
燕麦β-葡聚糖对2型糖尿病大鼠肠黏膜屏障的影响
2016-11-12 06:21:04姜燕飞张召锋徐美虹北京大学公共卫生学院营养与食品卫生学系北京009北京大学营养与保健食品评价中心北京009
食品科学 2016年11期
刘 灿,姜燕飞,张召锋,徐美虹,李 勇,2,*(.北京大学公共卫生学院营养与食品卫生学系,北京 009;2.北京大学营养与保健食品评价中心,北京 009)
燕麦β-葡聚糖对2型糖尿病大鼠肠黏膜屏障的影响
刘 灿1,姜燕飞1,张召锋1,徐美虹1,李 勇1,2,*
(1.北京大学公共卫生学院营养与食品卫生学系,北京 100191;2.北京大学营养与保健食品评价中心,北京 100191)
目的:探究燕麦β-葡聚糖对2型糖尿病大鼠肠黏膜屏障的影响。方法:雄性SD大鼠高脂饲料喂养4 周,腹腔注射链脲佐菌素(streptozotocin,STZ)建立2型糖尿病模型,成模后随机分为模型对照组和燕麦β-葡聚糖低、中、高剂量组。低、中、高剂量组由基础饲料添加燕麦β-葡聚糖至其质量分数分别为0.275%、0.550%、1.100%,模型对照组和正常对照组均以基础饲料喂养。10 周后,测定其空腹血糖、肠道菌群以及结肠β-防御素-2、分泌型免疫球蛋白A(secretory immunoglobulin A,sIgA)、紧密连接蛋白Occludin含量。结果:高剂量组较模型对照组血糖水平显著降低(P<0.05);燕麦β-葡聚糖各干预组结肠紧密连接蛋白Occludin的表达水平显著高于模型对照组(P<0.05),β-防御素-2水平显著低于模型对照组(P<0.05),结肠sIgA水平与模型对照组之间的差异不显著(P>0.05);中、高剂量组结肠大肠杆菌数量上升幅度减小(P<0.05),燕麦β-葡聚糖各干预组双歧杆菌、乳酸杆菌数量上升幅度增大(P<0.05)。结论:燕麦β-葡聚糖可以改善2型糖尿病大鼠肠黏膜机械屏障和生物屏障的损伤,起到保护肠黏膜屏障的作用。
燕麦β-葡聚糖;2型糖尿病;肠黏膜屏障
肠黏膜屏障具有重要的防御作用,可以保护机体免受食物抗原、微生物及其有害代谢产物的损伤,保持机体内环境的相对稳定以维持正常生命活动[1]。由于肠道是机体内最大的细菌库和毒素库,肠黏膜屏障的损伤能够产生甚至加重多种疾病[2]。近年来已有研究证实,肠黏膜屏障受损而导致代谢性内毒素血症与2型糖尿病(type 2 diabetes mellitus,T2DM)的发生发展也有密切联系[3-4]。因此,维护肠黏膜屏障的完整对于2型糖尿病患者有着重要的意义。
燕麦β-葡聚糖是由β-(1→3)和β-(1→4)糖苷键连接β-D吡喃葡萄糖单位而形成的短链葡聚糖,是一种可溶性膳食纤维[5]。目前已有研究发现,燕麦β-葡聚糖对肠道健康有着重要的促进作用[6]。燕麦β-葡聚糖可以增加乳酸杆菌和双歧杆菌的数量及其活性[7],使大肠杆菌、梭状芽胞杆菌数量减少[8],具有调节肠道菌群的作用;其被酵解后的终产物二氧化碳、氢气、甲烷等气体能够刺激肠黏膜、促进肠蠕动,加快粪便的排出速度[9]。
但目前对于燕麦β-葡聚糖促进肠道健康的研究并未从肠黏膜屏障完整性的角度进行细致讨论,尚未见其对2型糖尿病肠黏膜屏障的影响的有关报道。本实验以链脲佐菌素(streptozotocin,STZ)注射联合高脂饲料喂养诱导2型糖尿病大鼠模型,成模后采用含燕麦β-葡聚糖的燕麦饲料进行干预,观察其肠黏膜机械屏障、化学屏障、免疫屏障及生物屏障的变化,以研究燕麦β-葡聚糖对2型糖尿病大鼠肠黏膜屏障的影响,从而为2型糖尿病的预防和治疗提供实验依据。
1 材料与方法
1.1 动物、材料与试剂
