
亚热带分层水库固氮微生物时空分布格局
2016-11-28 08:14:02王丽娜陈辉煌刘乐冕
生态学报 2016年18期
王丽娜,陈辉煌,刘乐冕,余 正,杨 军,*
1 中国科学院城市环境研究所,城市环境与健康重点实验室,水生态健康研究组, 厦门 361021 2 中国科学院大学, 北京 100049
亚热带分层水库固氮微生物时空分布格局
王丽娜1, 2,陈辉煌1,刘乐冕1,余 正1,杨 军1,*
1 中国科学院城市环境研究所,城市环境与健康重点实验室,水生态健康研究组, 厦门 361021 2 中国科学院大学, 北京 100049
生物固氮作用是水生态系统氮元素的重要来源途径之一,通常通过固氮微生物实现。但是,目前人们对亚热带分层水库固氮微生物多样性、分布和丰度认识还非常有限。以厦门市汀溪水库为例,基于固氮基因(nifH)综合应用克隆文库、定量PCR、定量RT-PCR研究固氮微生物在不同季节和不同水层的时空分布格局与演替规律。结果表明,汀溪水库具有丰富多样的固氮微生物,包括蓝藻、α-变形菌、β-变形菌、γ-变形菌、厚壁菌,以及少量未知的固氮细菌和序列;固氮微生物的群落组成、丰度、多样性和活性均呈现显著的时空差异。春、夏和秋3季表层和底层蓝藻nifH基因序列所占比例均超过50%,其中表层高于底层;冬季表层和底层蓝藻OTU数目比例超过50%。聚类分析表明,冬季表层和底层群落汇聚为一类;春、夏和秋三个季节表层首先聚为一类,然后与底层分别汇为一支。汀溪水库热分层时期的固氮微生物群落组成的空间差异大于季节差异,而且表层水体蓝藻在所有固氮微生物中占据绝对优势地位。相关分析表明,固氮微生物RNA丰度和RNA/DNA分别与氨氮、水温显著负相关;固氮微生物DNA丰度与溶解氧、pH、叶绿素a显著负相关,与硝氮显著正相关。综上所述,亚热带水库热分层对固氮微生物的群落结构具有显著的影响,在水库环境保护和生态管理中,特别是蓝藻水华防控时,要充分考虑水体热分层的生态效应。
生物固氮;nifH基因;分层水库;汀溪水库;时空变异

在淡水生态系统中,生物固氮受到氮磷浓度与光照强度的影响[19],然而不同固氮微生物多样性和活性的时空分布规律还不是很明确[13]。Short[20]等人在切萨皮克湾生态系统中发现两种nifH基因型有着不同的季节和空间分布格局,暗示这些生物群落受到不同环境过程的影响。Moisander[9]等人研究切萨皮克湾水柱和沉积物固氮微生物组成的时空变化,发现多样性最高的是nifHCluster 3,但是Cluster 1(α-,β-,γ-变形菌,蓝藻)具有较高的丰度;底层水体固氮微生物群落具有较高的相似性,而表层水体群落变异较大;盐度、溶解性无机氮、溶解性有机碳和溶解性有机磷等环境因子与固氮微生物群落组成有显著的关系。Yu[12]等人研究东圳水库中固氮微生物秋季垂直分布特征,发现温跃层的出现使水温与溶解氧发生明显垂直变化,从而对固氮微生物群落组成造成了显著影响。以上研究报道显示,水生固氮微生物的垂直分布和时空动态过程的相关研究是最新发展趋势和研究热点。
我国亚热带深水水库多属于典型的暖单次混合型水库,除冬季有短暂的混合期外,在一年当中会持续出现热分层现象[21]。在这些水库中,明显的热分层是春季、夏季与秋季的典型特征,在温跃层上下水体中明显差异的环境对于微生物群落的组成、多样性及丰度可能有着显著的影响[22]。显然,亚热带分层水库同样非常适合研究固氮微生物垂直分布和季节动态变化规律。本研究以厦门汀溪水库为例,综合应用克隆文库、定量PCR、定量RT-PCR技术分析固氮微生物在4个季节不同水层的组成、多样性、活性及丰度。本研究的目的是:1)揭示分层水库固氮微生物群落的季节和空间分布格局;2)探讨固氮微生物优势类群的特征与成因。
1 材料与方法
1.1 样品采集及理化分析
汀溪水库(24°48′N,118°08′E)位于福建省厦门市,是一座典型的亚热带深水水库。本研究选取汀溪水库湖泊区的深水区设置站位进行采样,此站位水深可达25 m。水体样品采集周期为1a:2012年的7月(夏季),10月(秋季),以及2013年的1月(冬季),4月(春季)。每次采样均采集5个水层的样品,分别是:0、6、12、18、24 m(夏),0、10、15、20、25 m(秋),0、5、10、15、21 m(冬),0、5、10、15、20 m(春)。每份样品均采集10 L水样,水样采集后立即送往实验室进行过滤,采用孔径为0.22 μm的滤膜(直径47 mm,Millipore,美国)过滤1 L水样,并保留4份重复样品。将载有微生物的滤膜储存在-80℃的冰箱,用于后续DNA和RNA的提取。
现场利用美国哈希HACH多参数水质分析仪(Hydrolab DS5)监测水温、溶解氧、pH、叶绿素a等参数,每隔1 m测定1次。水体营养盐分析在实验室进行,总氮、氨氮、硝态氮、总磷、正磷酸盐均按照国家标准进行测定。
1.2 DNA和RNA提取
根据说明书,利用试剂盒Fast DNA spin kit(MP Biomedicals, 美国)和E.Z.N.A. total RNA kit(Omega Bio-Tek, 美国)分别提取固氮微生物DNA和RNA。提取的RNA立即反转录成cDNA,使用的是试剂盒OneStep RT-PCR kit Version 2.0(Takara,日本)。反转录程序是:37 ℃,15 min; 95 ℃,5 s。提纯后的DNA和cDNA均置于冰箱-40 ℃保存备用。
