
白菜型冬油菜抗寒相关性状的遗传分析
2016-11-14 01:32:18刘林波孙万仓刘自刚武军艳李学才曾秀存袁金海
中国农业科学 2016年21期
刘林波,孙万仓,刘自刚,武军艳,方 彦,李学才,曾秀存,杨 刚,董 云,陈 奇,方 园,袁金海
白菜型冬油菜抗寒相关性状的遗传分析
刘林波1,孙万仓1,刘自刚1,武军艳1,方 彦1,李学才1,曾秀存2,杨 刚1,董 云3,陈 奇1,方 园1,袁金海1
(1甘肃农业大学农学院/甘肃省油菜工程技术研究中心/甘肃省作物遗传改良与种质创新重点实验室/甘肃省干旱生境作物学重点实验室,兰州 730070;2河西学院,甘肃张掖734000;3甘肃省农业科学院,兰州 730070)
【目的】了解白菜型冬油菜抗寒相关性状的遗传方式,为油菜抗寒性的遗传育种提供一定的理论依据。【方法】以6个抗寒性不同的白菜型冬油菜品种(系)为亲本材料,按照GriffingⅠ配制完全双列杂交组合,将亲本和F1种植后测定抗寒性相关的性状以获得试验数据。采用QGAStation软件中的加性-显性遗传模型和MINQUE(1)法,对白菜型冬油菜越冬率及11个理化性状的遗传方差分量比率、遗传相关性、亲本加性效应、组合显性效应和遗传率以及杂种优势进行分析。【结果】越冬率、POD活性、SOD活性、可溶性糖含量、相对电导率、叶绿素含量、蒸腾速率和气孔导度的遗传主要以加性效应为主,显性效应影响较小;CAT活性、丙二醛含量、可溶性蛋白含量和净光合速率则主要以显性效应为主。在基因型和表现型的相关分析上,越冬率与CAT活性、POD活性、SOD活性、可溶性蛋白含量、可溶性糖含量、叶绿素含量、蒸腾速率、气孔导度和净光合速率呈极显著正相关,与丙二醛含量和相对电导率则呈极显著负相关。陇油7号和陇油8号在越冬率、CAT活性、POD活性、SOD活性、可溶性蛋白含量、可溶性糖含量、叶绿素含量、蒸腾速率、气孔导度和净光合速率上的加性效应均呈正值,表明陇油7号和陇油8号可用作提高后代抗寒性的亲本。显性效应分析表明,陇油7号×GY和陇油7号×陇油8号是抗寒性强的组合。狭义遗传率以SOD活性、相对电导率、蒸腾速率和越冬率的表现最好,分别为74.63%、59.06%、57.49%和56.87%,CAT活性最低,为2.83%。CAT活性、可溶性蛋白含量和净光合速率3个抗寒相关性状具有很强的群体平均优势和群体超亲优势。【结论】越冬率、SOD活性、相对电导率和蒸腾速率适宜在早代选择,亲本评价分析指出,陇油7号的一般配合力高, 可作为冬油菜抗寒性改良的首选亲本。
冬油菜;加性效应;显性效应;抗寒性;杂种优势
0 引言
【研究意义】白菜型油菜(L.,2n=20,AA)是中国栽培历史最悠久的油菜种类,具有抗旱、早熟、抗寒等突出优点[1],因而是北方地区重要的油料作物和生态作物[2],由于成熟早(5月下旬),成为促进北方耕作制度改革的先驱作物。选育具有优异抗寒性的强冬性白菜型冬油菜品种仍然是该地区发展冬油菜需要优先研究的重大课题[3]。【前人研究进展】国内外对油菜产量性状[4-7]、农艺性状[8-10]和品质性状[11-13]的杂种优势及遗传分析进行了研究。对于油菜抗寒性的研究主要基于生理生化指标、抗冻蛋白和光合特性,杨宁宁等[14]对冬油菜进行研究,发现可溶性蛋白含量的变化是影响冬油菜抗寒性的决定性因素。刘自刚等[15]研究认为白菜型冬油菜在低温胁迫阶段,强抗寒品种的净光合速率较高,可溶性蛋白含量与抗寒性呈显著正相关。史鹏辉等[16]研究表明低温下冬油菜根部抗氧化酶活性变化与抗寒性之间存在相关性。刘自刚等[17]研究了夜间低温对白菜型冬油菜光合结构的影响,发现当温度降低至5℃时,不同抗寒品种的气孔导度、净光合速率和蒸腾速率下降,叶绿素含量升高。孔德晶等[18]利用通径分析对冬油菜的生理生化指标进行分析,发现它们对越冬率的重要性为:幼苗习性>SOD活性>MDA含量>游离脯氨酸含量>POD活性>CAT活性>可溶性蛋白含量。朱惠霞等[19]通过研究8个白菜型冬油菜在不同低温条件下的生理生化指标,发现POD活性、可溶性蛋白含量和可溶性糖含量与品种抗寒性关系密切,可作为选择抗寒性品种的指标。