80 只无特定病原体(specific pathogen free,SPF)级雄性Sprague-Dawley(SD)大鼠(动物合格证号:SCXK(京)2012-0001;动物使用许可证号:SYXK(京)2011-0039),6 周龄,体质量180~200 g,购自北京维通利华实验动物技术有限公司。饲养于北京大学医学部实验动物科学部,室温恒定21~25 ℃,相对湿度40%~50%,节律光照。大鼠每2 只为1笼进行饲养,自由采食与饮水。
高脂饲料配方:基础饲料66%(质量分数,下同)、猪油15%、蔗糖10%、酪蛋白6%、蛋黄粉3%;燕麦饲料:燕麦β-葡聚糖(纯度95%,燕麦提取物β-葡聚糖(HPLC级))添加入基础饲料中,使其燕麦β-葡聚糖质量分数分别为0.275% 、0.550%、1.100%。以上饲料均购自北京科澳协力饲料有限公司,具体成分如表1所示。

表1 实验动物饲料配方Table 1 Ingredients and formulations of experimental diets
链脲佐菌素(streptozotocin,STZ) 美国Sigma公司;柠檬酸钠、柠檬酸、磷酸盐缓冲液(phosphate buffer solution,PBS) 北京化工厂;大鼠分泌型免疫球蛋白A(secretory immunoglobulin A,sIgA)、β-防御素-2酶联免疫吸附法试剂盒、免疫组化试剂盒 北京中杉金桥生物技术有限公司;细胞组织快速裂解液、二喹啉甲酸(bicinchonininc acid,BCA)蛋白浓度测定试剂盒、酶放大化学发光免疫分析(electro-chemi-luminescence,ECL)试剂盒 上海碧云天生物技术有限公司;脱脂奶粉 美国BD公司;兔抗大鼠紧密连接蛋白Occludin多克隆抗体、小鼠抗β-肌动蛋白多克隆抗体 英国Abcam公司;乳酸杆菌选择性(Lactobacillus selective,LBS)琼脂培养基、双歧杆菌选择性(Bifidobacterium selective,BS)培养基 青岛高科园海博生物技术有限公司;伊红-美兰琼脂培养基 北京奥博星生物技术有限责任公司。
1.2 仪器与设备
快速血糖仪 美国强生公司;Bio-Rad550型酶联免疫检测仪、蛋白质电转移装置 美国Bio-Rad公司;UV-762型分光光度计 上海精密科学仪器有限公司;垂直板蛋白电泳仪 北京六一仪器厂;LX-C35L型立式自动电热压力蒸汽灭菌器 合肥华泰医疗设备有限公司;HPX-9052 MBE电热恒温培养箱 上海博迅实业有限公司医疗设备厂;生物显微镜 日本尼康公司。
1.3 方法
1.3.1 2型糖尿病大鼠模型的建立
采用低剂量STZ腹腔注射联合高脂饮食喂养,即通过破坏部分胰岛β细胞、诱导胰岛素抵抗,建立2型糖尿病大鼠模型。将80 只大鼠用基础饲料适应性喂养1 周后,随机选取68 只经高脂饲料喂养4 周,腹腔注射STZ(0.01 mol/L pH 4.5柠檬酸钠-柠檬酸缓冲液配制)25 mg/kg(以体质量计)每周1 次,共注射2 次。每周尾静脉采血检测血糖水平,2 周后空腹血糖达到11.1 mmol/L以上并稳定7 d者确定为2型糖尿病模型大鼠。
1.3.2 动物分组及干预
12 只基础饲料喂养的正常大鼠设为正常对照组。取48 只造模成功的大鼠随机分为4 组,每组12 只,包括:糖尿病模型(diabetic model,DM)对照组及燕麦β-葡聚糖低剂量(low dose oat β-glucan,OL)、中剂量(medium dose oat β-glucan,OM)、高剂量(high dose oat β-glucan,OH)干预组。其中正常对照组、糖尿病模型对照组均采用基础饲料,OL、OM、OH组分别采用相应剂量的燕麦饲料。将开始干预记为第0周,连续饲养10 周。
