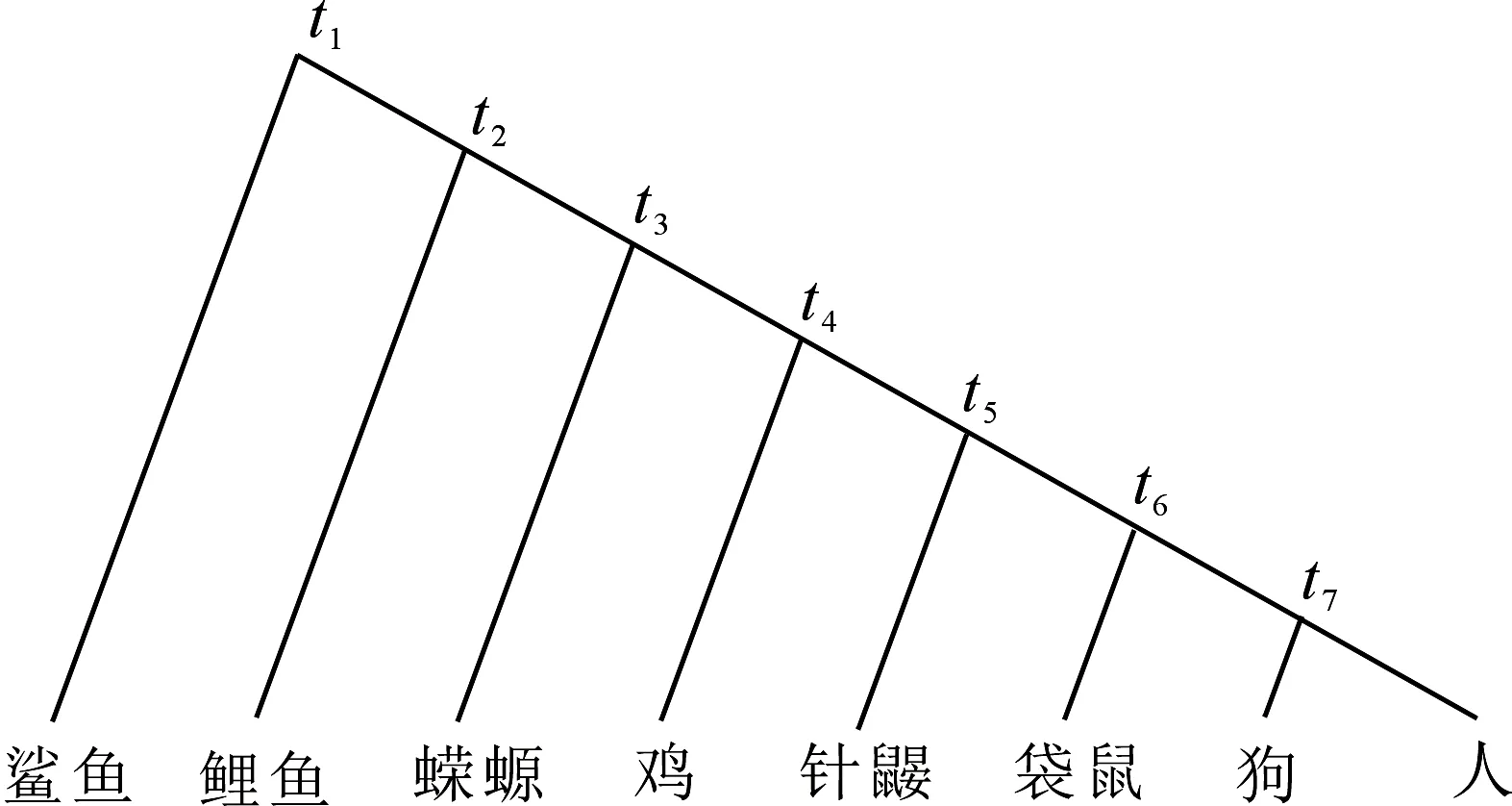
图1 八种脊椎动物的系统进化示意Fig 1Phylogenetic scheme of 8 kinds of vertebrates
Kimura给出了前7种脊椎动物根据化石确定的物种分歧时间,其数值分别为0.468,0.423,0.368,0.291,0.232,0.139和0.081,单位为10亿年.我们依据这些物种分歧时间,并使用我们提出的生物分子绝对进化速率计算公式以及生物分子绝对进化速率与物种分歧时间之间的定量关系公式,对前7种脊椎动物一些蛋白质和核苷酸序列分子的种间差异率数据进行了拟合,所得结果在未考虑多重突变校正的情况下平均差值(即平均每个式(1)两边的残差)仍仅为0.02~0.03,甚至更小(表1)[5];我们还使用鲨鱼的物种分歧时间成功地解析出一些寒武纪[6]和晚前寒武纪[7]动物门类的物种分歧时间,结果与化石记录相符,这些结果都说明Kimura提供的脊椎动物物种分歧时间是可靠的,因此不难理解我们分别将针鼹(单孔类哺乳动物)、袋鼠(有袋类哺乳动物)和狗(胎盘类哺乳动物)的物种分歧时间分别确定为2.32亿年前、1.39亿年前和8.1千万年前.