1.3 PCR扩增及克隆文库分析
本研究扩增nifH基因选择的引物是nifH-F (5′-CGTAGGTTGCGACCCTAAGGCTGA-3′) 和nifH-R (5′-GCATACATCGCCATCATTTCACC-3′)[23]。PCR的扩增体系是50 μL,包含:5 μL 10 × PCR缓冲液(10 nmol/L),4 μL dNTPs(10 mmol/L),0.5 μLTaq聚合酶(5 U/μL),前后引物各1 μL(10 μmol/L),100 ng DNA模板,然后使用ddH2O补充到50 μL。PCR反应体系如下:94 ℃预变性5 min,然后40个循环(94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸40 s),最终72 ℃延伸10 min。PCR产物利用1%的琼脂糖凝胶进行电泳检测。利用胶回收试剂盒提纯PCR产物,然后根据操作指南将其连接到pMD18-T载体(Takara, 日本)。将连接后的载体转移到大肠杆菌DH5α(Takara, 日本)中,置于液体培养基中37 ℃培养过夜。利用添加氨苄的固体培养基进行蓝白斑筛选,挑取白色克隆进行测序分析。本文选用四个季节的表层和底层样品,共构建8个克隆文库,每个克隆文库挑选70个克隆子进行测序。
1.4 定量PCR
定量PCR与克隆文库所用的引物是相同的。利用Applied Biosystems 7500 Real-Time PCR System(ABI,美国)进行定量PCR,每个样品设置3个重复[12]。本研究采用的是20 μL体系:0.5 μL前后引物(10 μmol/L),10 μL SYBR Premix Ex TaqTM,2 μL DNA 或cDNA模板和7 μL RNase-free water。本研究的标准曲线的R2值为0.994(DNA)和0.991(RNA),扩增效率分别是97.45%(DNA)和98.19%(RNA)。RNA/DNA比值可以作为反映固氮微生物基因表达活性的参数,因此也进行了分析。
1.5 数据分析
克隆文库的操作分类单元(Operational taxonomic unit,OTU)划分采用97%的相似度,利用软件MOTHUR v.1.33.0分析8个文库的稀释曲线,然后计算OTU个数、覆盖率、Shannon-Wiener指数、Simpson倒数指数、Pielou均匀度指数、以及反映OTU丰富度的Chao1指数和 ACE指数[24]。将测序获得的序列在GenBank数据库中进行比对,查找最相似序列并确定其分类归属,统计分析主要类群的OTUs比例和序列比例。利用PRIMER 6.0计算样品间的Bray-Curtis相似系数,进而对样品进行聚类分析[25]。利用SPSS 19.0进行单因素方差方法,确定混合期及分层水体中不同水层之间水体理化及生物指标的差异是否显著(P< 0.05),应用Spearman相关分析确定DNA拷贝数、RNA拷贝数及RNA/DNA比值与环境因子之间的相关性是否显著(P< 0.05)。
2 结果
2.1 水库环境因子的时空变化

图1 汀溪水1周年内的水温、溶解氧、pH和叶绿素a垂直变化Fig.1 Changes in water temperature, dissolved oxygen (DO), pH and chlorophyll a in the water column of the Tingxi Reservoir during one year水库底层用灰色表示,黑色圆点表示固氮微生物采样点
汀溪水库是一座典型的亚热带分层水库,在冬季呈现短时期的上下混合,在春季、夏季和秋季出现稳定的热分层(图1)。本研究中,春季4月份、夏季7月份、秋季10月份水库的温跃层分别出现在3—14 m、18—22 m、21—24 m。冬季1月份处于上下混合的状态,水体物理化学参数从表层到底层无明显差异。春季分层开始出现时,温跃层跨度较大,为11 m。夏季和秋季的温跃层厚度相对狭窄(厚3—4 m)。显然,随着热分层状态的变化,水体物理化学参数也呈现梯度变化,使得不同季节同一水层或同一季节不同水层的生境特征差异显著(表1)。通常,分层时期的水温、pH、溶解氧、叶绿素a在湖上层均显著高于湖下层(P< 0.05)。营养盐在不同水层差异也比较显著,湖上层总氮、氨氮、总磷、正磷酸盐均低于湖下层。

表1 汀溪水库水体理化参数和固氮微生物群落特征
不同小写字母表示显著性差异(P<0.05),数据为平均值±标准误
2.2 固氮微生物的丰度和活性

图2 汀溪水库4个季节不同水层的nifH DNA与RNA拷贝数及RNA/DNA比值Fig.2 Number of nifH DNA and RNA copies per liter and RNA/DNA ratio from different water depths and different seasons in the Tingxi Reservoir误差线表示3个重复样品的标准误
