
苦荞株高及主茎分枝数的遗传相关分析
2016-11-28 02:21唐链梁成刚梁龙兵张启迪饶庆琳陈
江苏农业科学 2016年9期
唐链+梁成刚+梁龙兵+张启迪+饶庆琳+陈庆富


摘要:以高杆材料小米荞为母本、中杆材料晋荞2号为父本进行有性杂交,经自交获得杂交后代F2、F3群体株系。对亲本及杂交后代F2、F3群体株高、主茎分枝数等性状进行统计分析。结果发现,亲本小米荞的株高、单株粒质量、单株种子粒数、千粒质量与晋荞2号差异达到显著或极显著水平,但主茎分枝数差异不显著。F2、F3群体株高与主茎分枝数均呈现连续变异,符合偏正态分布,表明为数量性状。株高与主茎分枝数的遗传力平均值分别为0.912、0.503。株高与主茎分枝数、单株粒质量、单株种子粒数均呈极显著正相关,其中与单株种子粒数相关系数达到0.398**。表明株高与主茎分枝数可作为苦荞高产杂交育种的质量要目标性状。
关键词:苦荞;株高;主茎分枝数;相关分析
中图分类号: S517.03 文献标志码: A
文章编号:1002-1302(2016)09-0129-04
苦荞(Fagopyrum tataricum),蓼科(Polygonaceae)荞麦属(Fagopyrum Mill)栽培品种,别称鞑靼荞麦[1-2]。苦荞具有营养保健和食疗双质量功效,是国际粮农组织公认的粮药兼用粮种,被誉为五谷之王、三降(降血压、降血糖、降血脂)食品[3]。苦荞的黄酮含量比其他粮食作物高,能有效防治糖尿病、预防和治疗心血管硬化疾病,还能健胃消食,增强机体免疫力,对癌症也有预防作用[1,4-5]。此外,苦荞的蛋白质含量较高,其氨基酸组成、配比适宜,接近于标准蛋白,营养价值明显高于其他谷类作物[5-7]。
由于苦荞具有很高的营养价值和药用价值,日益备受人们关注。但是,苦荞的产量仍不能满足市场的需求。杂交育种早已成为大宗作物提高产量的最主要育种手段。因此,开展苦荞杂交工作,对于提高苦荞的适应性和产量水平有着重要意义。不过,苦荞花朵较小,表现出闭花受精的习性,较难进行人工去雄和有性杂交工作,制约了苦荞的杂种优势利用,导致产量一直未能有突破性的提高[8]。陈庆富等通过对苦荞开花情况的观察,发现苦荞花在刚开放时花药未开裂,也就是并未完成受精。这个未开裂的花药一般可以持续几分钟,若在这段时间里完成人工去雄授粉,则可以提高苦荞品种间的杂交成功概率[9-11]。株高、主茎分枝数、单株粒数等性状都是影响苦荞生产潜力的重要农艺性状,它们的优劣对苦荞的高产性、稳定性和适应性均有重要影响。因此,开展苦荞杂交育种工作,并对苦荞株高、主茎分枝数等参数进行遗传及相关分析,发现其遗传传递规律,以优化选择效果,增强对杂交后代的预见性,可为苦荞杂交育种研究提供一定的参考。
1 材料与方法
1.1 试验材料
本试验选用小米荞为母本、晋荞2号为父本进行有性杂交。小米荞瘦果、薄壳、无沟槽、易脱壳、迟熟、高秆、黄酮含量高[9,12]。晋荞2号瘦果、厚壳、有沟槽、不易脱壳,为早熟、中秆、抗旱、抗倒伏的高产品种[9,12-13]。