蒲媛媛等[20]认为SOD活性、CAT活性、可溶性蛋白含量和MDA含量在一定程度上能反映白菜性冬油菜的抗寒性。关于植物抗寒性的遗传研究在其他作物上报道甚多,常硕其等[21]研究认为选育耐冷性强的亲本,可提高杂交稻的耐冷能力。贺荣华等[22]研究发现水稻苗期抗寒性的杂种优势表现为中亲优势。裴玉贺等[23]通过研究玉米生理指标发现玉米抗寒性主要是细胞核遗传,POD活性和脯氨酸含量主要受加性效应的影响,可溶性蛋白含量和丙二醛含量主要受非加性效应的影响,可溶性糖含量受加性与非加性效应的共同作用。【本研究切入点】近年来,已经对油菜农艺性状和品质性状的遗传分析进行了大量的研究,但针对油菜抗寒相关性状的遗传研究却尚未见报道。【拟解决的关键问题】本研究以6个抗寒性不同的白菜型冬油菜品种(系)为亲本,按Griffing方法Ⅰ配制6×6完全双列杂交组合,根据朱军[24]提出的混合线性模型中的加性-显性(AD)遗传模型,对白菜型冬油菜越冬率及抗寒相关的性状进行分析,以估算遗传中各方差分量和各性状遗传相关性,为鉴定抗寒基因资源以及冬油菜抗寒性的遗传改良提供理论依据。
1 材料与方法
1.1 试验材料
试验以6个白菜型冬油菜品种(系)为亲本,品种(系)的抗寒性是根据孙万仓等[2]利用越冬率作为划分标准进行的鉴定。按Griffing方法Ⅰ配制杂交组合,6个亲本的来源及特性详见表1。
1.2 试验设计及试验点气候因素
试验材料于2014年8月22日播种在甘肃省兰州市上川镇甘肃省油菜工程技术研究中心试验基地,试验地纬度36°03¢、经度103°40¢,海拔高度2 180 m,该试验点的气象数据如表2所示。完全随机区组设计,设3次重复,每一小区种3行,行间距20 cm,行长为1.5 m,株距10 cm,出苗后及时间苗,五叶期定苗。于2014年11月29日(-10℃)取样,每一小区随机取3株,每株取同一生长位点健壮功能叶,将取的样品置于自封袋且放在冰盒中带回实验室,之后对相关指标进行测定,田间管理同常规大田管理方法。
1.3 相关指标的测定方法
1.3.1 越冬率调查 分别于油菜枯叶期前(2014年11月29日)和翌年返青后(2015年4月10日)统计各小区的植株存活苗数,并计算越冬率(越冬率=(返青后存活苗数/冬前基本苗数)×100%)。

表1 试验材料来源及特性

表2 试验点主要气象因子
1.3.2 生理生化指标测定 按照邹琦[25]的方法进行叶片生理生化指标的测定,超氧化物歧化酶(superoxide dismutase,SOD,U∙g-1FW)活性采用氮蓝四唑光还原法测定。过氧化物酶(peroxidase,POD,U∙g-1∙min-1)活性采用愈创木酚比色法测定。过氧化氢酶(hydrogen peroxidase,CAT,U∙g-1∙min-1)活性采用紫外吸收法测定。丙二醛(malondialdehyde,MDA,µmol∙g-1)含量采用硫代巴比妥酸法测定。可溶性蛋白(soluble protein,SP,mg∙g-1)含量采用考马斯亮蓝G-250法测定。可溶性糖(soluble sugar,SS,%)含量采用蒽酮显色法测定。相对电导率(%)采用DDS-302+纯水相对电导率仪进行测定。
1.3.3 叶绿素含量测定 于2014年11月29日在试验地每小区随机取3株,每株选同一部位新展开的叶片,采用SPAD-502 plus叶绿素计测定叶绿素(mg∙g-1)含量。
1.3.4 光合参数测定 冬前低温胁迫阶段,在天气晴朗的上午(10:00—12:00),采用LI-6400便携式光合仪测定光合参数,每小区随机取3株,每株选同一部位新展开的叶片测定并记录3组数据取平均值,测定参数:净光合速率(net photosynthetic rate,n,µmol CO2∙m-2∙s-1)、气孔导度(stomatal conductance,s,mol H2O∙m-2∙s-1)、蒸腾速率(transpiration rate,r,mmol H2O∙m-2∙s-1)等,测定条件为叶室(2 cm×3 cm)温度控制在20—26℃,光强为1 000 µmol∙m-2∙s-1,流速为200 µmol∙s-1。