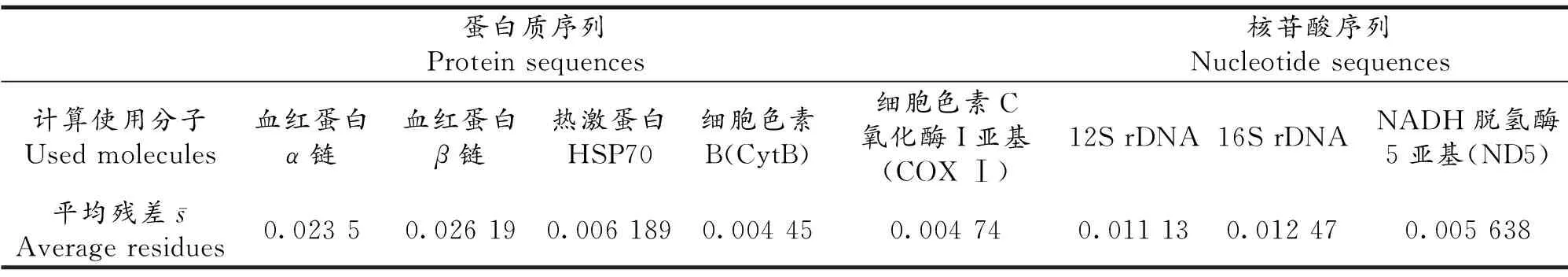
表1 前7种脊椎动物种间差异率拟合结果的平均残差Tab 1Simulation average residues of first 7 vertebrates
我们使用固定端点法来确定一些主要哺乳动物的物种分歧时间和演化关系.两个端点分别选取鲨鱼和狗(犬科动物),它们的物种名及其美国生物技术信息中心(NCBI)Genbank数据库分子序列号如表2.表2中各鲨鱼物种绝对进化速率来自文献[7]的计算结果,而犬科动物各物种绝对进化速率由下列目标函数使用最优化方法计算出:
(2)
其中,kA(i),kB(j)分别为第i种鲨鱼物种和第j种犬科动物的绝对进化速率;pAB(i,j)为它们之间的种间差异率;m,n分别为鲨鱼和犬科动物物种数.(2)式中鲨鱼和犬科动物物种分歧时间t2和t1的数值分别为0.468和0.081.