1.3.3 标本采集
实验期间,每周观察各组大鼠的一般情况,包括毛色、精神状态、摄食、饮水、尿量、体质量及日常活动等。在第0、10周取新鲜粪便样品进行细菌平板培养。第10周经股动脉采血后处死全部大鼠,立即于距回盲部3 cm处取结肠标本,分别进行组织匀浆测sIgA、β-防御素-2含量,研磨提取蛋白测紧密连接蛋白Occludin,以及用中性缓冲福尔马林溶液固定、常规脱水、石蜡包埋并切片。
1.3.4 指标测定
1.3.4.1 血糖含量
各组大鼠禁食不禁水12 h,于第0、10周分别采用血糖仪和血糖试纸,剪尾取血测定各组大鼠空腹血糖水平。
1.3.4.2 结肠紧密连接蛋白Occludin含量
免疫组织化学法:参照免疫组化试剂盒说明书将结肠石蜡切片进行免疫组织化学染色。显微镜下观察并拍照,呈棕黄色染色为阳性。
蛋白质印迹法:取结肠组织加入细胞组织快速裂解液于冰上充分碾碎匀浆,离心提取总蛋白,采用BCA蛋白浓度测定试剂盒测定样本总蛋白浓度。将样本加上样缓冲液后,99 ℃变性。上样40 μg样本总蛋白,采用质量分数10%的分离胶进行蛋白分离,电泳初始电压为80 V。电泳后进行湿式电转,200 mA恒流转膜1.5 h。质量分数5%的脱脂牛奶室温封闭2 h。一抗紧密连接蛋白Occludin(1∶1000,V/V)、内参β-肌动蛋白(1∶5 000,V/V)4 ℃孵育过夜,体积分数0.1%三羟甲基氨基甲烷-吐温缓冲液(tris-buffered saline with tween,TBST)洗3 次,每次10 min。二抗(1∶4 000,V/V)室温孵育2 h,体积分数0.1% TBST洗3 次,每次10 min。使用ECL发光试剂盒进行发光,显影定影后检测结果经拍照后获得图像。应用Image-Pro Plus图像分析系统对图片进行半定量研究,分别测定各组待测蛋白与内参照蛋白的灰度值,计算各组两者之间的比值。
1.3.4.3 结肠防御性物质
用预冷的PBS冲洗结肠组织,将组织剪碎后与PBS混合于冰上充分研磨匀浆,离心取上清,采用酶联免疫吸附法(enzyme linked immunosorbent assay,ELISA)并参照相应的试剂盒说明书分别检测结肠β-防御素-2、sIgA的水平。
1.3.4.4 肠道菌群
第10周在无菌条件下取新鲜粪便0.1 g,加入10 mL灭菌生理盐水,振荡混匀,初始质量浓度为1×10-2g/mL,依次10 倍递增稀释成一系列不同稀 释度的含菌液至1×10-7g/mL。选择适宜的稀释度在超净台内进行接种,每个质量浓度梯度做2~3 个平行样。选择性培养基提前煮制、分装并灭菌。细菌平板培养条件:1)乳酸杆菌:LBS琼脂培养基,37 ℃恒温厌氧条件下培养48 h;2)双歧杆菌:BS培养基,37 ℃恒温厌氧条件下培养48 h;3)大肠杆菌:伊红-美蓝琼脂培养基,37 ℃恒温下有氧培养24 h。分别于培养结束时进行形态和生化实验以鉴定菌属、挑选特征性菌落,进行菌落计数,计算每克粪便的菌落数,以菌落形成单位(colony forming unit,CFU)表示,公式如下。
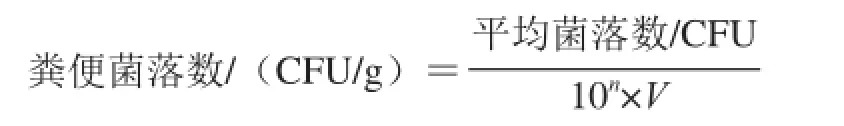
式中:10n为该菌落数对应点样质量浓 度/(g/mL);V为对应点样体积/mL。
将结果按以10为底的对数形式转换lg(CFU/g)后进行统计分析。
1.4 数据处理
2 结果与分析
2.1 大鼠一般情况
在整个实验过程中,正常对照组大鼠生长、进食及活动情况正常,毛色白而光泽,精神状态良好;模型对照组及燕麦β-葡聚糖干预组大鼠均出现多饮、多食、多尿现象,活动减少,毛色脏乱无光泽现象。