1.4 统计分析
运用朱军[26]提出的基因型与环境互作的加性-显性(AD)遗传模型进行数据分析,采用MINQUE(1)法估算性状的各项遗传方差分量,采用调整无偏预测法(AUP法)预测各项遗传效应和杂种优势值,采用以区组为抽样单位的Jackknife抽样方法估算各项统计量的标准误差,并用-test对参数进行显著性检验,所有数据的运算和分析采用QGAStation软件进行。
2 结果
2.1 亲本及F1抗寒相关性状的平均表现
与抗寒相关的12个性状中,F1的越冬率、CAT活性、SOD活性、可溶性蛋白含量的均值、极小值和极大值均大于亲本,POD活性、蒸腾速率和净光合速率与亲本相近,而亲本的丙二醛含量和相对电导率大于F1。亲本越冬率、CAT活性、SOD活性、蒸腾速率、气孔导度和净光合速率的变异系数大于F1,其他性状的变异系数则F1大于亲本(表3)。综合各性状的特征值来看,F1的POD活性、SOD活性、可溶性糖含量、相对电导率、叶绿素含量、蒸腾速率等性状的表现介于双亲之间,而越冬率、CAT活性、SOD活性、可溶性蛋白含量、净光合速率表现出一定的超亲优势。
2.2 越冬率与生理生化和光合性状间的相关性分析
通过进行相关性分析(表4),越冬率与CAT活性、POD活性、SOD活性、可溶性蛋白含量、可溶性糖含量呈极显著正相关关系,与丙二醛含量和相对电导率呈极显著负相关关系。越冬率与叶绿素含量呈显著正相关关系,与蒸腾速率、气孔导度、净光合速率呈极显著的正相关关系。表明这些性状与抗寒性密切相关。
2.3 越冬率与各抗寒性状间的遗传相关分析
表现型与基因型相关分析表明(表5),越冬率与CAT活性、POD活性、SOD活性、可溶性蛋白含量、可溶性糖含量、蒸腾速率和净光合速率呈极显著正相关,与丙二醛含量和相对电导率呈极显著负相关。在所有显著性相关的性状中,SOD活性、可溶性蛋白含量和蒸腾速率相关系数相对较高,表现型与基因型相关系数都在0.60以上。越冬率与各抗寒相关性状间的加性相关都达到极显著水平,除与丙二醛含量和相对电导率呈负相关外,与其他性状均呈正相关,且越冬率与CAT活性、SOD活性、可溶性蛋白含量、可溶性糖含量、叶绿素含量、气孔导度和净光合速率的加性相关系数达到最高。越冬率与CAT活性、SOD活性和可溶性蛋白含量呈显著和极显著正相关,而与丙二醛含量呈极显著负相关。
2.4 抗寒相关性状的遗传方差分析
通过对抗寒相关性状的方差分析(表6),发现各性状的遗传效应表现各不相同,对于12个抗寒相关性状的加性方差比率和显性方差比率,除了CAT活性、POD活性和SOD活性的加性方差比率或显性方差比率不显著外,其余性状均达到显著或极显著水平,对于不同的性状,其加性方差比率和显性方差比率所占比重有一定差异,越冬率、POD活性、SOD活性、可溶性糖含量、相对电导率、叶绿素含量、蒸腾速率和气孔导度的加性方差比率大于显性方差比率,表明这8个性状主要受加性效应控制,而CAT活性、丙二醛含量、可溶性蛋白含量和净光合速率的显性方差比率大于加性方差比率,说明这些性状的遗传以显性效应为主。
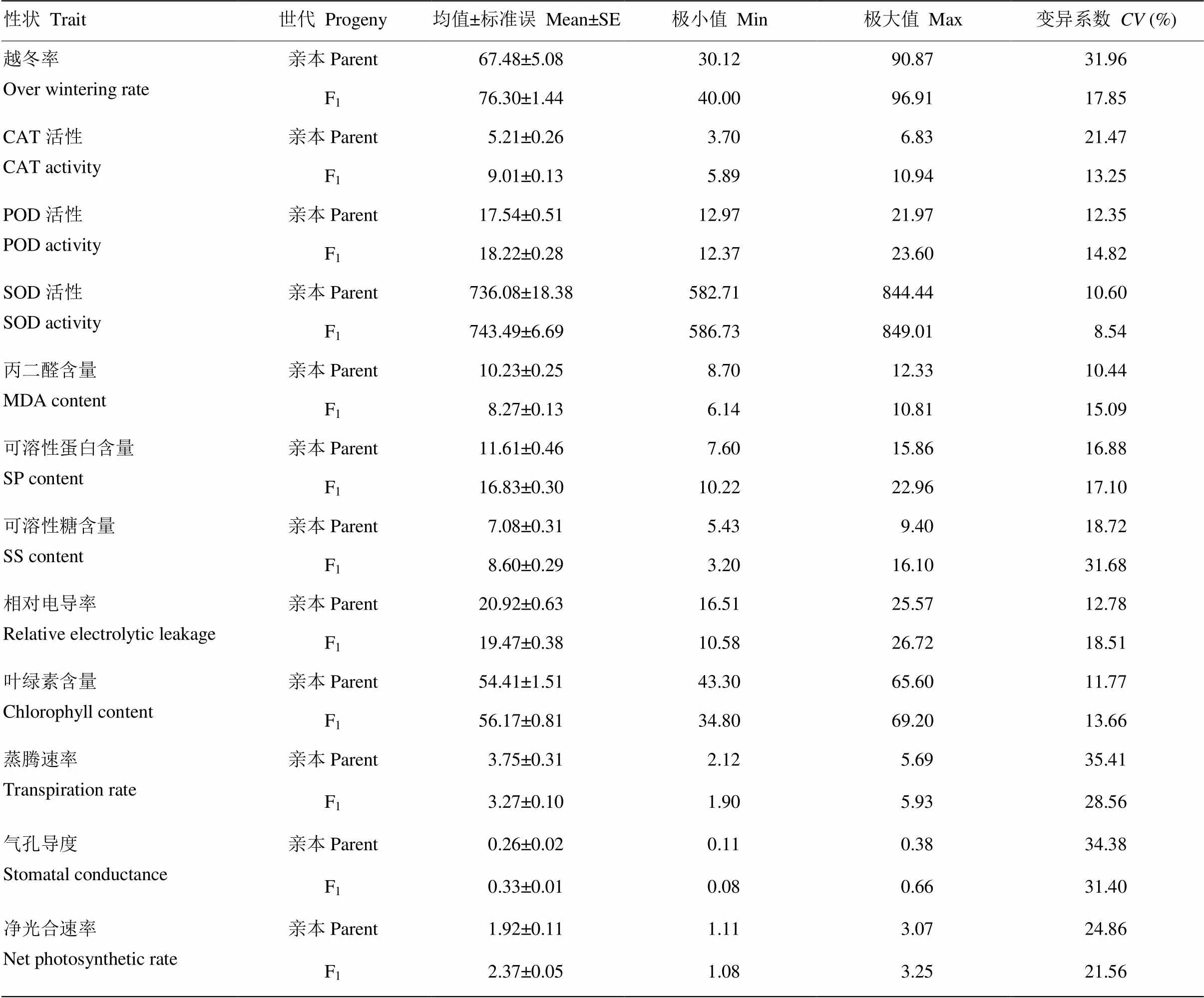
表3 亲本和F1抗寒相关性状的极值、平均值和变异系数

表4 越冬率与11个性状间的相关系数
*表示在5%水平时差异显著;**表示在1%水平时差异极显著。下同
* indicate significant difference at 5% level; ** indicate significant difference at 1% level. The same as below
2.5 亲本抗寒相关性状的加性遗传效应