由于线粒体COXⅡ分子绝对进化速率数值大多相对较大,需对(2)式进行多重突变校正,(2)式中2个自然指数项的校正公式分别为0.1+0.9e-kA(i)t2和0.1+0.9e-(kB(j)t1+k(t2-t1)),其具体原理参见文献[7],此时公式(2)转化为
(3)
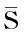
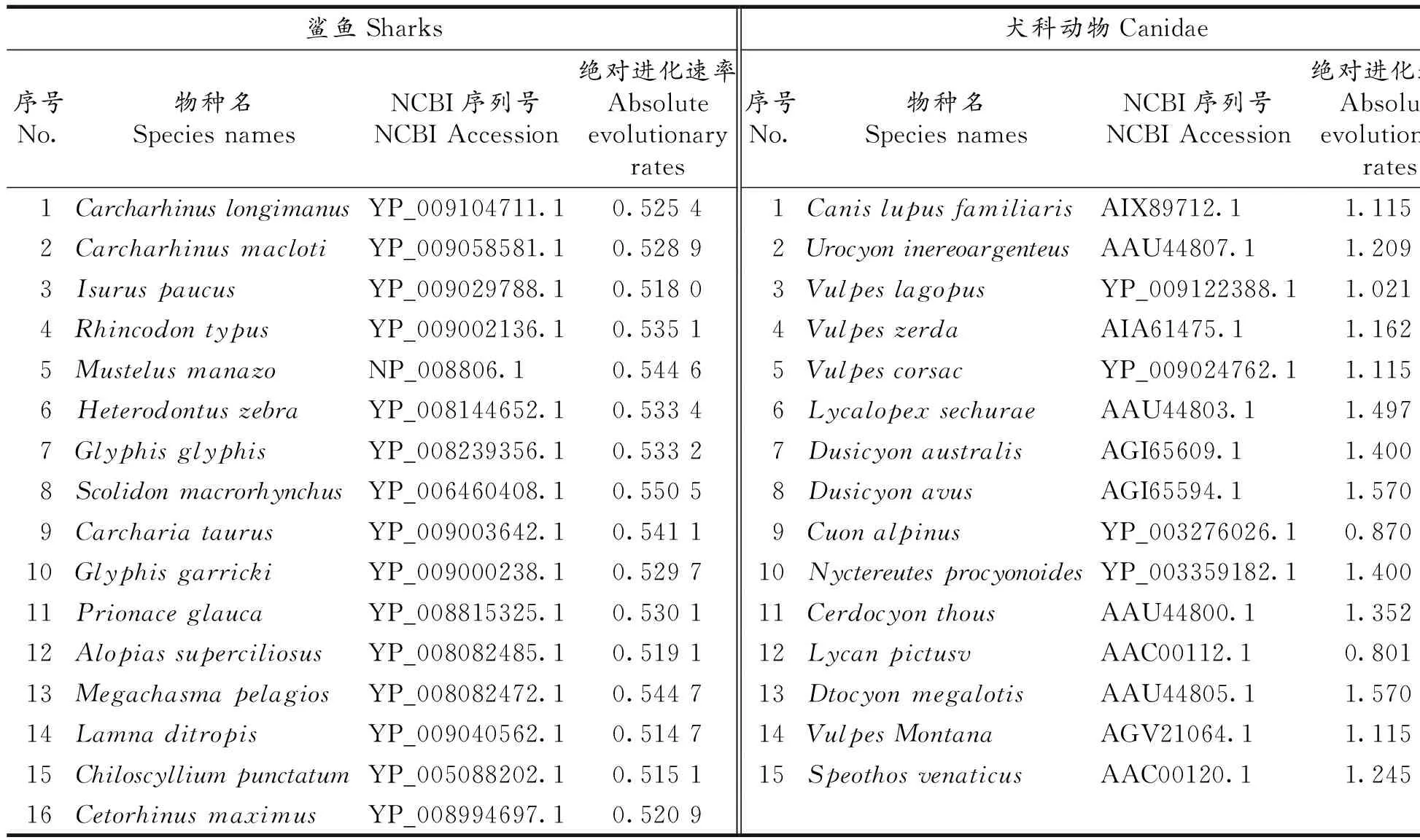
表2 计算所用鲨鱼和犬科动物COXⅡ蛋白质分子及其绝对进化速率Tab 2The species name and molecular evolutionary rates of sharks and canidaes used in the calculation
1.3 固定端点法确定一些主要哺乳动物物种分歧时间及演化关系
1.3.1 固定端点法 固定端点法的计算原理用图2来表示.