试验于2012年7月在贵州师范大学荞麦产业技术研究中心观水基地进行有性杂交工作,具体方法参照陈庆富等的操作步骤[9]。根据杂交时所使用的3个不同母本植株,使用代号A、B、C来分别表示3个杂交后代群体。将按单株收获得到的3个杂交后代种子分别播种于贵阳市永乐乡柏杨村试验基地,获得F2分离群体。F2群体单株收获后,再按单株收获种子进行播种,即为F3株系。亲本F2、F3群体的栽培均按照株间距5 cm、行间距33~40 cm的播种方式,进行常规栽培管理。
1.2 性状考察及方法
在苦荞成熟期对亲本小米荞与晋荞2号、杂交F2和F3株系群体的株高、主茎分枝数、单株粒质量和千粒质量等性状进行考种和记录。其中,杂交F2、F3群体样本量均为994株。
采用SPSS 17.0和 Excel 2010软件对试验数据进行处理分析。杂交后代目标性状的遗传力、表型和遗传变异系数等参考陈庆富等的计算方法[13-19]。
2 结果与分析
2.1 亲本及F2、F3群体的主要农艺性状
2.1.1 小米荞、晋荞2号的主要农艺性状 从表1中可以看出,亲本小米荞与晋荞麦2号之间株高、单株粒质量、单株种子粒数、千粒质量差异极显著,说明试验材料在性状上存在一定的遗传差异。不过,主茎分枝数无明显差异。小米荞的株高极显著高于晋荞麦2号,同时,其单株粒质量和单株种子粒数也极显著高于晋荞2号,不过,小米荞的千粒质量极显著低于晋荞麦2号。
2.1.2 杂交苦荞F2与F3群体的性状分析
对杂交苦荞F2与F3群体的株高和主茎分枝数进行表型值分布频率分析(图1)。结果发现,苦荞杂交后代F2和F3株系的株高和主茎分枝数性状大体上呈现出连续分布,表明为微效多基因控制的数量性状。从表2中可以看出,株高与主茎分枝数的偏度均值小于0,说明分布呈左偏。
从表2可见,F2群体和F3群体株系平均株高分别为128.38、105.94 cm;而亲本小米荞与晋荞2号平均株高分别为100.79、66.71 cm(表1),可见杂交后代群体株高超过双亲。F2、F3群体株高变异总范围分别为55~192、41~200 cm,后代群体株高变异范围较亲本大。小米荞和晋荞2号主茎平均分枝数分别为8.00、8.06个(表1);而在F2、F3群体中主茎分枝数平均值分别为6.98、7.59个,比亲本小,变幅比亲本大。主茎分枝数变异范围F2为1~12个、F3为1~13个,差异不明显。
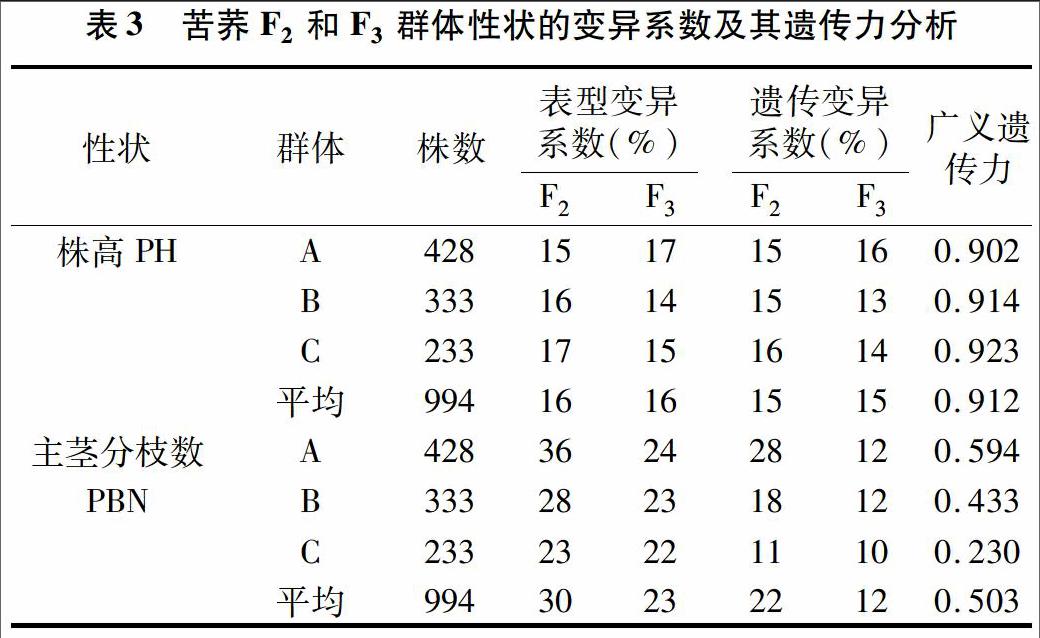
2.2 苦荞F2与F3群体表型和遗传变异系数相关分析