在越冬率上,陇油7号、陇油8号、陇油9号和GY具有正向加性效应,天油2号和Nieb具有负向加性效应(表7),其中,陇油7号和Nieb达到极显著水平,陇油8号达到显著水平。具有正向加性效应的亲本中,陇油7号最高,达10.467,其余依次为陇油8号、陇油9号和GY,加性效应值分别为6.665、1.941和1.030。具有负向加性效应的亲本中,负向加性效应值最大的为天油2号,为-5.504,最小的为Nieb,为-14.600。陇油7号在丙二醛含量和相对电导率上的加性效应为负值,其余9个性状均为正值,且CAT活性、SOD活性、可溶性蛋白含量、可溶性糖含量、叶绿素含量、蒸腾速率、气孔导度、净光合速率都达到显著或极显著。陇油8号在丙二醛含量、相对电导率和叶绿素含量上的加性效应为负值,其余性状的加性效应均为正值,其中,在越冬率、CAT活性、SOD活性、可溶性糖含量和气孔导度上的值仅次于陇油7号并达到显著或极显著水平,说明以陇油7号和陇油8号作为亲本,在一定程度上可以提高其后代的抗寒性。Nieb除丙二醛含量和相对电导率的加性效应值为正值外,其余性状的加性效应值均为负值,故Nieb是参试材料中抗寒性最差的亲本,以Nieb作为亲本,不利于油菜抗寒性的改良。
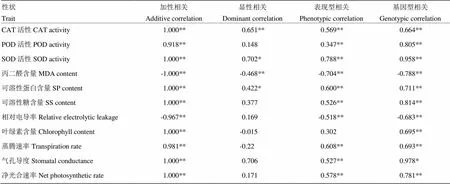
表5 越冬率与抗寒相关性状间遗传相关系数

表6 抗寒相关性状的方差分量比率估计值
Va:加性方差;Vb:显性方差;Vp:表型方差;Ve:机误方差 Va: Additive variance; Vb: Dominant variance; Vp: Phenotypic variance; Ve: Error variance
2.6 杂交组合抗寒相关性状的显性遗传效应
通过对杂交组合的抗寒性分析(表8),4个性状在15个杂交组合上的显性效应有正效应也有负效应,其中,组合陇油7号×GY、陇油7号×陇油8号在CAT活性、可溶性蛋白含量和净光合速率上显性效应表现突出。这些组合也反映出效应值大小与亲本的抗寒性密切相关。
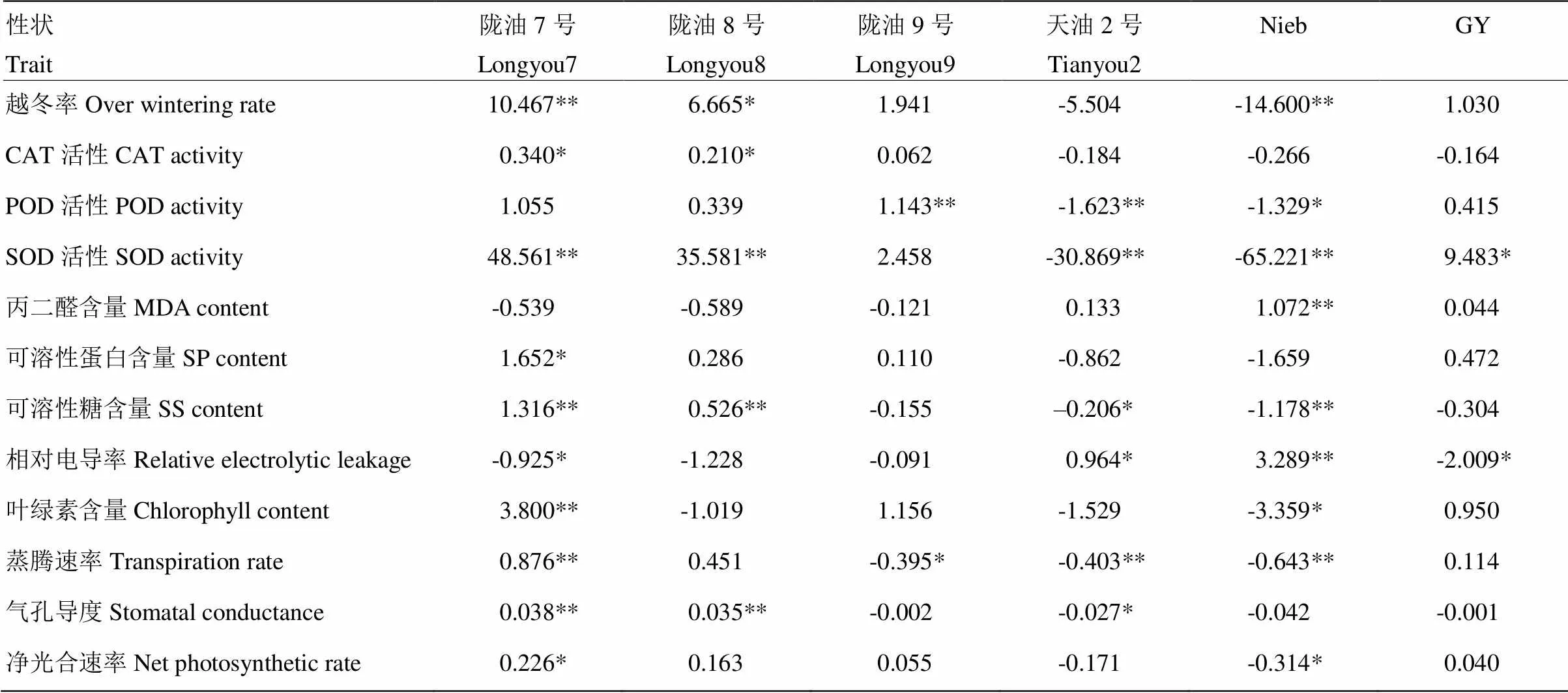
表7 亲本各抗寒相关性状的加性效应预测值

表8 F1杂交组合显性效应预测值
2.7 抗寒相关性状的遗传率分析
除CAT活性的狭义遗传率外,其余12个性状的狭义遗传率和广义遗传率都达到极显著水平,但各性状间差异表现较大(表9)。越冬率、SOD活性、相对电导率和蒸腾速率的狭义遗传率都达到50%以上,表明在育种中可以对这4个性状进行早代选择。在广义遗传率方面,CAT活性的广义遗传率达到最高,为96.18%,其次是可溶性蛋白含量、丙二醛含量、相对电导率、SOD活性、蒸腾速率、越冬率和净光合速率,它们的广义遗传率均在60%以上,表明这些性状遗传受环境影响较小。POD活性、可溶性糖含量、叶绿素含量和气孔导度的广义遗传率较低,受环境影响大,故对其进行遗传改良时应注重栽培条件等环境因素对选择的影响。
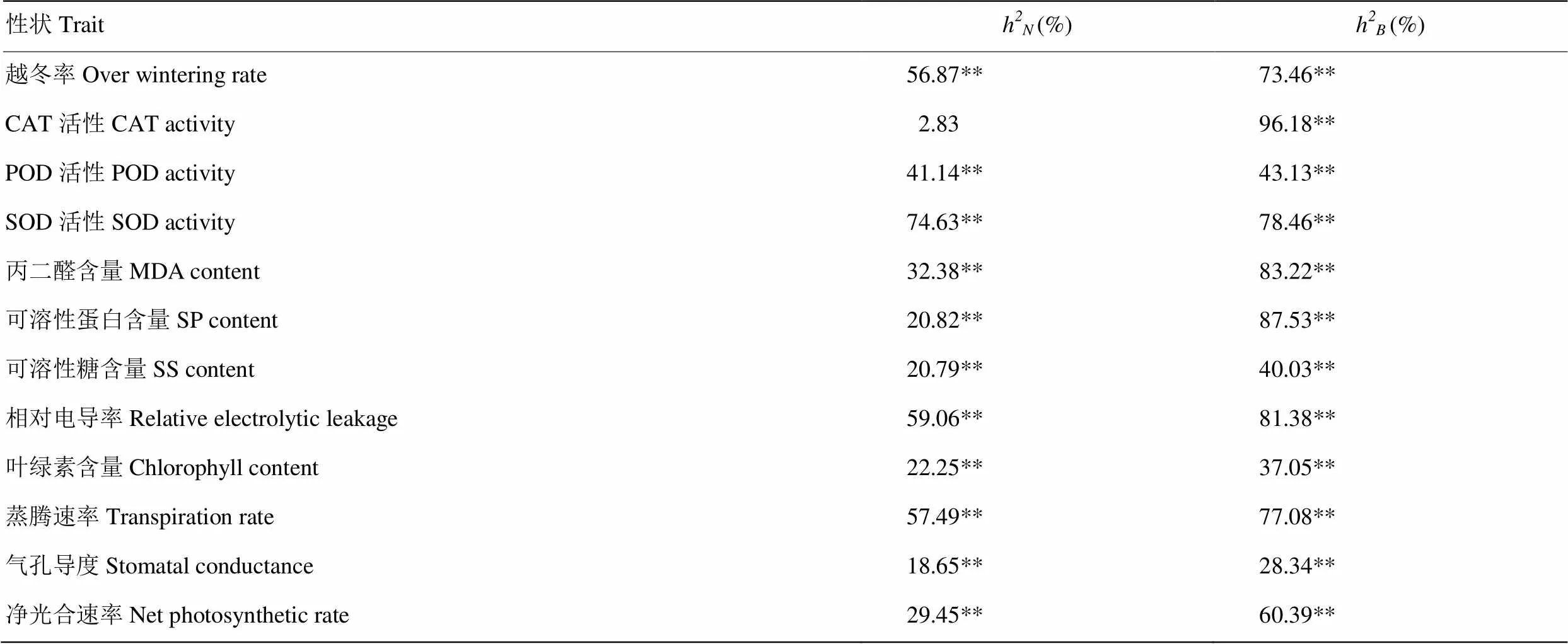
表9 抗寒相关性状的遗传率
h:狭义遗传率;h:广义遗传率
h: Heritability in the narrow sense;h: Heritability in the broad sense
2.8 抗寒相关性状的基因型值和杂种优势分析
除CAT活性、丙二醛含量和可溶性蛋白含量在F1上的基因型预测值达到显著或极显著外,其他性状在F1和F2上的基因型预测值均未达到显著水平(表10),表明白菜型冬油菜在CAT活性、丙二醛含量和可溶性蛋白含量上的基因型易受环境条件的影响。除POD活性、SOD活性、可溶性糖含量和叶绿素含量外,其他性状在F1和F2上的群体平均优势均达到显著或极显著水平,丙二醛含量、相对电导率和蒸腾速率的群体平均优势为负值,其中CAT活性在F1和F2上的群体平均优势最大,分别为0.485(F1)和0.242(F2)。除可溶性糖含量、叶绿素含量、气孔导度外,其他性状在F1和F2上的群体超亲优势达到显著或极显著水平,其中CAT活性、可溶性蛋白含量和净光合速率均表现为正向超亲优势,且这3个性状的F1、F2超亲优势均大于5%。
各杂交组合抗寒相关性状杂种优势的实测结果(表11)与基于模型预测的杂种优势值(表10)相比,二者在F1抗寒相关性状杂种优势上的表现一致,且CAT活性在群体平均优势、群体超亲优势、平均优势和超亲优势上均表现最大,但实测结果比模型预测的抗寒相关性状杂种优势值偏大,产生这种差异的原因可能是模型预测采用群体平均数µ的无偏估计值(双亲杂交所有亲本和组合的算术平均数)来估算群体平均优势和群体超亲优势[27]。