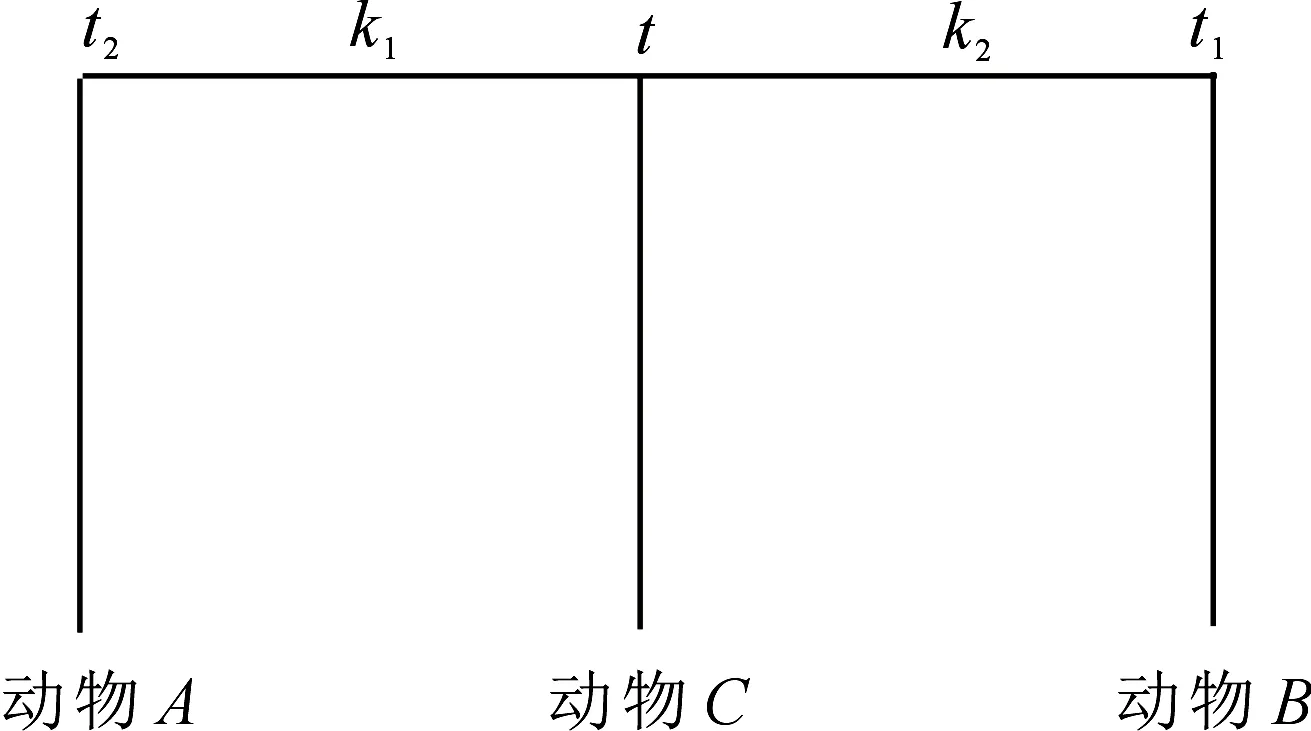
图2 固定端点法示意图Fig 2The scheme of fixed endpoints method
图2中动物A和动物B分别为前述的鲨鱼和犬科动物物种,动物C为需确定分歧时间的哺乳动物.将利用动物C分别与动物A和动物B两两比较得到的种间差异率PCA(i,j)和PCB(i,l),鲨鱼和犬科动物物种分歧时间及前面得到的它们的绝对进化速率等数据代入下列最优化目标函数中:
(4)
(4)式中t为待确定哺乳动物C的物种分歧时间,k1和k2分别为图2中主干枝t2至t段以及t至t1段的平均绝对进化速率.同样由于线粒体COXⅡ蛋白质分子绝对进化速率数值大多相对较大,对式(4)进行多重突变校正后有
(5)
1.3.2 一些有袋类哺乳动物的物种分歧时间 表3列出了一些有袋类哺乳动物(袋鼩等5种)使用(5)式得到的物种分歧时间等结果,种间差异率同样地采用NCBI网站提供的Protein blast软件进行对齐比较,并使用优化软件Lingo 11.0得到相应结果.
表3中参与计算的动物物种名及其NCBI序列号(括号内)分别如下:
袋鼩:1.Sminthopsispsamnophia(AIG23646.1);2.Sminthopsisdolichura(AIG23633.1);3.Sminthopsisdouglasi(YP_161185.1)
袋鼹:1.Notoryctescaurinus(AIG23321.1);2.Notoryctestyphlops(YP_161250.1)
负鼠:1.Caluromylanatus(AIG22970.1);2.Philanderfrenatus(AIG23503.1)
袋鼬:1.Parantechinusapicalis(AIG23360.1);2.Pseudantechinusmacrdonnellensis(AIG23555.1)
袋鼠:1.Macropusrefogriseus(AIG23188.1);2.Dorcopsulusvanheurni(AIG23087.1);3.Setonixbrachyurus(AIG23620.1);4.Dendrolagusdorianus(AIG23048.1);5.Lagorchestesconspicillatus(AIG23139.1)
由表3还可看出,前4种有袋类哺乳动物袋鼩、袋鼹、负鼠和袋鼬的物种分歧时间均在0.20左右,而袋鼠的物种分歧时间为0.139,这与文献[8]关于两个类群分属于哺乳动物有袋超目的多门齿亚目和双门齿亚目,且两个亚目于早期分化相符.同时文献还指出最早的有袋类哺乳动物化石出现于白垩纪中期的北美洲[8],而现有的分子生物学方法给出有袋类与胎盘类哺乳动物分歧时间为1.73~1.93之间,取决于使用的分子类型和计算方法[9],本文计算结果与现有分子生物学结果也大致相符.