造模前(即干预前第8周),各组的基线体质量之间差异不显著(P>0.05)。截至造模完成时(即干预第0周),正常对照组的体质量已高于各模型组的体质量水平,差异显著(P<0.05),各模型组间的体质量差异不显著(P>0.05)。如图1所示,在整个干预期间,正常对照组大鼠体质量稳定增长;DM、OL、OM、OH组体质量呈下降趋势,其各组之间体质量的差异不显著(P>0.05),但均明显低于正常对照组的体质量(P<0.05)。

图1 各组体质量随干预时间的变化Fig. 1 Change in body weight of rats
2.2 大鼠血糖水平

表2 干预期间各组大鼠空腹血糖水平(n==1122)Table 2 Fasting plasma glucose levels of rats during the administration of ooaatt β-gluccaann ((n = 1122))
由表2可知,干预第0周,DM组与燕麦β-葡聚糖各干预组(OL、OM、OH组)大鼠血糖之间的差异不显著(P>0.05),但均高于正常对照组(P<0.05),造模成功。干预第10周,OL、OM、OH组大鼠血糖水平呈下降趋势,DM组呈升高趋势,其中OH组与DM组大鼠血糖之间的变化差异显著(P<0.05)。
2.3 大鼠结肠紧密连接蛋白Occludin表达水平
由图2可知,正常对照组大鼠结肠黏膜中紧密连接蛋白Occludin分布位于肠上皮细胞顶端,染色成点状聚积,蜂巢状分布,在整张切片上均可见到表达很强的棕褐色信号(图2A);模型对照组显示棕褐色阳性信号明显减弱,阳性细胞数减少,分布不均(图2B);燕麦β-葡聚糖各干预组棕褐色阳性信号较DM组增强,阳性细胞数增加,其中OH组的阳性表达最强,但均弱于正常对照组(图2C、D、E)。
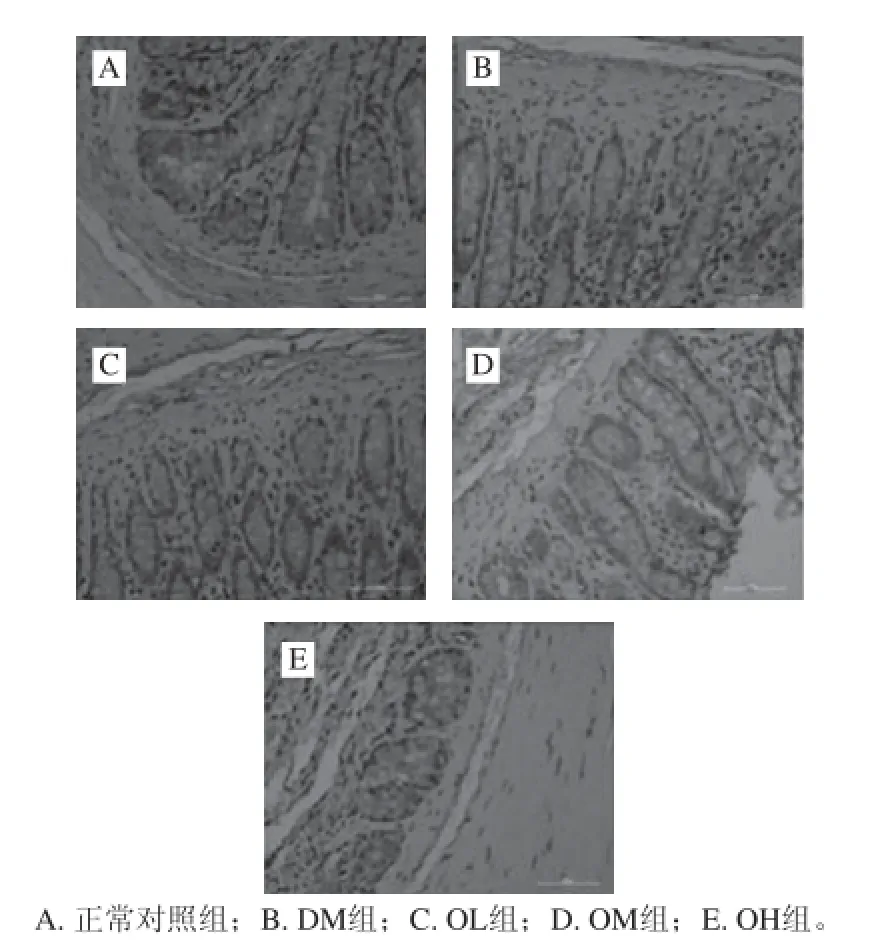
图2 结肠紧密连接蛋白Occludin免疫组化染色和苏木精染核(×400)Fig. 2 Immunohisochemical staining of Occludin in the colon of rats and hematoxylin staining of cell nuclei (× 400)
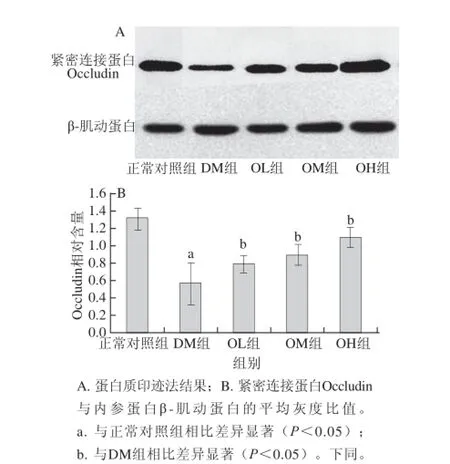
图33 燕麦β-葡聚糖对糖尿病大鼠结肠紧密连接蛋白Occludiinn的影响(±s,n=1122)Fig. 3 Effect of oat β-glucan on the expression of Occludin in the colon of diabetic rats (± s, n=12)
由图3可知,DM组大鼠结肠中紧密连接蛋白Occludin平均灰度值与内参蛋白β-肌动蛋白的比值为0.57±0.24,明显低于正常对照组的1.31±0.13,其表达水平差异显著(P<0.05);与DM组相比,OL组、OM组、OH组大鼠结肠紧密连接蛋白Occludin的表达水平较高,差异显著(P<0.05)。
2.4 大鼠结肠β-防御素-2水平
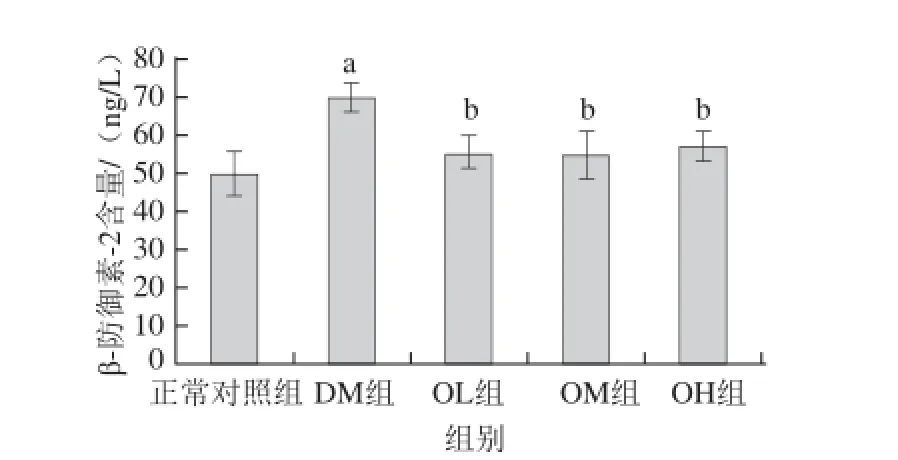
图44 燕麦β-葡聚糖对糖尿病大鼠结肠组织中β-防御素--22的影响(±s,n=1122)Fig. 4 Effect of oat β-glucan on β-defesin-2 level in the colon of diabetic rats (± s, n=12)
