
阿尔茨海默病患者血清中microRNAs表达谱的研究
2016-11-12 01:19:31曾庆宏聂红霞于善花姜建东王传淇彭艳艳张浩江庄爱霞
中风与神经疾病杂志 2016年2期
曾庆宏,刘 霞,周 芳,聂红霞,于善花,姜建东,王传淇,彭艳艳,张浩江,庄爱霞
阿尔茨海默病患者血清中microRNAs表达谱的研究
曾庆宏1,刘霞2,周芳1,聂红霞1,于善花1,姜建东1,王传淇1,彭艳艳1,张浩江1,庄爱霞1
目的探讨阿尔茨海默病(AD)患者血清中microRNAs与阿尔茨海默病之间的关系,寻求一种可能作为辅助诊断AD的早期生物标志物。方法研究对象分为AD组和对照组,分别取其血清并提取microRNAs,利用microRNA基因芯片检测出表达差异的microRNAs,并用实时荧光定量PCR(QT-PCR)的方法进行验证。 结果基因芯片筛选出43个与AD相关的差异表达microRNAs(P<0.05),其中microRNA-107及microRNA-222的表达水平在AD患者中均显著低于对照组(P<0.01),与QT-PCR验证结果一致。结论MicroRNA-107及microRNA-222可能通过多种途径影响AD的发生发展,有望成为一种辅助诊断AD的早期生物标志物。
阿尔茨海默病;microRNAs;microRNA-107;microRNA-222
阿尔茨海默病(Alzheimer’s disease,AD)是一种神经退行性疾病,表现为认知、语言、记忆等功能障碍[1,2],日常生活能力进行性减退,并可伴有各种神经精神症状和行为障碍,是导致老年人痴呆的主要原因。近年来一些新技术的开发应用,促进了蛋白组学的研究,基质辅助激光解析电离-飞行时间质谱(MALDI-TOF/TOF MSS)[3],高效液相色谱与电感藕合等离子质谱(HPLC-ICP-MS)[4],液相色谱-紫外光谱-质谱联用技术(HPLCUV-MS)[5,6],鸟枪法蛋白质组学[7]等对蛋白的分离、定性和定量的检测,可能有助于新的AD生物标记物的发现,但仍缺乏特异性和敏感性均高的单一的诊断方法。
microRNA是真核生物中一类长度约为22个核苷酸的参与基因转录后水平调控的单链非编码小分子RNA,主要功能是通过抑制靶mRNA翻译或促使其降解的方式对基因表达起调控作用[8],参与细胞周期调控及个体发育过程[9]。在神经系统中microRNA调节不同发育阶段及不同部位的病生理条件下神经细胞的增殖、分化与凋亡,并对人类的认知和记忆能力的形成起重要作用[10,11]。大量研究表明,血清中存在着稳定的microRNAs表达[12,13]。本研究利用μParaflo微流体芯片技术检测了AD患者与健康体检者血清中的microRNAs,并用QT-PCR方法进行验证,发现了与AD密切相关的microRNAs,有望成为AD的生物学诊断和判断预后的重要指标。
1 材料与方法
1.1研究对象(1)AD组:选取2013年6月-2015年6月在我院住院及门诊就诊首次诊断AD的患者30例;(2)设年龄相匹配的对照组:为正常健康体检者30例,具有正常功能活动能力,无器质性脑病,神经系统检查无异常,MMSE评分在28分以上。排除标准:(1)既往有脑血管疾病者;(2)有颅脑外伤者;(3)有中毒性、代谢性等脑病者;(4)诊断前进行药物治疗者;(5) 有血液系统疾病者;(6)血管性及其他原因所致痴呆;(7)未知情同意者。
1.2诊断标准阿尔茨海默病诊断标准根据2011年美国国家衰老研究所(National Institute of Aging,NIA)和阿尔茨海默病学会(Alzheimer’s Association,AA)制定的诊断标准(NIA-AA诊断标准)。简易智能精神状态量表(MMSE)评分标准:文盲组≤17分,小学组≤20分,中学组≤22分,大学组≤24分。
1.3方法
1.3.1资料收集记录所有研究对象的性别、年龄、既往史(高血压、糖尿病、冠心病等)、吸烟史、饮酒史以及纤维蛋白原、高半胱氨酸、C-反应蛋白、血脂等指标。
1.3.2样本采集AD组治疗前采取静脉血5 ml,对照组静脉血来自体检中心体检者,所有静脉血提取血清后放入-80 ℃冰箱保存备用。
1.3.3总RNA提取取100~200 μl低温冷藏的每例血清样本,置于冰上融化,且充分混匀。加入3倍体积的Trizol LS到样本中,在震荡器上充分混匀,室温放置5~15 min进行变性处理。按等体积加入氯仿,振荡器上充分混匀,室温放置15 min,在4 ℃条件下,14000 r/min离心15 min抽提RNA。
1.3.4芯片检测抽取每例AD组患者的2 μg总RNA,充分混匀后抽提RNA。使用Poly(A)聚合酶在总RNA 3’端加上Poly(A)尾巴,再将一个寡聚核苷酸标记与这个Poly(A)尾巴连接(ligation)用于后续的荧光标记。杂交反应利用微循环泵的循环作用在μParaflo微流体芯片上过夜进行。在微流体芯片上,每条检测探针都是由一个化学修饰核苷酸编码段或是与其他RNA和一个由聚乙二醇组成的间隔段组成。检测探针均使用PGR(PhotoGenerated Rea-gent)化学法进行原位合成。杂交解链温度是通过化学修饰检测探针进行平衡。RNA与探针杂交后,与标记特异结合的Cy3和Cy5染料在微流体芯片上循环流动进行染色。利用激光扫描仪采集杂交图像并使用Array-Pro图像分析软件进行图像数字化转换。数据分析首先是减除背景值,然后使用LOWESS过滤(Locally-Weighted Regression)进行信号归一化。计算两种检测信号的比值(log2转换)和t-test的P值。值<0.05的数据为差异性表达数据。
1.3.5逆转录用Taqman microRNA逆转录试剂盒和microRNA特异性茎环结构逆转录引物进行microRNA逆转录反应,操作按照ABI的流程,进行部分调整,总反应体系15 μl。
1.3.6QT-PCR检测根据芯片结果分别设计引物序列,以U6 snRNA作为内参,应用QT-PCR把两组患者的表达异常的microRNA进行定量分析,然后进行相互比较。用2-ΔΔCt计算microRNA的表达量,每例样本重复检测3次。
2 结 果
2.1一般资料结果AD组30例,男11例,女19例,平均年龄(63.3±11.5)岁;对照组30例,男13例,女17例,平均年龄(62.1±10.6)岁。两组在性别、年龄及常见危险因素方面差异无统计学意义,具有可比性(见表1)。
2.2基因芯片检测结果AD组检测出信号表达大于500的差异microRNAs43条(P<0.05),对差异表达microRNAs的信号比值进行聚类,可分为上调组和下调组,其中与对照组相比上调33条,下调10条(见图1)。
2.3MicroRNA-107QT-PCR结果选取芯片检测结果中差异表达有统计学意义的microRNA-107,进行QT-PCR验证。AD组患者血清中microRNA-107较对照组下调(P<0.01),与芯片结果一致(见表2、图2、图3)。
2.4MicroRNA-222QT-PCR结果选取芯片检测结果中差异表达有统计学意义的microRNA-222,进行实时荧光定量PCR 验证。AD组患者血清中microRNA-222明显下调(P<0.01),与芯片结果一致(见表3、图4、图5)。