3 讨论
抗寒性是植物抵御外界逆境的一种能力,但植物的抗寒性表现为复杂的数量性状,其遗传变异来源复杂。郭海林等[28]研究表明结缕草属植物的抗寒性具有主基因+多基因遗传特征,符合B-4遗传模型,即抗寒性受到2对等加性的主效基因控制。王鹏良等[29]研究发现假俭草的抗寒性符合B-1遗传模型,受2对加性-显性-上位性主效基因控制,主基因的遗传率为91.28%。张扬勇等[30]对结球甘蓝研究结果也认为抗寒性遗传符合加性-显性模型,并以加性效应为主。植物的性状主要是由基因的加性效应和非加性效应控制的,对于同一物种的不同性状和不同物种的同一性状,基因控制类型是各不相同的。本研究利用QGAStation软件对白菜型冬油菜12个抗寒相关性状进行了遗传分析,结果表明,越冬率、POD活性、SOD活性、可溶性糖含量、相对电导率、叶绿素含量、蒸腾速率和气孔导度主要以加性效应为主,CAT活性、丙二醛含量、可溶性蛋白含量和净光合速率主要以显性效应为主。性状间加性相关系数决定是否可在早代对某一个或几个性状进行间接选择[31],本文发现CAT活性、SOD活性、可溶性蛋白含量、可溶性糖含量、叶绿素含量、气孔导度和净光合速率与越冬率加性相关系数呈正向极显著相关,表明在油菜抗寒育种中可通过定向选择CAT活性、SOD活性、可溶性蛋白含量、可溶性糖含量、叶绿素含量、气孔导度和净光合速率来间接提高冬油菜的抗寒性。越冬率、SOD活性、相对电导率和蒸腾速率的狭义遗传率均大于50%,说明这4个性状适宜在早期世代进行选择。在油菜育种中亲本的遗传评价是非常重要的[32],它是抗寒性改良的基础。本研究所用6个亲本中,陇油7号是一般配合力最大的亲本。此外,陇油7号组配的杂交组合:陇油7号×GY和陇油7号×陇油8号在CAT活性、可溶性蛋白含量和净光合速率上的显性效应表现突出。故在培育油菜抗寒品种时,陇油7号是选育强抗寒品种的最佳亲本。
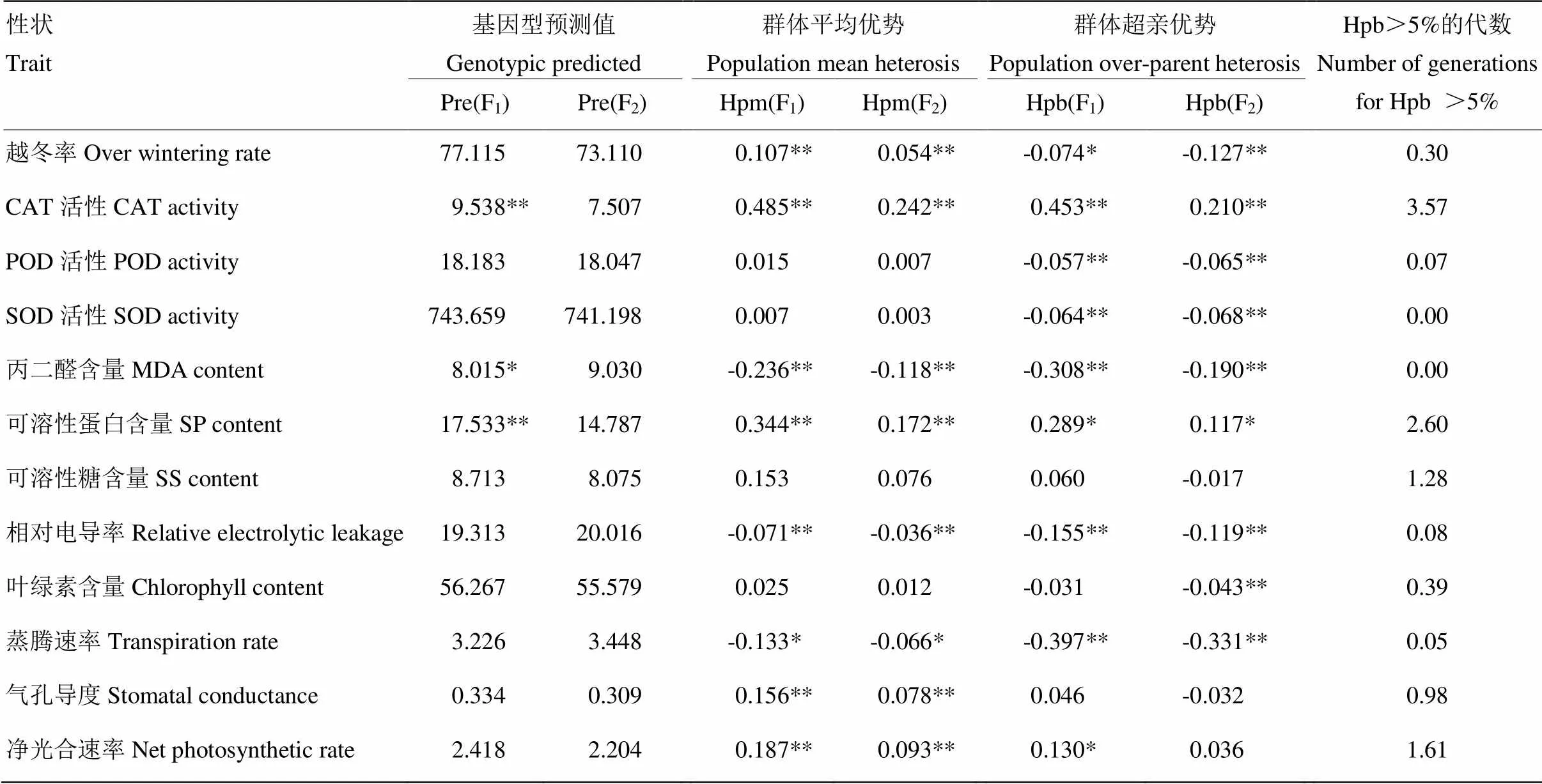
表10 抗寒相关性状的杂种优势预测值

表11 F1抗寒相关性状杂种优势及其变异幅度
4 结论
越冬率、POD活性、SOD活性、可溶性糖含量、相对电导率、叶绿素含量、蒸腾速率和气孔导度等抗寒相关性状的遗传以加性效应为主,其中,越冬率、SOD活性、相对电导率、蒸腾速率的狭义遗传率大于50%且极显著,宜在早代选择。陇油7号可作为冬油菜抗寒性改良的首选亲本。
References:
[1] 刘后利. 几种芸薹属油菜的起源和进化. 作物学报, 1984, 10(1): 9-18.
Liu H L. Origin and evolution of rapeseeds., 1984, 10(1): 9-18. (in Chinese)
[2] 孙万仓, 马卫国, 雷建民, 刘秦, 杨仁义, 武军艳, 王学芳, 叶剑, 曾军, 张亚宏, 康艳丽, 郭秀娟, 魏文惠, 杨杰, 蒲媛媛, 曾潮武, 刘红霞. 冬油菜在西北旱寒区的适应性和北移的可行性研究. 中国农业科学, 2007, 40(12): 2716-2726.
Sun W C, Ma W G, Lei J M, Liu Q, Yang R Y, Wu J Y, Wang X F, Ye J, Zeng J, Zhang Y H, Kang Y L, Guo X J, Wei W H, Yang J, Pu Y Y, Zeng C W, Liu H X. Study on adaptation and introduction possibility of winter rapeseed to dry and cold areas in north-west china., 2007, 40(12): 2716-2726. (in Chinese)
[3] 孙万仓, 曾秀存, 刘自刚, 杨刚, 方彦, 武军艳, 李学才. 轮回选择对白菜型冬油菜抗寒性及经济性状的影响. 中国油料作物学报, 2015, 37(4): 443-452.