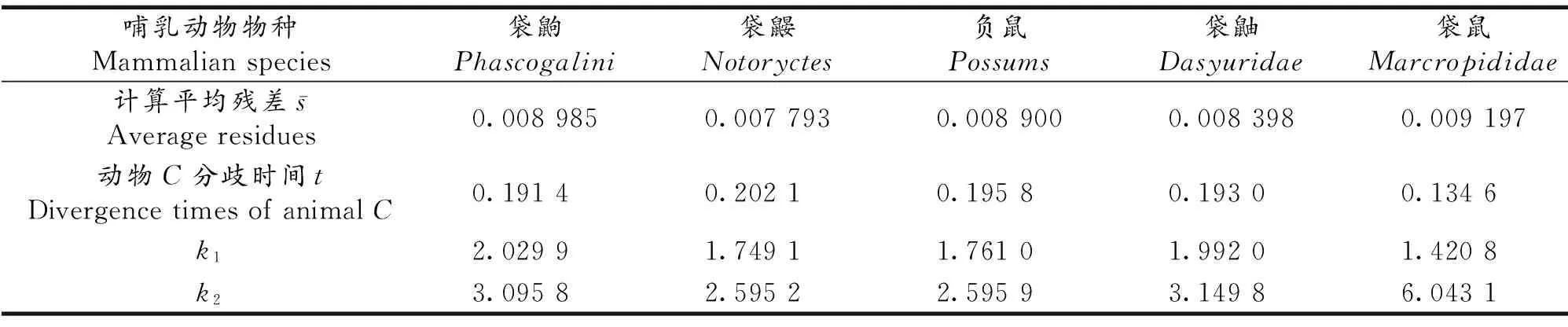
表3 一些有袋类哺乳动物的计算结果Tab 3The calculation results of some marpsupials
由图1和本文得到的3大类群单孔类、有袋类和胎盘类哺乳动物的物种分歧时间可以看出,它们的演化顺序为:单孔类→有袋类→胎盘类,即本文计算结果进一步支持哺乳动物进化的“兽类假说”.不过需要说明的是,表3中列出的最晚分异的有袋类哺乳动物为袋鼠,说明胎盘类哺乳动物可能自有袋类哺乳动物其中一个分枝进化而来,这与文献[10]认为有袋类与胎盘类哺乳动物具有共同祖先的结果存在差异.计算显示,袋鼠的物种分歧时间为1.39亿年前,若哺乳动物进化的“兽类假说”关于胎盘类哺乳动物为单系类群的假说正确的话,则胎盘类哺乳动物最早起源时间不应早于1.39亿年前,即袋鼠的物种分歧时间.这一结论与文献[11-12]认为胎盘类哺乳动物起源时间在1.25~1.30亿年前相符.而图1中我们得到的单孔类哺乳动物(针鼹)与有袋类和胎盘类哺乳动物分歧时间为2.32亿年前也与其他文献结果2.25亿年前相符[13].
2 人与其他人猿总科成员物种分歧时间的确定
人与其他人猿总科成员(黑猩猩、大猩猩、猩猩和长臂猿)的系统发育关系争论由来已久.由于缺乏明确的早期黑猩猩化石记录,人与其最亲缘的物种之间的演化关系一般主要依据分子生物学和形态学来研究.而目前古生物学家一般认为现有“分子钟”理论存在缺陷,未能考虑分子在不同谱系中进化速率的差异,因此其结果不可靠[14].为此,我们利用生物分子绝对进化速率公式对这一问题进行探讨.
2.1 计算模型
由于比犬科动物更进化的物种类群目前还没有非常准确的物种分歧时间,因此无法直接使用图2所示的固定端点法,对它进行改进,如图3.