固氮微生物的丰度和活性随着季节和水层的变化发生明显的变化(图2)。冬季固氮微生物的nifHDNA丰度随着深度的增加有一个缓慢上升的趋势,在底层达到最大值4.52×107拷贝/L;春季固氮微生物nifHDNA丰度在温跃层(5,10 m)要低于湖上层和湖下层,在湖下层15 m水深处为最大值4.95×107拷贝/L;夏季nifHDNA丰度在6—12 m急剧上升,在温跃层开始点18 m达最高值7.66×107拷贝/L,24 m稍微下降,但并不明显;秋季nifHDNA丰度随着深度的增加而逐渐减少,水深20 m水层丰度最低,有趣的是nifHDNA丰度最大值位于秋季表层。固氮微生物nifHRNA水平变化趋势与DNA存在一定的差异性,而且RNA丰度显著低于DNA丰度,相差2个数量级。冬季水深10 m和15 m处的nifHRNA丰度要明显高于其他水层;春季随水深增加先降低后上升,在10 m处为最低值;夏季,随着深度的增加呈现明显上升的趋势,在底层24 m达到最高值;秋季呈现总体下降的趋势,在15—20 m急剧下降,与nifHDNA变化趋势一致。RNA/DNA是一个能够表征基因表达活性的指标,而4个季节表现出不同的垂直分布格局。例如,4个季节活性最高出现的位置不同:10 m(冬季)、20 m(春季)、24 m(夏季)、15 m(秋季)。方差分析显示,nifHDNA和RNA在不同水层之间差异不显著,而RNA/DNA在温跃层显著低于混合期水层(表1)。显然,固氮微生物群落的DNA、RNA丰度变化趋势呈现出一定差异性;基因表达活性的显著特征是在春季和夏季的底层活性最高,而在秋季湖上层的活性显著高于湖下层。据此推测,温跃层的出现是造成湖上层与湖下层差异的主要原因。
2.3 固氮微生物的多样性
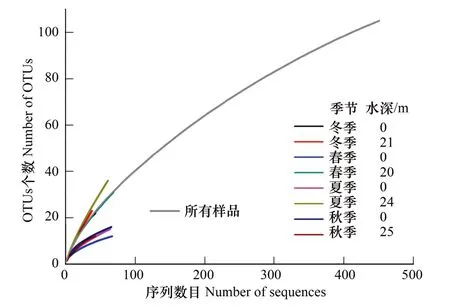
图3 汀溪水库固氮微生物nifH基因克隆文库稀释曲线(OTUs在97%相似性水平划分)Fig.3 Rarefaction curves of nifH gene sequences, which were grouped into OTUs based on 97% sequence similarity level
4个季节的表层和底层8个样品克隆文库共成功获得451条序列,尽管稀释曲线没有平缓,在97%相似度水平共鉴定106个固氮微生物OTUs(图3)。主要类群包括:蓝藻(Cyanobacteria)46个OTUs、α-变形菌(Alphaproteobacteria)21个OTUs、β-变形菌(Betaproteobacteria)3个OTUs、γ-变形菌(Gammaproteobacteria)9个OTUs、厚壁菌(Firmicutes)1个OTU,以及其他未知的固氮细菌18个OTUs、未知序列8个OTUs(表S1)。其中98个OTUs的序列与GenBank中各种生态坏境中的固氮微生物序列相似,相似度水平范围是80%—99%。然而,在50%相似性水平下,20条序列(8个OTUs)在数据库中没有找到相似的序列,因此属于未知序列。8个克隆文库中,冬季混合期水库表层和底层OTUs数目并没有差异,然而分层期水库表层和底层显示出明显的差异。例如,春季和夏季底层OTUs数目高于表层,而秋季表层OTUs数目稍高于底层(表2)。在本研究测序范围内,固氮微生物群落的多样性指数,包括Shannon-Wiener指数、Simpson倒数指数、Pielou均匀度指数均呈现一定时空变化,变化趋势与OTUs相似(表2)。根据稀释曲线计算的OTU丰富度指数Chao1 和 ACE 均表现出分层差异,冬、春、夏3个季节水库底层大于表层,而秋季却呈现为表层高于底层的特征(表2)。
2.4 固氮微生物的群落组成
除秋季底层蓝藻OTUs的比例为16.67%外,在所有样品中蓝藻物种所占比例是最大的一类,OTUs占据38.89%—56.52%,序列占据34.21%—76.92%(图4)。α-变形菌OTUs为8.33%—27.78%,其中秋季表层多样性最高,序列丰度为2.33—10.29%;β-变形菌在冬季底层、春季表层、夏季表层,以及秋季表层和底层均没有检测到,而且OTUs和序列所占比例均小于5%;γ-变形菌在4个季节的表层与底层都被检测到,OTUs为4.00%—18.75%,序列为2.70%—13.24%;厚壁菌仅仅在秋季表层被检测到,OTUs为5.55%,序列为1.52%;未知固氮菌在4个季节中均被发现,而且底层OTUs和序列数均高于表层;未知序列在夏季表层和秋季表层之外其他水层都有检测到,而且底层均高于表层(图4)。
表2 汀溪水库4个季节表层与底层固氮微生物nifH基因的多样性
Table 2 Diversity and predicted richness of thenifHgene sequences from both surface and bottom waters of the Tingxi Reservoir in four different seasons

季节Season水深/mDepth序列数SequencesnumberOTU0.03个数OTU0.03number覆盖率/%Coverage香农威纳指数Shannon-WienerindexSimpson倒数指数SimpsoninverseindexPielou均匀度指数PielouevennessChao1指数Chao1indexACE指数ACEindex冬季Winter0442368.182.8718.50.633869冬季21372548.652.7612.80.6180491春季Spring0671291.041.573.030.441736春季20693368.122.9614.70.60108182夏季Summer0641685.941.945.000.502795夏季24613654.103.2221.30.6299445秋季Autumn0661787.882.196.670.552540秋季25431288.371.703.130.471416