由图4可知,DM组大鼠结肠β-防御素-2表达水平为(69.5±3.8) ng/L,明显高于正常对照组的(49.6±5.6) ng/L,差异显著(P<0.05);与DM组相比,OL、OM、OH组大鼠结肠β-防御素-2水平较低,其差异显著(P<0.05)。
2.5 大鼠结肠sIgA水平
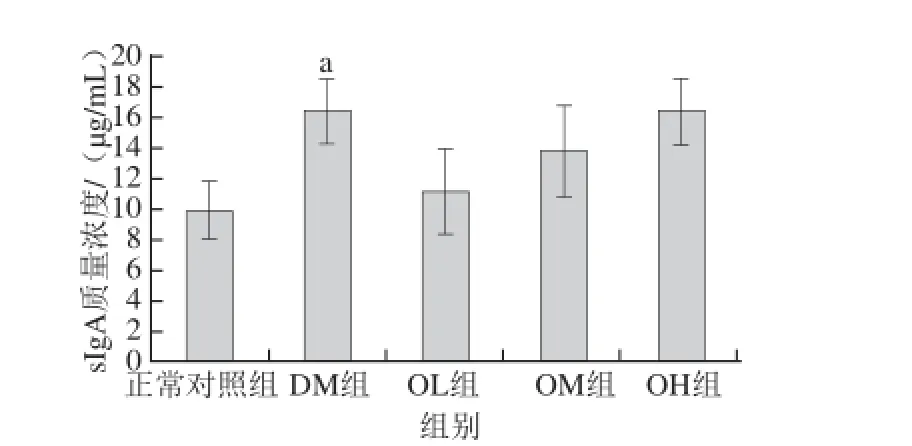
图5 燕麦β-葡聚糖对2型糖尿病大鼠结肠组织sIggAA的影响Fig. 5 Effect of oat β-glucan on sIgA levels in the colon of diabetic ra ts
由图5可知,DM组大鼠结肠组织中的sIgA质量浓度为(16.6±2.0)μg/mL,远高于正常对照组的(9.9±1.9) μg/mL,其差异显著(P<0.05);与DM组相比,OL、OM、OH组大鼠结肠组织中的sIgA水平的差异均不显著(P>0.05)。
2.6 大鼠肠道菌群平板培养
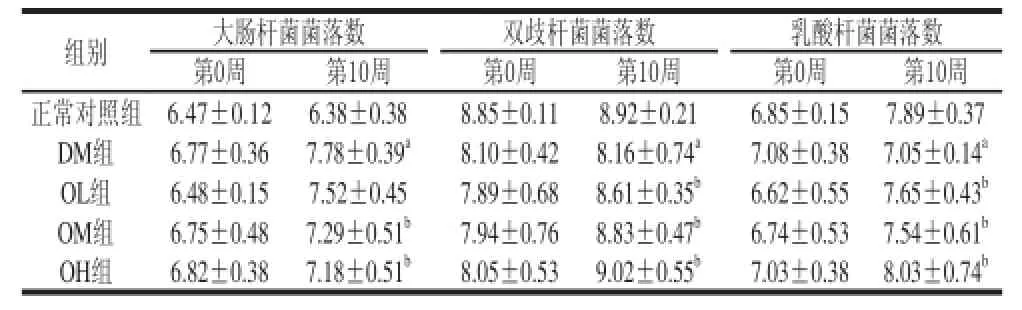
表3 各组大鼠肠道细菌平板计数结果Table 3 Change in intestinal bacteria of rats during administration of oat β-glucan lg(CFU/g)
由表3可知,第0周,正常对照组与DM组大鼠大肠杆菌、双歧杆菌、乳酸菌菌落数之间的差异不显著(P>0.05),DM组与OL、OM、OL组以上菌群菌落数之间的差异不显著(P>0.05)。第10周,正常对照组与DM组大鼠大肠杆菌、双歧杆菌、乳酸菌菌落数之间的差异显著(P<0.05)。对于大肠杆菌,DM组和燕麦β-葡聚糖各干预组大肠杆菌菌落数均呈上升趋势,但燕麦β-葡聚糖各干预组上升幅度较小,其中OM、OH组与DM组大鼠大肠杆菌菌落数上升幅度之间的差异显著(P<0.05);对于双歧杆菌,DM组和燕麦β-葡聚糖各干预组双歧杆菌菌落数均呈上升趋势,但燕麦β-葡聚糖各干预组上升幅度较大,其中OL、OM、OH组与DM组大鼠双歧杆菌菌落数上升幅度之间的差异显著(P<0.05),但未检验到剂量反应关系;对于乳酸杆菌,DM组乳酸杆菌菌落数呈下降趋势,但燕麦β-葡聚糖各干预组均呈上升趋势,其中OL、OM、OH组与DM组大鼠乳酸杆菌菌落数上升幅度之间的差异显著(P<0.05),但未检验到剂量反应关系。
3 讨 论
本研究参考前期研究结果[10-11],根据燕麦β-葡聚糖在人群中表现出明显改善作用的推荐剂量,设置OL、OM、OH 3 个剂量组对成模后的糖尿病大鼠进行干预。通过检测大鼠结肠紧密连接蛋白Occludin表达水平、β-防御素-2水平、sIgA水平、肠道菌群菌落数的变化,讨论了其对肠黏膜的机械屏障、化学屏障、免疫屏障及生物屏障影响,为进一步研究燕麦β-葡聚糖对2型糖尿病的作用及其机制提供依据。
紧密连接蛋白Occludin是紧密连接中的重要结构蛋白,能够调节肠黏膜细胞间黏附、移动及通透性,其存在和数量决定肠道选择性屏障,是肠黏膜机械屏障功能的重要指标[12-13]。紧密连接蛋白Occludin一旦发生变异、减少和缺失可引起肠上皮细胞间隙通透性增加,对其表达水平的检测能反映肠道紧密连接及屏障破坏的情况[14]。本研究结果显示,DM组大鼠的紧密连接蛋白Occludin在结肠黏膜分布不均匀,其平均灰度值与内参蛋白β-肌动蛋白的比值为0.57±0.24,低于正常对照组的1.31±0.13(P<0.05),说明DM组大鼠紧密连接蛋白Occludin的表达水 平较正常对照组低,提示2型糖尿病大鼠肠黏膜机械屏障功能降低。而在燕麦β-葡聚糖各干预组中,紧密连接蛋白Occludin表达水平均高于DM组(P<0.05),提示燕麦β-葡聚糖具有一定的改善肠黏膜机械屏障损伤的作用。