图3中动物A有已知确切物种分歧时间t2,动物B和动物C为比动物A更进化的物种,它们物种分歧时间t1有待确定,不难理解求取t1时的最优化目标函数可表达为
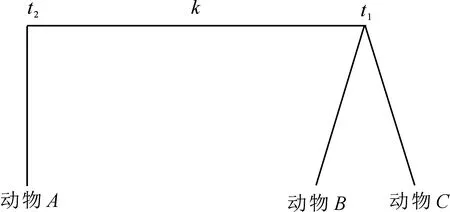
图3 改进后的固定端点法示意图Fig 3Scheme of modified fixed endpoint method
(6)
其中,kA(i),kB(j)和kC(l)分别为第i个动物A、第j个动物B和第l个动物C在它们各自进化分枝内绝对进化速率;pAB(i,j)和pBC(j,l)分别为动物A与动物B以及动物B与动物C两两比较得到的种间差异率.若考虑线粒体COXⅡ蛋白质分子的多重突变校正,则(6)式转化为
(7)
实际计算表明,自犬科动物分歧以后,进化主干枝进入一个快速进化期间,即图3中进化主干枝上t2至t1的遗传距离较各个进化分枝大很多.为了保证优化过程中(7)式中各变量在同一尺度范围内变化,且实际计算表明(7)式中第2个加和项的2个自然指数项的指数数值一般较小,我们将这2个自然指数项先分别用泰勒级数表达式展开(即进行线性化),再合并成同一个自然指数项,并让这一过程产生的误差并入残差,此时(7)式转化为
(8)
为了使计算模型稳定,将动物C的遗传距离kC(l)t1用一个新的变量c(l)替代.实际计算表明,选用类群内绝对进化速率存在一定差异的动物B和动物C物种有利于得到准确的结果.
2.2 人与旧世界猴和黑猩猩物种分歧时间
由于黑猩猩与犬科动物的遗传距离远大于其与人的遗传距离,为此我们首先求出旧世界猴的物种分歧时间.图3中动物A选用表2中的犬科动物,而动物B和动物C所选用动物物种名、NCBI序列号(括号内第一个数值)以及计算所得的动物B的绝对进化速率(括号内第二个数值)是:
动物B(旧世界猴):1.Macacafascicularis(AAA31889.1,2.157 3);2.CercopithecusDiana(YP_009024895.1,0.432 4);3.Rapiokindae(ACP43669.1,1.004 2);4.Rhinopithecusroxellana(AIX11520.1,0.568 7);5.Chlorocebuscynosuros(YP_009059462.1,1.885 3);6.Trachypithecuspilentus(YP_009047758.1,0.175 4);7.Pygathrixnigripes(YP_006460520.1,0.579 7);8.Cercocebustorquatus(ACP43668.1,1.992 0);9.Mandrillussphix(ACP43667.1,1.571 7);10.Erythocebuspatas(AHN49841.1,2.206 8);11.Theropithecusgelada(ACP43659.1,1.010 2);12.Lophocebusaterrimus(ACP43661.1,1.425 7);13.Miopithecusogouensis(AFA46428.1,1.271 9);14.Nasalislarvatus(YP_659450.1,0.750 4);15.Allenopithecusnigroviridis(YP_009024921.1,1.497 7);16.Semnopithecusentellus(YP_659437.1,0.678 8)
动物C(人猿总科):1.Homosapiens(ACM51349.1);2.HomoHeidelbergensis(YP_008963990.1);3.Pantroglodytesellioti(AIV00482.1);4.Pantroglodytestroglodytestroglodytes(AEO20839.1);5.Pantroglodytes(AEO20410.1);6.Gorillagorillagorilla(AHH93077.1);7.Gorillaberingeigraueri(AHH93064.1);8.Gorillagorilla(NP_008215.1);9.Pongoabelii(NP_007838.1);10.Panpymaeus(AAA61412.1)
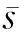
计算得到旧世界猴物种分歧时间为0.039 347 36,即3 934.736万年前,这与文献中大多数专家比较赞同猿类最早祖先是在大约3 000万年前从旧世界猴分离出去的说法基本相符[15].另外计算得到主干枝上t2至t1的绝对进化速率为15.738 6,人猿总科自t1的遗传距离在0.24~0.28之间,可以看出主干枝上该段遗传距离确实较进化分枝大很多.
计算还可得到人与黑猩猩的物种分歧时间为0.045 361 21,即453.61万年前,这与文献[16-17]一般认为人亚科最早物种为南方古猿,其化石年龄为440~460万年前相符.另得到主干枝上t2至t1的绝对进化速率为0.704 3氨基酸位点/亿年.
需说明的是,为了计算和表达的方便,从本处起除长臂猿外分子绝对进化速率单位改为氨基酸位点/亿年,物种分歧时间单位改为亿年.