图4 汀溪水库固氮微生物主要类群OTUs和序列所占比例Fig.4 Proportions of the main diazotroph groups detected in the Tingxi Researvor. The proportions are computed in terms of OTUs and sequences
聚类分析结果显示,4个季节表层与底层的固氮微生物群落组成明显不同(图5)。冬季表层与底层的群落组成比较相似,差异相对较小。存在分层现象的春、夏、秋3个季节,表层固氮微生物群落组成最相似;春、夏底层的群落组成较相似,秋季底层显示出其独特性。据此推测,水体分层对固氮微生物群落组成的影响比季节差异的影响更明显。

图5 汀溪水库固氮微生物的聚类分析展示群落组成的时空分布格局Fig.5 Cluster dendrogram showing seasonal and spatial patterns of diazotrophic bacteria community from the Tingxi Reservoir based on clone library data
2.5 固氮微生物群落与环境因子的关系
汀溪水库nifHDNA丰度与pH、溶解氧、叶绿素a显著负相关,与硝态氮显著正相关;nifHRNA丰度则与氨氮存在极显著的负相关关系;RNA/DNA与水温显著负相关(表3)。显然,热分层发生时温跃层的出现及其导致的水体理化分层与固氮微生物群落组成、丰度和活性密切相关。
3 讨论
3.1 固氮微生物群落的时空变化
汀溪水库在水体分层期间(春、夏、秋),固氮微生物群落在上下水层空间上的差异要明显高于季节上的差异。在冬季处于混合期,水体上下混合导致表层到底层物理化学因子比较接近,同样表层与底层的固氮微生物群落组成也比较接近,然而nifHDNA和RNA在底层的丰度却高于表层。利用nifH基因可以检测到广泛的自养和异养固氮微生物:蓝藻、古菌、厚壁菌、螺旋菌、α-变形菌、β-变形菌、γ-变形菌和δ-变形菌[1]。本研究同样检测到蓝藻、α-变形菌、β-变形菌、γ-变形菌、厚壁菌、未知固氮菌,还有少量不能确定归属的新序列。在所有的已知类群中,蓝藻序列占据最优势的地位。水库在春季开始出现分层,固氮微生物群落组成在表层和底层就呈现出明显的差异,例如春季底层OTU数目,以及多样性指数均明显高于表层,而且表层没有检测到β-变形菌。夏季相较于春季而言温跃层下降,有趣的是夏季底层检测到的nifHDNA丰度、RNA丰度、RNA/DNA、OTU数目都明显高于表层,说明底层固氮微生物丰度、活性、多样性高于表层。秋季温跃层位置继续下降,表层OTU数目高于底层;而且表层nifHDNA丰度、RNA丰度、RNA/DNA明显高于底层,呈现出与夏季相反的垂直分布格局。Moisander[9]等人发现在4月、10月切萨皮克湾的底层水体固氮微生物群落组成比较相似,而表层样品变化更加明显,这与本文的结果存在差异。汀溪水库4月、7月和10月表层水体的固氮微生物组成更加相似,而底层样品多样性高,群落组成变化较大。有趣的是,在秋季表层检测到厚壁菌(Firmicutes)。厚壁菌微生物一般大量出现在土壤环境[26],本文的检测结果暗示具有固氮作用的厚壁菌也能生存在水体环境或者来源于库区流域的土壤。由于在春夏表层没有发现β-变形菌,在底层却检测到,因此认为具有固氮功能的β-变形菌在分层水库中可能更加偏好底层的低温、缺氧的环境。
表3 固氮微生物nifHDNA、RNA、RNA/DNA与环境因子之间的Spearman相关系数
Table 3 The Spearman correlation coefficients betweennifHDNA, RNA, RNA/DNA ratio, and environmental variables in the Tingxi Reservoir

相关性CorrelationDNARNARNA/DNA相关性CorrelationDNARNARNA/DNA水温WatertemperatureNSNS-0.320∗pH-0.433∗∗NSNS溶解氧Dissolvedoxygen-0.316∗NSNS叶绿素aChlorophylla-0.315∗NSNS总氮TotalnitrogenNSNSNS氨氮NH4-NNS-0.362∗∗NS硝态氮NOx-N0.382∗NSNS总磷TotalphosphorusNSNSNS正磷酸盐PO4-PNSNSNS
*P< 0.05; **P< 0.01; NS:不显著 not significant