β-防御素-2由肠上皮细胞分泌的一种抗菌肽,具有广谱的抗微生物活性,对其水平的检测能一定程度上反映肠道化学屏障的功能状况[15-16]。β-防御素-2表达水平的升高与肠道的炎症过程有关[17],细菌、真菌、脂多糖等的刺激可明显诱导或增强β-防御素-2的表达[16,18]。本研究显示,模型对照组大鼠的结肠组织β-防御素-2水平为(69.5±3.8) ng/L,明显高于正常对照组的(49.6±5.6) ng/L(P<0.05),提示2型糖尿病大鼠肠黏膜免疫功能失衡。而燕麦β-葡聚糖的干预则使其水平下降(P<0.05),从而有助于减轻结肠黏膜的炎症反应,避免过强的免疫反应,提示燕麦β-葡聚糖具有一定的调节肠黏膜化学屏障的作用。
对于肠黏膜免疫屏障的主要抗体sIgA,本研究结果显示,DM组大鼠的结肠组织sIgA抗体分泌增加,达(16.6±2.0) μg/mL,远高于正常对照组的(9.9±1.9) μg/mL(P<0.05)。但本研究未发现燕麦β-葡聚糖干预对2型糖尿病大鼠结肠组织的sIgA抗体水平的影响。
肠道内优势菌群结构的动态平衡及乳酸杆菌、双歧杆菌等专性厌氧菌构成了肠道的生物屏障[19]。本研究发现,DM组鼠大肠杆菌、乳酸杆菌菌落数高于正常对照组(P<0.05),双歧杆菌菌落数低于正常对照组(P<0.05),而燕麦β-葡聚糖可以使2型糖尿病大鼠大肠杆菌菌落数增加幅度下降(P<0.05),使双歧杆菌菌落数增加幅度上升(P<0.05),并且使乳酸杆菌菌落数增加(P<0.05),虽并未发现剂量反应关系,但这也一定程度上提示燕麦β-葡聚糖具有降低有害菌、增加有益菌数量、调节肠道菌群的作用。近年来也有学者观察到,燕麦β-葡聚糖能够增加乳酸杆菌和双歧杆菌菌落数[20-21],减少肠杆菌菌落数,并且具有剂量反应关系[22],这都提示燕麦β-葡聚糖具有一定的改善肠黏膜生物屏障损伤的作用。
肠黏膜屏障损伤与2型糖尿病发生发展有着密切的联系。有研究发现,当肠黏膜屏障受到损伤时,内毒素的主要来源脂多糖可突破肠黏膜进入循环系统,与单核巨噬细胞的CD14形成复合物,被免疫细胞表面的Toll样受体4识别,通过髓样分化分子88激活核转录因子-κB,促进肿瘤坏死因子α、干扰素γ、白细胞介素6等多种炎症因子的合成和分泌,造成机体慢性低度炎性反应,从而对胰岛素抵抗及2型糖尿病等代谢性疾病的发生、发展产生影响[23-24]。本研究发现燕麦β-葡聚糖对于维持肠黏膜屏障结构和功能的完整性具有一定的作用,可以改善肠黏膜机械屏障和生物屏障的损伤,为进一步研究燕麦β-葡聚糖对2型糖尿病的作用提供了依据。由于本研究干预时间较短(10 周),这对于实验结果有一定的影响,建议今后开展长期的燕麦β-葡聚糖干预研究,以获得更为准确、稳定的结果。
[1] ANASTASILAKIS C D, IOANNIDIS O, GKIOMISI A I, et al. Artificial nutrition and intestinal mucosal barrier functionality[J]. Digestion, 2013, 88(3): 193-208. DOI:10.1159/000353603.
[2] SALVO R E, ALONSO C C, PARDO C C, et al. The intestinal barrier function and its involvement in digestive disease[J]. Revista Espanola de Enfermedades Digestivas, 2015, 107(11): 686-696. DOI:10.17235/ reed.2015.3846/2015.