对苦荞杂交后代F2群体和F3群体的表型及遗传变异系数进行分析,结果发现F2、F3群体株高、主茎分枝数变异丰富,其中主茎分枝数变异系数较大,但随着世代加深,趋于稳定(表3)。F2、F3群体的株高表型变异系数平均值皆为16%,表型变异范围分别为15%~17%、14%~17%,其遗传变异系数平均值皆为15%,广义遗传力较高,均值为0.912。可见株高性状稳定性较好,遗传力较大。F2、F3群体的主茎分枝数表型变异系数平均值分别为30%、23%,表型变异范围分别为23%~36%、22%~24%,其遗传变异系数分别为22%、12%,广义遗传力均值为0.503。对于主茎分枝数性状来说,F3群体比F2群体更稳定一些。总体而言,株高遗传力大于主茎分枝数。表明苦荞在生产进化过程中,自然环境和人为作用因素对苦荞的生长发育产生很大的影响。
2.3 F2与F3 群体主要农艺性状的相关性分析
2.3.1 F2群体主要农艺性状的相关性分析
从表4可以看出,F2群体的单株粒质量与株高、主茎分枝数、单株种子粒数、千粒质量皆呈极显著正相关。说明株高、主茎分枝数、单株种子粒数、千粒质量是影响单株粒质量的重要因素。株高与主茎分枝数、单株种子粒数、单株粒质量呈现极显著正相关,相关系数平均分别为0.482**、0.434**、0.402**;与千粒质量相关性不显著。主茎分枝数与株高、单株种子粒数、单株粒质量呈极显著正相关,相关系数平均分别为0.482**、0.418**、0.376**;与千粒质量呈显著负相关,相关系数为-0.078*,说明主茎分枝数与株高、单株种子粒数、单株粒质量密切相关。单株种子粒数与株高、主茎分枝数、单株粒质量、千粒质量皆呈极显著正相关,其中与单株粒质量的相关系数最大,简单相关系数平均为0.950**。说明适当的单株种子粒数的增加有利于单株粒质量、千粒质量的提升。千粒质量与单株种子粒数、单株粒质量呈极显著正相关,但与株高的相关性不显著。
2.3.2 F3群体主要农艺性状的相关性分析
从表5可以看出,F3群体主要农艺性状的相关性与F2群体相似。同时,A、B、C群体均表现出相同的规律。单株粒质量与株高、主茎分枝数、单株种子粒数、千粒质量皆呈现极显著的正相关,其中与单株种子粒数的相关性最为紧密,平均相关系数达到0.914**。株高与主茎分枝数、单株粒质量、单株种子粒数呈极显著正相关,其中与单株种子粒数相关性最大,相关系数平均为0.398**;与千粒质量相关性不显著。说明适当的株高有利于单株种子粒数的增加。主茎分枝数与株高、单株粒质量、单株种子粒数相关性显著,相关系数平均分别为0.266**、0.393**、0.395**。单株种子粒数与株高、主茎分枝数、单株粒质量极显著正相关,与千粒质量呈极显著负相关。千粒质量与单株粒质量相关性显著,相关系数平均为0.203**;与株高、主茎分枝数相关性不显著。
3 讨论