随着大量相关研究的进行,固氮微生物的多样性和复杂性逐步被人们所认知[1],但是影响水生态系统固氮微生物的分布及多样性的因子还不是很清楚[20]。在贫营养海洋中,固氮微生物的丰度低于湖泊或者近海岸水体,但是多样性却更高[1]。固氮蓝藻是湖泊和水库中常见的水华种类[27],而且通常占据优势地位,如纽约乔治湖[28]。有趣的是,氮是莫诺湖的限制因子,尽管在垂直水柱中没有检测到固氮作用的发生,表层和底层水体却发现多样性很高且群落差异显著的固氮微生物[1]。河口具有多样的固氮微生物,如切萨皮克湾与纽斯河具有丰富的α-变形菌、β-变形菌、γ-变形菌[1, 29]。同样,在河口沉积物和沼泽中,固氮微生物是非常多样的,而且不论氮含量高低或者环境变化,总是会保持相对稳定的状态[30-31]。综上所述,固氮微生物种类在水环境中具有广泛的分布,包括微生物垫、湖泊、盐沼、河口等多种生境,而在不同环境下存在着各自的分布特点[20]。本文的研究结果在一定程度和前人的结果一致,例如,蓝藻是淡水生境优势类群;但也存在一定的特殊性,比如检测到厚壁菌。值得注意的是,本研究中8个OTUs在GenBank上没有找到比较接近的序列,暗示这些OTUs可能是新的未知序列,迫切需要后续的实验分析确定其分类地位信息。此外,这些未知固氮菌和未知序列通常在4个季节底层的出现频率高于表层,主要原因可能是过去对湖泊水库的研究往往忽视底层水体,对于底层水体微生物的研究远远少于表层。
3.2 固氮蓝藻占优势的原因
在汀溪水库的8个克隆文库中,固氮微生物的多样性非常高,更重要的特征是蓝藻占据绝对优势。湖泊水库蓝藻优势的存在有其物理、化学、生物因素等多重原因[32]。研究表明,水库水体上下混合运动会影响物种间的相互作用,在小范围内存在水体运动的环境中,有竞争作用的物种也可以共同存在[33]。Zani[28]等人在乔治湖发现固氮蓝藻与其他固氮菌共同存在,这与本文在同一样品中检测到多样的固氮微生物是一致的,虽然有竞争作用,但是可以共同生存。通常,氧气对固氮酶活性具有抑制作用,蓝藻占据优势地位也与自身生理结构有关[1]。例如蓝藻有两种固氮机制:空间上区分开光合作用和固氮作用;时间上错开光合作用和固氮作用[34]。因此,蓝藻在水体表层和底层都可以进行固氮作用,而且氧气不会对其固氮作用造成不利影响。有研究报道,海洋蓝藻束毛藻(eg.Trichodesmiumspp.)是热带贫营养盐海域优势的固氮微生物[35],而具有异形胞的蓝藻在淡水和半咸水环境中都可以成为占优势的类群[36]。具有异形胞的蓝藻相对于非异形胞蓝藻更加适合固氮作用[35],因为异形胞可以空间上分开光合作用与固氮作用,这样不仅固氮效率更高,而且也有效避免了氧气的抑制作用,因此可以解释在氮磷比低的淡水湖泊和水库中固氮蓝藻容易形成水华[37-38]。变形菌门的固氮微生物多数是异养型固氮微生物,相对喜欢底层低氧或厌氧环境[39]。Toepel[40]等人研究了Cyanothecesp. ATCC 51142株在白天黑夜及持续性光照条件下固氮基因表达差异性,实验证明其在白天会储存糖原,连续光照能够诱导晚上进行固氮作用。另一方面,水库稳定的分层现象也为蓝藻提供了有利的条件,蓝藻可以在分层水体上下自由移动,这是因为蓝藻体内有特殊的气泡结构,可以通过自身调节提供浮力[41]。蓝藻可以借助气泡在水层中找到光、营养盐等合适生境的水层,进行生长繁殖[42]。由此可以解释,在亚热带深水水库蓝藻不论季节和空间上的变化都可以占据优势地位。
Langois[43]等人发现蓝藻更偏好表层水体,这与我们研究结果一致,因为汀溪水库4个季节表层蓝藻的丰度比例和物种多样性均高于底层。蓝藻喜欢表层并不是一个因素决定的,可能是蓝藻属于自养光合生物,偏好较高水温、更适宜光照的表层水体,或者是同其他生态因子的综合效应[27, 35]。Bentzon-Tilia[44]等人研究了两个河口湾的固氮蓝藻,发现两个河口湾表层水体比海洋束毛藻(Trichodesmium)水华爆发期的固氮效率还要高。由此推测,水库生态系统中蓝藻在固氮作用中扮演着很重要的角色。Voss[45]等人发现随着海水深度变化的总溶解铁与固氮作用是显著相关的。这需要后续的野外控制实验验证水库中微量元素溶解铁与固氮过程的关系,进而可以更加深入的了解蓝藻固氮的生态过程与机制。另外一种原因,可能是蓝藻体内有气泡、具有很强的漂浮能力,也更有利于在表层占据优势地位[41-43]。本文研究结果提示,在亚热带深水水库全年具有蓝藻水华爆发的条件和可能,因此在蓝藻水华预防和控制时要充分考虑水体分层的变化规律。
4 结论
本研究鉴定固氮微生物OTUs超过100个,表明汀溪水库中具有多样性很高的固氮微生物,主要类群包括蓝藻、α-变形菌、β-变形菌、γ-变形菌、厚壁菌,以及少量未知的固氮细菌和序列。固氮微生物的丰度、活性、多样性及群落组成具有明显的季节和垂直分布格局,在春夏秋水库热分层时期,群落组成在水体表层和底层的空间变化明显大于时间变化,暗示热分层对固氮微生物群落具有更为显著的影响。春夏秋冬4个季节里,蓝藻在水库中始终占据最优势地位,特别是在水库分层时期(春、夏、秋),蓝藻的优势更加突出。高温和热分层环境,以及蓝藻的独特生理结构有利于表层水体固氮蓝藻占优势。有趣的是,汀溪水库夏季和秋季固氮微生物丰度和活性呈现相反的垂直变化趋势。在后续研究中,有必要更加深入系统研究并揭示热分层条件下固氮微生物群落变化的生态机制,进而为预防和控制蓝藻水华发生提供对策。
[1] Zehr J P, Jenkins B D, Short S M, Steward G F. Nitrogenase gene diversity and microbial community structure: a cross-system comparison. Environmental Microbiology, 2003, 5(7): 539-554.