[3] CANI P D, AMAR J, IGLESIAS M A, et al. Metabolic endotoxemia initiates obesity and insulin resistance[J]. Diabetes, 2007, 56(7): 1761-1772. DOI:10.2337/db06-1491.
[4] CANI P D, BIBILONI R, KNAUF C, et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat dietinduced obesity and d iabetes in mice[J]. Diabetes, 2008, 57(6): 1470-1481. DOI:10.2337/db07-1403.
[5] ROUBROEKS J P, ANDERSSON R, MASTROMAURO D I, et al. Molecular weight, structure and shape of oat (1→3),(1→4)-β-D-glucan fractions obtained by enzymatic degradation with (1→4)-β-D-glucan 4-glucanohydrolase from Trichoderma reesei[J]. Carbohydrate Polymers, 2001, 46(3): 275-285. DOI:10.1016/S0144-8617(00)00329-5.
[6] THIES F, MASSON L F, BOFFETTA P, et al. Oats and bowel disease: a systematic literature review[J]. British Journal of Nutrition, 2014, 112(Suppl 2): 31-43. DOI:10.1017/S0007114514002293.
[7] METZLER-ZEBELI B U, ZIJLSTRA R T, MOSENTHIN R, et al. Dietary calcium phosphate content and oat beta-glucan influence gastrointestinal microbiota, butyrate-producing bacteria and butyrate fermentation in weaned pigs[J]. FEMS Microbiology Ecology, 2011, 75(3): 402-413. DOI:10.1111/j.1574-6941.2010.01017.x.
[8] KEDIA G, VAZQUEZ J A, CHARALAMPOPOULOS D, et al. In vitro fermentation of oat bran obtained by debranning with a mixed culture of human fecal bacteria[J]. Current Microbiology, 2009, 58(4): 338-342. DOI:10.1007/s00284-008-9335-1.
[9] 汪海波, 刘大川, 汪海婴, 等. 燕麦β-葡聚糖对小肠蠕动及淀粉酶活性的影响研究[J]. 营养学报, 2006, 28(2): 148-151. DOI:10.3321/ j.issn:0512-7955.2006.02.023.
[10] MA X, GU J, ZHANG Z, et al. Effects of Avena nuda L. on metabolic control and cardiovascular disease risk among Chinese patients with diabetes and meeting metabolic syndrome criteria: secondary analysis of a randomized clini cal trial[J]. European Journal of Clinical Nutrition, 2013, 67(12): 1291-1297. DOI:10.1038/ejcn.2013.201.
[11] 井路路, 张召锋, 顾娇娇, 等. 优化全胚芽裸燕麦膳食模式对T2DM伴胆固醇异常患者影[J]. 中国公共卫生, 2013, 29(11): 1587-1591. DOI:10.11847/zgggws2013-29-11-09.
[12] ASSIMAKOPOULOS S F, PAPAGEORGIOU I, CHARONIS A. Enterocytes’ tight junctions: from molecules to diseases[J]. World Journal of Gastrointestinal Pathophysiology, 2011, 2(6): 123-137. DOI:10.4291/wjgp.v2.i6.123.
[13] SUZUKI T. Regulation of intestinal epithelial permeability by tight junctions[J]. Cellular and Molecular Life Sciences, 2013, 70(4): 631-659. DOI:10.1007/s00018-012-1070-x.
[14] AL-SADI R, KHATIB K, GUO S, et al. Occludin regulates macromolecule flux across the intestinal epithel ial tight junction barrier[J]. AJP: gastrointestinal and Liver Physiology, 2011, 300(6): G1054-G1064. DOI:10.1152/ajpgi.00055.2011.
[15] METZ-BOUTIGUE M H, SHOOSHTARIZADEH P, PREVOST G, et al. Antimicrobial peptides present in mammalian skin and gut are multifunctional defence molecules[J]. Current Pharmaceutical Design, 2010, 16(9): 1024-1039. DOI:10.2174/138161210790963823.
[16] HILL D R, KESSLER S P, RHO H K, et al. Specifi c-sized hyaluronan fragments promote expression of human beta-defensin 2 in intestinal epithelium[J]. Journal of Biological Chemistry, 2012, 287(36): 30610-30624. DOI:10.1074/jbc.M112.356238.
[17] WEHKAMP J, FELLERMANN K, HERRLINGER K R, et al. Human beta-defensin 2 but not beta-defensin 1 is expressed preferentially in colonic mucosa of inflammatory bowel disease[J]. European Journal of Gastroenterology and Hepatology, 2002, 14(7): 745-752. DOI:10.1097/00042737-20 0207000-00006.
[18] YANG D, LIU Z H, TEWARY P, et al. Defensin participation in innate and adaptive immunity[J]. Current Pharmaceutical Design, 2007, 13(30): 3131-3139. DOI:10.2174/138161207782110453.
[19] TANABE S, SUZUKI T, WASANO Y, et al. Anti-inflammatory and intestinal barrier-protective activities of commensal lactobacilli and bifidobacteria in thoroughbreds: role of probiotics in diarrhea prevention in neonatal thoroughbreds[J]. Journal of Equine Science, 2014, 25(2): 37-43. DOI: 10.1294/jes.25.37.