苦荞为闭花受精作物,花朵小,较难进行人工去雄和杂交育种工作,制约了苦荞的杂种优势利用[8]。陈庆富等利用人工去雄授粉法完成对苦荞的有性杂交,可以显著提高苦荞单株产量、千粒质量,达到优化育种的目的[9-11]。姚金保等对7个小麦品种进行株高构成因素的遗传和相关性分析,表明株高的遗传可能涉及3~4对主效显性基因控制,遗传力高,株高与其构成因素呈极显著遗传正相关[19]。潘凡等研究表明单株粒质量与单株粒数、主茎分枝数呈极显著正相关,而主茎分枝数与株高呈极显著负相关[20],这与本研究结论有所不同,可能是由于材料、样本量以及环境不同所致。
本研究对亲本小米荞与晋荞2号及杂交后代群体的株高和主茎分枝数等性状进行研究,结果表明两亲本除主茎分枝数未达到显著差异外,其余性状皆达到极显著或显著差异。后代群体的株高、主茎分枝数呈现连续变异,符合偏正态分布,说明这些性状是由微效多基因控制的数量性状。后代群体株高平均值与双亲之间存在显著差异,变异范围比亲本较大,存在超亲现象;而后代群体主茎分枝数平均值与双亲之间差异不明显,变幅比亲本大,可能是环境因素所致。
杨玉霞等研究结果显示苦荞品种间株高和总分枝数的变异系数分别为14.75%、30.86%,总分枝数的变异程度较大,选择范围大[21]。株高的变异程度相对较小,选择范围窄。汪灿等研究发现苦荞品种间主要农艺性状变异丰富,株高和分枝数的变异系数分别为12.8%、14.2%[22]。嵇怡等对黄瓜株高性状进行遗传分析,发现黄瓜株高性状以主基因遗传为主,遗传率较高,表明群体的遗传变量显著高于环境误差[23]。本研究结果显示小米荞与晋荞2号杂交后代F2、F3群体株高和主茎分枝数表型变异系数均值分别为16%、30%、16%、23%,可见株高性状稳定性较好,遗传力较大。F2代到F3代的株高和主茎分枝数的表型与遗传变异系数均变小,暗示了随着世代的增多,群体性状趋于稳定,选择范围变小。本研究发现F2、F3群体株高、主茎分枝数的广义遗传力分别为0.912、0.503,可见株高的遗传力更大,主茎分枝数受环境条件、人为因素影响较大。
汪灿等认为株高、分枝数、单株粒质量、千粒质量是影响苦荞产量的主要因素[22]。杨明君等认为不同性状对苦荞产量的影响大小顺序为株高>主茎节数>单株粒质量>单株粒数>千粒质量,选择高杆、主茎节数多、单株粒质量高、单株粒数多的品种是提高苦荞产量的主要途径[24]。李秀莲等研究发现苦荞各性状间的表行相关与遗传相关方向相同,表明在育种实践中可以根据表型相关来选择所需性状[12]。本研究发现在F2、F3群体相关分析中,单株粒质量与株高、主茎分枝数、单株种子粒数、千粒质量皆呈极显著正相关,这与前人的研究结果一致。本研究发现后代群体的株高与主茎分枝数平均水平上呈极显著正相关。表明可以把株高和主茎分枝数作为苦荞高产选育的质量要目标性状。同时,也应把单株种子粒数、千粒质量作为提高产量一个质量要参数。协调好各性状之间、各性状与产量之间的关系,从而选育出优质高产荞麦。
参考文献:
[1]陈庆富. 荞麦属植物科学[M]. 北京:科学出版社,2012:3-5.
[2]林汝法. [HJ1.83mm]中国荞麦[M]. 北京:中国农业出版社,1994:160.