[2] Zehr J P, Paerl H W. Molecular ecological aspects of nitrogen fixation in the marine environment // Kirchman D L, ed. Microbial Ecology of the Oceans, Second Edition. New York: John Wiley & Sons, Inc., 2008: 481-525.
[3] Gaby J C, Buckley D H. A comprehensive evaluation of PCR primers to amplify thenifHgene of nitrogenase. PLoS One, 2012, 7(7): e42149.
[4] DeLong E F, Preston C M, Mincer T, Rich V, Hallam S J, Frigaard N-U, Martinez A, Sullivan M B, Edwards R, Brito B R, Chisholm S W, Karl D M. Community genomics among stratified microbial assemblages in the ocean′s interior. Science, 2006, 311(5760): 496-503.
[5] Short S M, Zehr J P. Nitrogenase gene expression in the Chesapeake Bay Estuary. Environmental Microbiology, 2007, 9(6): 1591-1596.
[6] Farnelid H, Öberg T, Riemann L. Identity and dynamics of putative N2-fixing picoplankton in the Baltic Sea proper suggest complex patterns of regulation. Environmental Microbiology Reports, 2009, 1(2): 145-154.
[7] Young J P W. Phylogenetic classification of nitrogen-fixing organisms // Stacey G, Evans H J, Burris R H, eds. Biological Nitrogen Fixation. New York, USA: Chapman and Hall, 1992: 43-86.
[8] Gaby J C, Buckley D H. A global census of nitrogenase diversity. Environmental Microbiology, 2011, 13(7): 1790-1799.
[9] Moisander P H, Morrison A E, Ward B B, Jenkins, B D, Zehr J P. Spatial-temporal variability in diazotroph assemblages in Chesapeake Bay using an oligonucleotidenifHmicroarray. Environmental Microbiology, 2007, 9(7): 1823-1835.
[10] Church M J, Björkman K M, Karl D M, Saito M A, Zehr J P. Regional distributions of nitrogen-fixing bacteria in the Pacific Ocean. Limnology and Oceanography, 2008, 53(1): 63-77.
[11] Langlois R J, Hümmer D, LaRoche J. Abundances and distributions of the dominantnifHphylotypes in the Northern Atlantic Ocean. Applied and Environmental Microbiology, 2008, 74(6): 1922-1931.
[12] Yu Z, Zhou J, Yang J, Yu X Q, Liu L M. Vertical distribution of diazotrophic bacterial community associated with temperature and oxygen gradients in a subtropical reservoir. Hydrobiologia, 2014, 741(1): 69-77.
[13] Zehr J P, Church M J, Moisander P H. Diversity, distribution and biogeochemical significance of nitrogen-fixing microorganisms in anoxic and suboxic ocean environments // Neretin L N, ed. Past and Present Water Column Anoxia. Netherlands: Springer, 2006: 337-369.
[14] Mahaffey C, Michaels A F, Capone D G. The conundrum of marine N2fixation. American Journal of Science, 2005, 305(6-8): 546-595.
[15] Dang H Y, Yang J Y, Li J, Luan X W, Zhang Y B, Gu G Z, Xue R R, Zong M Y, Klotz M G. Environment-dependent distribution of the sedimentnifH-harboring microbiota in the northern South China Sea. Applied and Environmental Microbiology, 2013, 79(1): 121-132.
[16] Farnelid H, Bentzon-Tilia M, Andersson A F, Bertisson S, Jost G, Labrenz M, Jürgens K, Riemann L. Active nitrogen-fixing heterotrophic bacteria at and below the chemocline of the central Baltic Sea. The ISME Journal, 2013, 7(7): 1413-1423.
[17] 陈因, 方大惟. 蓝藻Anabaena7120固氮的光调节. 植物生理学报, 1983, 9(1): 51-59.
[18] Howarth R W, Marino R, Cole J J. Nitrogen fixation in freshwater, estuarine, and marine ecosystems. 2. Biogeochemical controls. Limnology and Oceanography, 1988, 33: 688-701.
[19] Havens K E, James R T, East T L, Smith V H. N∶P ratios, light limitation, and cyanobacterial dominance in a subtropical lake impacted by non-point source nutrient pollution. Environmental Pollution, 2003, 122(3): 379-390.