表1 两组一般资料对比
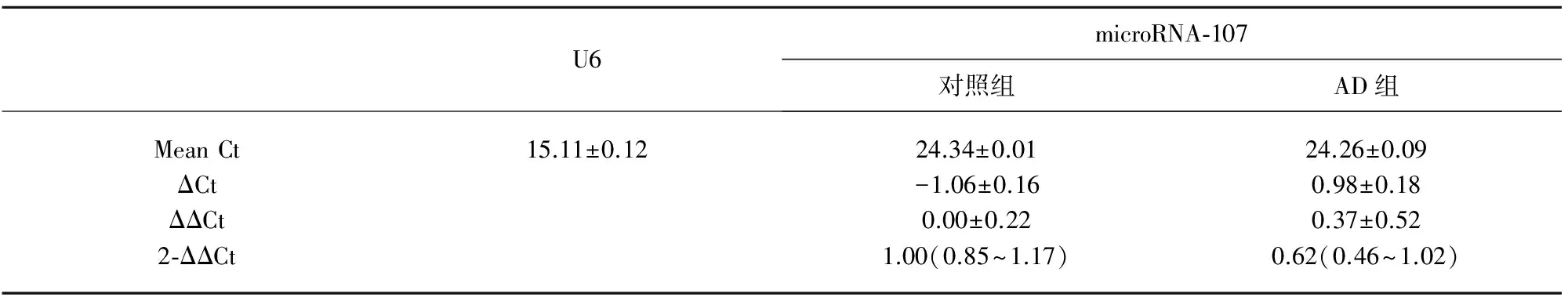
表2 两组microRNA-107定量结果
ΔCt表示同一个样品目标基因和内参基因平均Ct值的差值;ΔΔCt表示特定目标基因的待检样品和校准样品ΔCt的差值