Sun W C, Zeng X C, Liu Z G, Yang G, Fang Y, Wu J Y, Li X C. Effects of recurrent selection on cold tolerance and economic traits of.,2015, 37(4): 443-452. (in Chinese)
[4] 沈金雄, 傅廷栋, 杨光圣, 马朝芝, 涂金星. 甘蓝型油菜杂种优势及产量性状的遗传改良. 中国油料作物学报, 2005, 27(1): 5-9.
Shen J X, Fu T D, Yang G S, Ma C Z, Tu J X. Analysis of heterosis reveals genetic improvement for yield traits in rapessed (L.)., 2005, 27(1): 5-9. (in Chinese)
[5] 邓武明, 阳小虎, 文凤君, 陈胜荣, 赵昌斌. 甘蓝型油菜产量性状的遗传及相关与通径分析. 中国油料作物学报, 2004, 25(4): 27-30.
Deng W M, Yang X H, Wen F J, Chen S R, Zhao C B. Inheritance model, correlation and path analysis of yield in rape (L).,2004, 25(4): 27-30. (in Chinese)
[6] Ali M, Copeland L O, Elias S G, Kelly J D. Relationship between genetic distance and heterosis for yield and morphological traits in winter canola (L.)., 1995, 91(1): 118-121.
[7] Pradhan A K, Sodhi Y S, Mukhopadhyay A, Pental D. Heterosis breeding in Indian mustard (L.Czern & Coss): analysis of component characters contributing to heterosis for yield., 1993, 69(3): 219-229.
[8] Jiang J, Pu X, Zhang J, LI H, CHAI L, HUANG C, HU H, ZHENG B, NIU Y, JIANG L. Analysis on combining ability and heritability of the main agronomic characters for JA CMS inL.., 2014, 15(1): 39.
[9] 魏忠芬, 李德文, 王军, 张太平. 甘蓝型杂交油菜亲本含油量及主要农艺性状配合力与遗传力分析. 西北农业学报, 2014, 23(6): 102-108.
Wei Z F, Li D W, Wang J, Zhang T P. Analysis on combining ability and heritability of main agronomic traits and oil contents of hybrid parents inL.., 2014, 23(6): 102-108. (in Chinese)
[10] 姜磊, 陶诗顺, 张敏, 黄霞, 彭雅利. 甘蓝型油菜亲本主要农艺性状的配合力与遗传力分析. 西北农业学报, 2011, 3(2): 5.
Jiang L, Tao S S, Zhang M, Huang X, Peng Y L. Analysis on combining ability and genetic ability of main agronomic characters inL.., 2011, 3(2): 5. (in Chinese)
[11] 刘定富, 刘后利. 甘蓝型油菜脂肪酸成分的基因作用形式和效应. 作物学报, 1990, 16(3): 193-199.