[3]万丽英,穆建稳. 贵州苦荞的营养保健功能与开发利用价值[J]. 贵州农业科学,2004,32(2):74-75.
[4]杨克理. 我国荞麦种质资源研究现状与展望[J]. 作物品种资源,1995(3):11-13.
[5]黄元射,李 明,孙富年,等. 苦荞品种在质量庆低海拔地区的主要经济性状表现[J]. 种子,2008,27(4):66-68.
[6]张美莉,胡小松. 荞麦生物活性物质及其功能研究进展[J]. 杂粮作物,2004,24(1):26-29.
[7]柴 岩,刘荣厚. 荞麦的营养成分与营养价值[C]. 中国荞麦科学研究论文集,1990:198-202.
[8]Jacquemart A L,Ledent J F,Quinet M,et al. (Fagopyrum esculentum Moench) still a valuable crop today? [J]. European Journal of Plant Science & Biotechnology,2012,6(Special issue 2):1-10.
[9]陈庆富,陈其饺,石桃雄,等. 苦荞厚果壳性状的遗传及其与产量因素之间的相关性研究[J]. 作物杂志,2015,165(2):27-31.
[10]Chen Q F. A study of resources of Fagopyrum (Polygonaceae)native to China[J]. Botanical Journal of the LinneanSociety,1999,130(1):53-64.
[11]Chen Q F. Wide hybridization among Fagopyrum (Polygonaceae) species native to China[J]. Botanical Journal of the LinneanSociety,1999,131:177-185.
[12]李秀莲,史兴海,高伟等. 苦荞新品种‘晋荞麦2号丰产稳产性分析及应用前景[J]. 农学学报,2011,1(12):6-10.
[13]陈庆富. 生物统计学[M]. 北京:高等教育出版社,2011:91-163.
[14]姚世鸿,王景佑,陈庆富. 遗传学[M]. 贵阳:贵州人民出版社,2001:157-175.
[15]李春喜,邵 云,姜丽娜. 生物统计学[M]. 北京:科学出版社,2008:85-92.
[16]孔繁玲. 植物数量遗传学[M]. 北京:中国农业大学出版社,2006:1-7,38-56,160-176.
[17]马惠馨. 辣(甜)椒杂种F2数量性状遗传相关与遗传的研究初报[J]. 沈阳农业大学学报,1988,19(1):15-20.
[18]杨光宇,郑惠玉,韩春凤. 栽培大豆(G.max)×半野生大豆(G.gracilis)后代主要农艺性状遗传参数的初步分析[J]. 作物学报,1992,18(6):439-446.
[19]姚金保,任丽娟,张平平,等. 小麦株高及其构成因素的遗传及相关性分析[J]. 麦类作物学报,2011,31(4):604-610.
[20]潘 凡,石桃雄,陈其皎,等. 苦荞种植主要农艺性状的变异及其对单株粒质量的贡献研究[J]. 植物科学学报,2015,33(6):829-839.
[21]杨玉霞,吴 卫,郑有良,等. 苦荞主要农艺性状与单株籽质量的相关和通径分析[J]. 安徽农业科学,2008,36(16):6719-6721.
[22]汪 灿,胡 丹,杨 浩,等. 苦荞主要农艺性状与产量关系的多质量分析[J]. 作物杂志,2013(6):18-22.
[23]嵇 怡,徐 强,陈学好. 黄瓜株高性状遗传模型分析[J]. 扬州大学学报,2009,30(3):75-79.
[24]杨明君,杨 媛,郭忠贤,等. 旱作苦荞麦籽粒产量与主要性状的相关分析[J]. 内蒙古农业科技,2010(2):49-50.
猜你喜欢
农村百事通(2019年17期)2019-10-08
青年歌声(2018年5期)2018-10-29
现代农业科技(2017年1期)2017-03-06
广东饲料(2016年3期)2016-12-01
中国集体经济(2016年26期)2016-11-19
中国粮油学报(2014年8期)2014-02-06