[20] ZHOU A L, HERGERT N, ROMPATO G, et al. Whole grain oats improve insulin sensitivity and plasma cholesterol profi le and modify gut microbiota composition in C57BL/6J mice[J]. Journal of Nutri tion, 2015, 145(2): 222-230. DOI:10.3945/jn.114.199778.
[21] CONNOLLY M L, TUOHY K M, LOVEGROVE J A. Wholegrain oat-based cereals have prebiotic potential and low glycaemic index[J]. British Journal of Nutrition, 2012, 108(12): 2198-2206. DOI:10.1017/ S0007114512000281.
[22] SHEN R L, DANG X Y, DONG J L, et al. Effects of oat beta-glucan and barley beta-glucan on fecal characteristics, intestinal microfl ora, and intestinal bacterial metabolites in rats[J]. Journal of Agricultural and Food Chemistry, 2012, 60(45): 11301-11308. DOI:10.1021/ jf302824h.
[23] CANI P D, OSTO M, GEURTS L, et al. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity[J]. Gut Microbes, 2012, 3(4): 279-288. DOI:10.4161/gmic.19625.
[24] HAMEED I, MASOODI S R, MIR S A, et al. Type 2 diabetes mellitus: from a metabolic disorder to an infl ammatory condition[J]. W orld Journal of Diabetes, 2015, 6(4): 598-612. DOI:10.4239/wjd. v6.i4.598.
Effect of Oat β-Glucan on Intestinal Mucosal Barrier in Type 2 Diabetic Rats
LIU Can1, JIANG Yanfei1, ZHANG Zhaofeng1, XU Meihong1, LI Yong1,2,*
(1. Department of Nutrition and Food Hygiene, School of Public Health, Peking University, Beijing 100191, China; 2. Nutritional and Functional Food Assessment Center, Peking University, Beijing 100191, China)
Objective: To investigate the effect of oat β-glucan on intestinal mucosal barrier in type 2 diabetic rats. Methods: Totally 60 male SD rats were randomly divided into fi ve groups including normal control group and four diabetic model groups. Type 2 diabetic rat models were established by high-fat diet feeding and low-dose intraperitoneal injection of streptozotocin (STZ, 0.01 mol/L, 25 mg/(kg·d)). The diabetic rats were randomly divided into model control group and low, medium and high dose oat β-glucan treatment group. Oat β-glucan was administrated by lavage. Low (0.275%), med ium (0.550%) and high (1.100%) doses of oat β-glucan added to the basal diet were given to each intervention group, respectively. The rats in the model control and normal control groups were fed on the basal diet. After 10 weeks, fasting plasma glucose, intestinal flora and colon β-defensin-2, secretory immunoglobulin A (sIgA) and occludin level were measured. Results: Fasting plasma glucose of the high dose oat β-glucan treatment group was lower than that of the model control group (P < 0.05). Compared with model control group, the expression level of the colonic tight junction protein occludin was higher (P < 0.05), and β-defense-2 levels were lower (P < 0.05); yet no difference was found in sIgA levels (P > 0.05) in oat β-glucan treatment groups. The increase in Escherichia coli counts in the medium and high dose oat β-glucan treatment groups were smaller than that in the model control group (P < 0.05). The increase in bifi dobacteria and lactobacilli counts of oat β-glucan treatment groups were larger than that in the model control group (P < 0.05). Conclusion: Oat β-glucan has a positive effect on intestinal mucosal mechanical barrier and biological barrier injury in type 2 diabetic rats.
oat β-glucan; type 2 diabetes mellitus; intestinal mucosal barrier
10.7506/spkx1002-6630-201611029
R151.1
A
1002-6630(2016)11-0167-07
刘灿, 姜燕飞, 张召锋, 等. 燕麦β-葡聚糖对2型糖尿病大鼠肠黏膜屏障的影响[J]. 食品科学, 2016, 37(11): 167-173.
DOI:10.7506/spkx1002-6630-201611029. http://www.spkx.net.cn
LIU Can, JIANG Yanfei, ZHANG Zhaofeng, et al. Effect of oat β-glucan on intestinal mucosal barrier in type 2 diabetic rats[J]. Food Science, 2016, 37(11): 167-173. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201611029. http://www.spkx.net.cn
2015-09-27
国家自然科学基金青年科学基金项目(81402665);北京市自然科学基金青年科学基金项目(7154212)
刘灿(1990—),女,硕士研究生,研究方向为营养与疾病。E-mail:Catherine_Liu@126.com
*通信作者:李勇(1958—),男,教授,博士,研究方向为营养与疾病。E-mail:liyong@bjmu.edu.cn
猜你喜欢
中老年保健(2022年5期)2022-11-25 14:16:14
江苏安全生产(2022年8期)2022-11-01 09:14:32
现代畜牧科技(2021年10期)2021-11-19 08:42:24
小资CHIC!ELEGANCE(2021年36期)2021-10-15 14:36:34
今日农业(2021年7期)2021-07-28 07:07:24
艺术评鉴(2020年5期)2020-04-30 06:47:57
人大建设(2018年10期)2018-12-07 01:13:46
延河(2017年7期)2017-07-19 21:01:10
现代农业(2016年6期)2016-02-28 18:42:51
食品工业科技(2014年15期)2014-03-11 18:17:20