[20] Short S M, Jenkins B D, Zehr J P. Spatial and temporal distribution of two diazotrophic bacteria in the Chesapeake Bay. Applied and Environmental Microbiology, 2004, 70(4): 2186-2192.
[21] Wang S, Qian X, Han B P, Wang Q H, Ding Z F. Physical limnology of a typical subtropical reservoir in south China. Lake and Reservoir Management, 2011, 27(2): 149-161.
[22] Yu Z, Yang J, Zhou J, Yu X Q, Liu L M, Lv H. Water stratification affects the microeukaryotic community in a subtropical deep reservoir. Journal of Eukaryotic Microbiology, 2014, 61(2): 126-133.
[23] Olson J B, Steppe T F, Litaker R W, Paerl H W. N2-fixing microbial consortia associated with the ice cover of Lake Bonney, Antarctica. Microbial Ecology, 1998, 36(3-4): 231-238.
[24] Schloss P D, Westcott S L, Ryabin T, Hall J R, Hartmann M, Hollister E B, Lesniewski R A, Oakley B B, Parks D H, Robinson C J, Sahl J W, Stres B, Thallinger G G, Van Horn D J, Weber C F. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Applied and Environmental Microbiology, 2009, 75(23): 7537-7541.
[25] Clarke K R, Gorley R N. PRIMER v6: User Manual/Tutorial. Plymouth: PRIMER-E, 2006.
[26] Kizilova A K, Titova L V, Kravchenko I K, Iutinskaya G A. Evaluation of the diversity of nitrogen-fixing bacteria in soybean rhizosphere bynifHgene analysis. Microbiology, 2012, 81(5): 621-629.
[27] Paerl H W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnology and Oceanography, 1988, 33(4part2): 823-843.
[28] Zani S, Mellon M T, Collier J L, Zehr J P. Expression ofnifHgenes in natural microbial assemblages in Lake George, New York, detected by reverse transcriptase PCR. Applied and Environmental Microbiology, 2000, 66(7): 3119-3124.
[29] Affourtit J, Zehr J P, Paerl H W. Distribution of nitrogen-fixing microorganisms along the Neuse River Estuary, North Carolina. Microbial Ecology, 2001, 41(2): 114-123.
[30] Piceno Y M, Lovell C R. Stability in natural bacterial communities: Ⅰ. Nutrient addition effects on rhizosphere diazotroph assemblage composition. Microbial Ecology, 2000, 39(1): 32-40.
[31] Bagwell C E, Lovell C R. Persistence of selectedSpartinaalterniflorarhizoplane diazotrophs exposed to natural and manipulated environmental variability. Applied and Environmental Microbiology, 2000, 66(11): 4625-4633.
[32] Shade A, Jones S E, McMahon K D. The influence of habitat heterogeneity on freshwater bacterial community composition and dynamics. Environmental Microbiology, 2008, 10(4): 1057-1067.
[33] Kerr B, Riley M A, Feldman M W, Bohannan B J. Local dispersal promotes biodiversity in a real-life game of rock-paper-scissors. Nature, 2002, 418(6894): 171-174.
[34] Kumar K, Mella-Herrera R A, Golden J W. Cyanobacterial heterocysts. Cold Spring Harbor Perspectives in Biology, 2010, 2(4): a000315.
[35] Karl D, Michaels A, Bergman B, Capone D, Carpenter E, Letelier R, Lipschultz F, Paerl H, Sigman D, Stal L. Dinitrogen fixation in the world′s oceans. Biogeochemistry, 2002, 57/58(1): 47-98.
[36] Laamanen M J. Environmental factors affecting the occurrence of different morphological forms of cyanoprokaryotes in the northern Baltic Sea. Journal of Plankton Research, 1997, 19(10): 1385-1403.
[37] Walsby A E. The permeability of heterocysts to the gases nitrogen and oxygen. Proceedings of the Royal society of London. Series B. Biological Sciences, 1985, 226(1244): 345-366.
[38] Staal M, Meysman F J R, Stal L J. Temperature excludes N2-fixing heterocystous cyanobacteria in the tropical oceans. Nature, 2003, 425(6957): 504-507.
[39] Jayakumar A, Al-Rshaidat M M D, Ward B B, Mulholland M R. Diversity, distribution, and expression of diazotrophnifHgenes in oxygen-deficient waters of the Arabian Sea. FEMS Microbiology Ecology, 2012, 82(3): 597-606.
[40] Toepel J, Welsh E, Summerfield T C, Pakrasi H, Sherman L A. Differential transcriptional analysis of the cyanobacteriumCyanothecesp. strain ATCC 51142 during light-dark and continuous-light growth. Journal of Bacteriology, 2008, 190(11): 3904-3913.
[41] Walsby A E, Hayes P K, Boje R, Stal L J. The selective advantage of buoyancy provided by gas vesicles for planktonic cyanobacteria in the Baltic Sea. New Phytologist, 1997, 136(3): 407-417.
[42] Walsby A E. Mechanisms of buoyancy regulation by planktonic cyanobacteria with gas vesicles // Fay P, Van Baalen C, eds. The Cyanobacteria. Amsterdam: Elsevier Science Publishers, 1987: 377-414.
[43] Langlois R J, LaRoche J, Raab P A. Diazotrophic diversity and distribution in the tropical and subtropical Atlantic Ocean. Applied and Environmental Microbiology, 2005, 71(12): 7910-7919.