Liu D F, Liu H L. Gene action and effects of fatty acids inL.., 1990, 16(3): 193-199. (in Chinese)
[12] 戚存扣, 盖钧镒, 章元明. 甘蓝型油菜芥酸含量的主基因+多基因遗传. 遗传学报, 2001, 28(2): 182-187.
Qi C K, Gai J Y, Zhang Y M. Major gene plus poly-gene inheritance of erucic acid content inL.., 2001, 28(2): 182-187. (in Chinese)
[13] Burns M J, Barnes S R, Bowman J G, Clarke M H E, Werner C P, Kearsey M J. QTL analysis of an intervarietal set of substitution lines in(i) Seed oil content and fatty acid composition., 2003, 90(1): 39-48.
[14] 杨宁宁, 孙万仓, 刘自刚, 史鹏辉, 方彦, 武军艳, 曾秀存, 孔德晶, 鲁美宏, 王月. 北方冬油菜抗寒性的形态与生理机制. 中国农业科学, 2013, 47(3): 452-461.
Yang N N, Sun W C, Liu Z G, Shi P H, Fang Y, Wu J Y, Zeng X C, Kong D J, Lu M H, Wang Y. Morphological characters and physiological mechanisms of cold resistance of winter rapeseed in Northern China., 2013, 47(3): 452-461. (in Chinese)
[15] 刘自刚, 孙万仓, 杨宁宁, 王月, 何丽, 赵彩霞, 史鹏飞, 杨刚, 李学才, 武军艳, 方彦, 曾秀存. 冬前低温胁迫下白菜型冬油菜抗寒性的形态及生理特征. 中国农业科学, 2013, 46(22): 4679-4687.
Liu Z G, Sun W C, Yang N N, Wang Y, He L, Zhao C X, Shi P F, Yang G, Li X C, Wu J Y, Fang Y, Zeng X C. Morphology and physiological characteristics of cultivars with different levels of cold-resistance in winter rapeseed (L.) during cold acclimation., 2013, 46(22): 4679-4687. (in Chinese)
[16] 史鹏辉, 孙万仓, 赵彩霞. 低温下抗氧化酶活性与冬油菜根细胞结冰关系的初步研究. 西北植物学报, 2013, 33(2): 329-335.
Shi P H, Sun W C, Zhao C X. Preliminary study on the relation of antioxidant enzyme activities at low temperature and the ice formation in root cells of winter rapeseed., 2013, 33(2): 329-335. (in Chinese)
[17] 刘自刚, 孙万仓, 方彦, 李学才, 杨宁宁, 武军艳, 曾秀存, 王月. 夜间低温对白菜型冬油菜光合机构的影响. 中国农业科学, 2015, 48(4): 672-682.
Liu Z G, Sun W C, Fang Y, Li X C, Yang N N, Wu J Y, Zeng X C, Wang Y. Effects of low nocturnal temperature on photosynthetic apparatus of winter rapeseed (L.)., 2015, 48(4): 672-682. (in Chinese)
[18] 孔德晶, 王月, 孙万仓, 曾秀存, 方彦. 鲁美宏, 杨宁宁. 北方白菜型冬油菜F2主要生理生化特性的变异与抗寒性相关分析. 草业学报, 2014, 23(4): 79-86.
Kong D J, Wang Y, Sun W C, Zeng X C, Fang Y, Lu M H, Yang N N. Analysis of variation in physio-biochemical characteristics and cold resistance in winter rapeseed F2populations., 2014, 23(4): 79-86. (in Chinese)
[19] 朱惠霞, 孙万仓, 邓斌, 燕妮, 武军艳, 范惠玲, 叶剑, 曾军, 刘雅丽, 张亚宏. 白菜型冬油菜品种的抗寒性及其生理生化特性. 西北农业学报, 2007, 16(4): 34-38.
Zhu H X, Sun W C, Deng B, Yan N, Wu J Y, Fan H L, Ye J, Zeng J, Liu Y L, Zhang Y H. Study on cold hardiness and its physiological and biochemical characteristics of winter turnip rape ()., 2007, 16(4): 34-38. (in Chinese)
[20] 蒲媛媛, 孙万仓. 白菜型冬油菜抗寒性与生理生化特性关系. 分子植物育种, 2010, 8(2): 335-339.
Pu Y Y, Sun W C. The Relationship between cold resistance of winter turnip rape varieties and its physiological characteristics., 2010, 8(2): 335-339. (in Chinese)
[21] 常硕其, 邓启云, 罗祎, 陈小龙. 超级杂交稻及其亲本的耐冷性研究. 杂交水稻, 2015, 30(1): 51-57.
Chang S Q, Deng Q Y, Luo Y, Chen X L. Studies on cold tolerance of super hybrid rice and its parents., 2015, 30(1): 51-57. (in Chinese)
[22] 贺荣华, 左佳, 李丁, 谢灵灵, 韩小霞, 高婧, 舒服, 李祺, 曹孟良. 水稻苗期抗寒性的杂种优势分析. 生命科学研究, 2011, 15(4): 356-358.
He R H, Zuo J, Li D, Xie L L, Han X X, Gao J, Shu F, Li Q, Cao M L. Analysis on the heterosis of cold tolerance at seedling stage in rice., 2011, 15(4): 356-358. (in Chinese)
[23] 裴玉贺, 王小丽, 张恩盈, 宋希云. 玉米抗寒生理指标的遗传效应分析. 植物生理学报, 2011, 47(3): 293-297.
Pei Y H, Wang X L, Zhang E Y, Song X Y. Genetic analysis of physiological indices related to cold resistance in maize., 2011, 47(3): 293-297. (in Chinese)
[24] 朱军. 数量性状的一般遗传模型和新的分析方法. 浙江农业大学学报, 1994, 20: 551-559.
Zhu J. General genetic models and new analysis methods for quantitative traits., 1994, 20: 551-559. (in Chinese)
[25] 邹奇. 植物生理学实验指导. 北京: 中国农业出版社, 2000.
Zou Q.. Beijing: China Agriculture Press, 2000. (in Chinese)
[26] 朱军. 遗传模型分析方法. 中国农业出版社, 1997.
Zhu J.. Beijing: China Agriculture Press, 1997. (in Chinese)
[27] 朱军, 季道藩, 许馥华. 作物品种间杂种优势遗传分析的新方法. 遗传学报, 1993, 20(3): 262-271.