2.3 人与大猩猩、猩猩和长臂猿的物种分歧时间
由于已获得人与黑猩猩的物种分歧时间,因此可以使用图2所示的固定端点法来分别确定人与猩猩、大猩猩和长臂猿的物种分歧时间.计算过程中将旧世界猴作为动物A,现代智人和海德堡人作为动物B,待定物种作为动物C,将旧世界猴以及现代智人和海德堡人的分子绝对进化速率、旧世界猴以及人与黑猩猩的物种分歧时间、序列分子两两比较得到的种间差异率代入(5)式,再使用优化软件Lingo11.0进行计算.为了便于计算,还使用约束条件t<0.393 413 1和t>0.045 361 21.计算结果见表6,表中长臂猿相关单位已修改,以便与其他人猿总科一致.
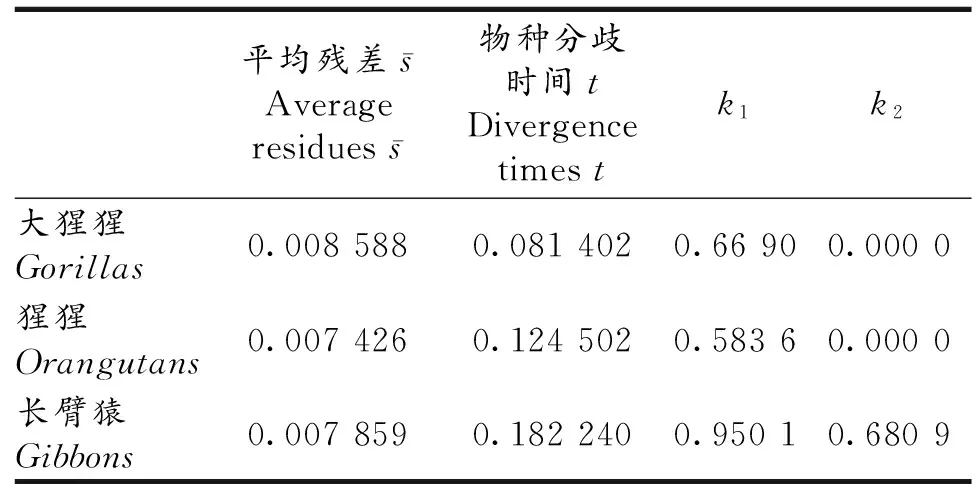
表6 人与一些人猿总科成员的物种分歧时间Tab 6 The divergence times of human and some other hominodaes
由表6可以看出,人与大猩猩、猩猩和长臂猿的物种分歧时间分别为814.02万年前、1 245.02万年前和1 822.40万年前,这与文献一般认为人猿总科成员演化路径为长臂猿→猩猩→大猩猩→黑猩猩→人相符,也与猩猩等物种分歧时间早于1 000万年前[17]等结果相符;同时,表6结果与分子生物学方法得到的长臂猿、猩猩、大猩猩和黑猩猩与人的物种分歧时间分别为1 800万年、1 400万年、700万年和300~500万年前的结果一致,它们也与目前基于化石记录得到的长臂猿与其他人猿总科动物的分异时间约在1 400~1 500万年前、猩猩的分异时间在1 200~1 300万年前、大猩猩的分异时间在800~900万年前、黑猩猩的分异时间在600~700万年前的结果相符.
由计算结果可以看出,除旧世界猴外,使用本文提出的确定哺乳动物分歧时间的方法得到的结果与化石记录或其他文献结果相符,说明本文使用的方法可行.另外需要说明的是计算得到的旧世界猴的分歧时间较现有文献结果略早,这可能与其计算时图3中时间t2至t1段和时间t1后人猿总科的遗传距离相差悬殊而现有分子生物学方法大多假定进化速率恒定有关,使用该结果得到的一些人猿总科成员的物种分歧时间与基于化石和分子生物学给出的结果相近进一步说明该结果是可信的.
[1] 周旭明,杨光.哺乳动物系统发育基因组学的研究进展[J].兽类学报,2010,30(3):339.
[2] 钟金城,吴玺.哺乳动物线粒体基因组的聚类分析及其系统进化研究[C]//中国遗传学会第八次代表大会暨学术讨论会摘要汇编(2004—2008).北京:中国遗传学会,2008.
[3] 李可群.分子绝对进化速率计算公式的推导及其应用方法[J].高师理科学刊,2015,35(1):19.
[4] KIMURA M.TheNeutralTheoryofMolecularEvolution[M].Cambridge:Cambridge University Press,1983.
[5] 李可群.分子绝对进化速率与物种分歧时间之间的定量关系[J].生物学杂志,2015,32(2):70.