[44] Bentzon-Tilia M, Traving S J, Mantikci M, Knudsen-Leerbeck H, Hansen J L S, Markager S, Riemann L. Significant N2fixation by heterotrophs, photoheterotrophs and heterocystous cyanobacteria in two temperate estuaries. The ISME Journal, 2015, 9(2): 273-285.
[45] Voss M, Croot P, Lochte K, Mills M, Peeken I. Patterns of nitrogen fixation along 10° N in the tropical Atlantic. Geophysical Research Letters, 2004, 31(23): L23S09.
Spatiotemporal patterns of diazotrophic microorganisms in a subtropical stratified reservoir
WANG Lina1,2, CHEN Huihuang1, LIU Lemian1, YU Zheng1, YANG Jun1,*
1 Aquatic EcoHealth Group, Key Laboratory of Urban Environment and Health, Institute of Urban Environment, Chinese Academy of Sciences, Xiamen 361021, China 2UniversityofChineseAcademyofSciences,Beijing100049,China
Biological nitrogen fixation, which is mediated by nitrogen-fixing microorganisms (diazotrophs), is an important source of fixed nitrogen in aquatic ecosystems. Recent advances in molecular biology techniques are leading to progress in the elucidation of the diazotrophic community possessing thenifHnitrogenase gene in various ecosystems. So far, the majority of studies of diazotrophic communities are based on marine samples, with freshwater habitats remaining largely unexplored. The aims of this study are to characterize the spatiotemporal patterns of a diazotrophic microbial community in a subtropical stratified reservoir (Tingxi Reservoir, southeast China), and to examine the relationships between the diazotrophic community structure and environmental variables. In this study, samples from the water column (five layers) were collected in July and October 2012, and in January and April 2013. Nitrogen-fixing microorganisms were studied using quantitative real-time PCR and clone library techniques. In Tingxi Reservoir, the water column was well mixed in winter, whereas there was clear and stable stratification in spring, summer, and autumn. Our results indicated that there were distinct spatial and seasonal patterns of abundance, activity, composition, and diversity in the diazotrophic community that was linked to water stratification in the Tingxi Reservoir. In total, 106 OTUs belonging to seven groups (i.e., 46 Cyanobacteria, 21 α-Proteobacteria, 3 β-Proteobacteria, 9 γ-Proteobacteria, 1 Firmicutes, 18 unidentified nitrogen-fixing bacteria, and 8 unknown taxa) were observed, the most diverse and dominant group of which was cyanobacteria. Both α-and γ-proteobacteria were much more common than β-proteobacteria in the reservoir.The deeper waters harbored a high percentage of unidentified bacteria and unknown taxa. The number of sequences similar to cyanobacteria in the surface water was higher than in the bottom water in spring, summer, and autumn. The Cyanobacteria OTUs number contributed more than 50% of the total OTUs in both surface and bottom waters in winter. Firmicutes were only detected in the surface water sample in October. Interestingly, our cluster analysis indicated that surface water diazotrophic communities in spring, summer, and autumn (with a stable stratification) first formed a group, then clustered with the bottom communities. However, both surface and bottom diazotrophic communities exhibited a relatively high similarity in winter due to water mixing. The pH, dissolved oxygen, and chlorophyllashowed a significant negative relationship with the DNA copy number, whereas the NOx-N showed a strong positive correlation with the DNA copy number. Water temperature and NH4-N had a negative significant relationship with RNA/DNA ratio and RNA copy number, respectively. It appeared that nitrogen-fixing bacteria showed distinctly nonrandom spatial and seasonal distributions in the Tingxi Reservoir, and their communities were either complexly structured by the thermal stratification or adapted to different environmental niches. Therefore, a stratification-based management strategy should be considered when developing methods for protecting drinking water quality and for controlling the cyanobacterial blooms.
nitrogen fixing;nifHgene; stratified reservoir; Tingxi Reservoir; spatiotemporal variation
国家自然科学基金项目(31370471, 31172114);福建省杰出青年科学基金项目(2012J06009)
2015-04-25;
日期:2016-01-05
10.5846/stxb201504250854
*通讯作者Corresponding author.E-mail: jyang@iue.ac.cn
王丽娜,陈辉煌,刘乐冕,余正,杨军.亚热带分层水库固氮微生物时空分布格局.生态学报,2016,36(18):5827-5837.
Wang L N, Chen H H, Liu L M, Yu Z, Yang J.Spatiotemporal patterns of diazotrophic microorganisms in a subtropical stratified reservoir.Acta Ecologica Sinica,2016,36(18):5827-5837.
猜你喜欢
语文世界(小学版)(2022年2期)2022-06-30 14:20:40
军民两用技术与产品(2022年2期)2022-06-01 06:29:44
当代水产(2019年9期)2019-10-08 08:02:42
当代水产(2018年8期)2018-11-02 05:30:42
广西林业科学(2016年2期)2016-03-20 05:53:24
——固氮微生物
新疆农垦科技(2016年3期)2016-02-20 23:26:18
幼儿智力世界(2015年5期)2015-08-20 09:41:39
小说林(2014年5期)2014-02-28 19:51:47
杂文选刊(2013年7期)2013-02-11 10:41:11
民生周刊(2012年25期)2012-04-29 00:44:03