Zhu J, Ji D F, Xu F H. A genetic approach for analyzing intra-cultivar heterosis in crops., 1993, 20(3): 262-271. (in Chinese)
[28] 郭海林, 高雅丹, 薛丹丹, 陈宣, 刘建秀. 结缕草属植物抗寒性的遗传分析. 草业学报, 2009, 18(3): 53.
Guo H L, Gao Y D, Xue D D, Chen X, Liu J X. Genetic analysis of cold tolerance of zoysia grass., 2009, 18(3): 53. (in Chinese)
[29] 王鹏良, 徐洋, 吕智鹏, 王海燕, 覃子海, 刘建秀, 王秀娥. 假俭草杂种F1抗寒性遗传分析. 草业学报, 2011, 20(2): 290-294.
Wang P L, Xu Y, Lü Z P, Wang H Y, Qin Z H, Liu J X, Wang X E. Genetic analysis for cold tolerance of F1hybrids in., 2011, 20(2): 290-294. (in Chinese)
[30] 张扬勇, 靳哲, 方智远, 刘玉梅, 杨丽梅, 庄木, 孙培田. 结球甘蓝抗寒性配合力分析及优良抗寒组合选育. 中国蔬菜, 2011(14): 23-27.
Zhang Y Y, Jin Z, Fang Z Y, Liu Y M, Yang L M, Zhuang M, Sun P T. Combining ability analysis of cold-resistance characteristic and elite combination breeding in cabbage (var.L.)., 2011(14): 23-27. (in Chinese)
[31] 吴吉祥, 王国建, 朱军, 许馥华, 季道藩. 陆地棉种子性状直接效应和母体效应的遗传分析. 作物学报, 1995, 21(6): 659-664.
Wu J X, Wang G J, Zhu J, Xu F H, Ji D F. Genetic analysis on direct and maternal effects of seed traits in upland cotton (L.)., 1995, 21(6): 659-664. (in Chinese)
[32] 李少钦, 王健胜, 张文学, 李殿荣, 郑磊, 田建华. 甘蓝型油菜优良亲本对杂种后代产量性状的遗传效应分析. 中国油料物学报, 2011, 33(6): 545-549.
Li S Q, Wang J S, Zhang W X, Li D R, Zheng L, Tian J H. Genetic analysis for hybrid yield traits using elite parents ofL.., 2011, 33(6): 545-549. (in Chinese)
(责任编辑 李莉)
Genetic Analysis of Traits related to Cold resistance in Winter Rapeseed (L.)
LIU Lin-bo1, SUN Wan-cang1, LIU Zi-gang1, WU Jun-yan1, FANG Yan1, LI Xue-cai1, ZENG Xiu-cun2, YANG Gang1, DONG yun3, CHEN Qi1, FANG Yuan1, YUAN Jin-hai1
(1Agronomy College, Gansu Agricultural University/Rapeseed Engineering Research Center of Gansu Province/Gansu Key Laboratory of Crop Improvement and Germplasm Enhancement/Key Laboratory of Arid Land Crop Science in Gansu Province ,Lanzhou 730070;2Hexi University, Zhangye 734000, Gansu;3Gansu Academy of Agricultural Sciences, Lanzhou 730070)
【Objective】The objective of this paper is to understand genetic way and to provide a theoretical foundation of cold resistance traits in winter rapeseed(L.). 【Method】In this study, six winter rapeseed cultivars with different cold resistances were used as parent materials to make complete diallel cross combinations with the Griffing methodⅠ. The parents and F1generations were planted to obtain the data of cold resistance traits. By using the additive-dominant model and MINQUE(1) method, the genetic variance, genetic correlation, parent additive effects, combination dominant effects and heritability, and heterosis of over wintering rate and 11 traits in winter rapeseed were analyzed.【Result】Over wintering rate, POD activity, SOD activity, SS content, relative electrolytic leakage, chlorophyll content, transpiration rate and stomatal conductance were mainly controlled by additive effects, and dominant effects were much less important. The dominant effects prevailed in the traits of CAT activity, MDA content, SP content and net photosynthetic rate. Over wintering rate was detected to be significantly and positively correlated with CAT activity, POD activity, SOD activity, SP content, SS content, chlorophyll content, transpiration rate, stomatal conductance and net photosynthetic rate, but negatively correlated with MDA content and relative electrolytic leakage. Longyou 7 and Longyou 8 had positively additive effects in the traits of overwintering rate, CAT activity, POD activity, SOD activity, SP content, SS content, chlorophyll content, transpiration rate, stomatal conductance and net photosynthetic rate, which indicated Longyou 7 and Longyou 8 could be used as parents that improved cold resistance of generations. The results of dominant effects analysis showed that Longyou 7×GY and Longyou 7×Longyou 8 had stronger cold resistance. As for the narrow heritabilities, SOD activity, relative electrolytic leakage, transpiration rate and overwintering rate were 74.63%, 59.06%, 57.49% and 56.87%, respectively, and CAT activity was the lowest (2.83%). CAT activity, SP content and net photosynthetic rate had stronger population mean heterosis and population over-parent heterosis. 【Conclusion】Overwintering rate, SOD activity, relative electrolytic leakage and transpiration rate were used in selection at early generation. Among the 6 elite breeding lines, Longyou 7 was recommended for using as direct parental line due to its higher general combining ability in cold resistance traits.
winter rapeseed; additive effect; dominant effect; cold resistance; heterosis
2016-06-27;接受日期:2016-08-26
国家自然科学基金(31460356, 31260334)、国家重点基础研究发展计划(973计划)(2011CB109300)、国家高技术研究发展计划(863计划)(2011AA10A104)、国家现代农业产业技术体系建设专项(CARS-13)、国家农业科技成果转化项目(2014G10000317)、甘肃省自然科学基金(145RJZG050)、油菜杂种优势利用技术与强优势杂交种创制(2016YFD0101300)
联系方式:刘林波,E-mail:444560733@qq.com。通信作者孙万仓,E-mail:18293121851@163.com
猜你喜欢
今日农业(2022年1期)2022-11-16 21:20:05
电脑迷(2022年10期)2022-11-08 02:41:30
今日农业(2021年20期)2021-11-26 01:23:56
河北果树(2020年4期)2020-11-26 06:04:28
西藏农业科技(2018年4期)2018-04-25 06:39:26
新疆农垦科技(2016年2期)2016-08-21 13:50:26
广西林业科学(2016年2期)2016-03-20 05:53:26
中国果菜(2016年9期)2016-03-01 01:28:40
中国马铃薯(2015年5期)2016-01-09 06:11:11
园艺与种苗(2015年10期)2015-02-27 11:27:51