[6] 李可群.COX I 蛋白质分子确定“寒武纪生物大爆发”时期物种分歧时间[J].生物学杂志,2016,33(1):62.
[7] 李可群.线粒体蛋白质确定晚前寒武纪后生动物的物种分歧时间及其早期演化的探索[Z].作者已写就论文.
[8] 王宗祎.有袋类[J].生物学通报,1986,21(4):7.
[9] RHEEDE T V,BASTIAANS T,BOONE D N,et al.The platypus is in its place:nuclear genes and indels confirm the sister group relationship of monotremes and therians[J].MolecularBiologyandEvolution,2006,23(3):587.
[10] 孙跃武,刘鹏举.古生物学导论[M].北京:地质出版社,2006.
[11] WEIL A.Mammalian evolution:upwards and onwards[J].Nature,2002,416:798.
[12] JANKE A,FELDMAIER-FUCHS G,THOMAS W K,et al.The marsupial mitochondrial genome and the evolution of placental mammals[J].Genetics,1994,137:243.
[13] BRAWAND D,WAHIL W,KAESSMANN H.Loss of egg Yolk gene in mammals and the origin of lactatim and placentation[J].PlosBiology,2008,6(3):e63.
[14] 高虹.人类起源问题及其两次大争论[J].生物学通报,1997,32(7):24.
[15] 夏志.人猿情未了[N].北京科技报,2015-03-16(52).
[16] HORAI S,SATTA Y,HAYASAKA K,et al.Man’s place in Hominoidea revealed by mitochondrial DNA genealogy[J].JournalofMolecularEvolution,1992,36:32.
[17] 李辉.遗传学对人科谱系的重构[J].科学,2013,66(2):7.
(责任编辑 俞诗源)
Resolving divergence times and phylogenetic relationship of mammals by COXⅡ protein molecules
LI Ke-qun
(Department of Chemistry,Tongji University,Shanghai 200092,China )
The divergence times of some major mammalian groups are obtained using the calculation equation for biological molecular evolutionary rates presented by the author and COXⅡ protein molecules. These results show thatMonotrememammals,such asechidnaseparated from the stem line towards primates about 232 myrs ago,this divergence time agrees with their fossil records.Marsupialmammals,such asPhascogalini,Notoryctes,PossumsandDasyuridaeall separated from the stem line towards primates around 200 myrs ago,whileKangaroosseparated around 139 myrs ago,these divergence times agree with the theory thatPhascogalini,Notoryctes,PossumsandDasyuridaebelong to polyprotodontia butKangaroosbelong to diprodontia,and two groups separated at early age.Our results support the “Theria hypothesis”,that is,mammals evolved from monotremes to marsupials and then to placentals.If the “monophyletic group hypothesis” of placentals is correct,the starting times of all placentals should not be earlier than 139 myrs ago,which is the divergence time of kangaroos.The divergence times of human with some primates,such asChimpanzees,Gorillas,Orangutans,Gibbonsand old world monkeys are also obtained by our calculation.They are 4.536 1 myrs ago,8.140 2 myrs ago,12.450 2 myrs ago,18.224 0 myrs ago and 39.347 4 myrs ago respectively,these results agree with other reference works or fossil records exceptChimpanzeeswith no early fossils found yet.Key words:mammal;primate;divergence time;molecular clock;evolutionary rate;COXⅡ protein molecules
10.16783/j.cnki.nwnuz.2016.01.017
2015-07-26;修改稿收到日期:2015-09-17
李可群(1968—),男,湖南长沙人,讲师,硕士.主要研究方向为分子遗传进化.
E-mail:likequn@tongji.edu.cn
Q 951
A
1001-988Ⅹ(2016)01-0079-07
猜你喜欢
中国生殖健康(2020年5期)2021-01-18 02:59:52
环球时报(2019-11-28)2019-11-28 04:14:34
中外文摘(2018年16期)2018-11-21 02:54:51
中国生殖健康(2018年5期)2018-11-06 07:15:42
小学科学(学生版)(2018年5期)2018-06-15 08:48:00
奇妙博物馆(2018年12期)2018-04-25 02:16:04
奇妙博物馆(2018年12期)2018-04-25 02:16:02
兽医导刊(2017年4期)2017-04-04 14:00:26
高中生·天天向上(2016年11期)2017-02-28 07:40:18
山西卫生健康职业学院学报(2014年5期)2014-04-09 06:36:50