表3 两组microRNA-222定量结果

图1 AD组与对照组差异microRNA聚类图
图2 microRNA-107扩增曲线及溶解曲线图

图3 以M为校准U6为内参microRNA-107基因表达水平

图4 microRNA-222扩增曲线及溶解曲线
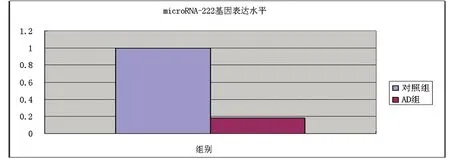
图5 以M为校准U6为内参microRNA-222基因表达水平
3 讨 论
近年来的研究表明,microRNAs与肿瘤、心血管疾病、糖尿病、人类遗传性疾病、神经系统疾病的发生发展密切相关。存在于脑部的microRNAs大约有20%~40%对脑的生长发育发挥着重要调控作用,甚至直接参与了哺乳动物神经细胞分化以及突触的形成。还有些microRNAs在神经退行性疾病中发挥着重要的作用[14]。Hebert等在AD的转基因小鼠模型里检测到对AD的影响microRNAs[15]。已有多项研究表明,microRNAs在细胞实验中能调节淀粉样前体蛋白(amyloid precursor protein,APP)的表达[16,17]及影响Aβ沉积[18]。某些microRNAs促进了Aβ产生,Aβ又可改变microRNAs表达,成为microRNAs网络失衡的始发因子[19],microRNAs与Aβ之间存在一种极其复杂的联系,在体内相互促进相互影响,从而影响着AD的发生发展。MicroRNA-125b被证明可引起tau蛋白高度磷酸化,其机制在于靶向抑制磷酸酶DUSP6(dual-specificity phosphatases 6)和PPPlCA而促进tau蛋白磷酸化,研究同时发现miR-125b可损害小鼠联合型学习[20]。可见,microRNAs从多种机制上参与了AD的发生。
目前有关microRNAs在AD患者中的研究少见报道,本研究通过生物芯片技术检测了AD患者与正常健康体检者血清中microRNAs的表达,结果表明有多种microRNAs表达差异有显著性。AD组检测出信号表达大于500的差异microRNAs43条,很多具有差异的microRNA在以往均少见相关研究报道,特别是差异较大的microRNA-4267、microRNA-30c-1-3p、microRNA-3197、microRNA-6820-5p、microRNA-6865-5p、microRNA-1910-3p等,这些microRNAs在AD发生发展中所起的作用还需要进一步的研究。
根据芯片结果,选取表达差异的microRNA-107采用QT-PCR进行验证,证实与芯片结果一致。本研究QT-PCR结果表明AD组患者血清中microRNA-107较对照组下调(P<0.01)。 microRNA-107隶属于microRNA-107/103家族,与AD的发生相关[21],Nelson发现AD和轻度认知功能障碍的患者中microRNA-107表达水平下降[22],与本研究结果一致。microRNA-107可能通过以下相关机制影响AD的发生发展。首先,Wang等[23]对不同阶段AD患者脑组织中的RNA进行分析,发现AD进程中miRNAl07的表达出现不同程度减少,而淀粉蛋白前β位分解酶1(BACEl)表达增加,从而出现相应的病理改变,该研究认为miRNA107可通过与靶基因BACEl的3’端UTR区结合调控BACEl的表达,继而增加APP水解后Aβ生成与沉积导致AD的发生,并可能通过调节BACE1参与加速该病的进展;其次,microRNA-107也可以诱导神经元细胞周期阻滞[24],并重新进入细胞周期,重新进入细胞周期是AD发病的早期事件甚至发生在Aβ和神经元纤维缠结(NFTs)形成之前[25],导致AD的发生。当然,microRNA-107影响AD的机制还需要进一步的研究。
QT-PCR结果表明AD组患者血清中microRNA-222明显下调(P<0.01),与芯片结果一致。众所周知,P27kipl是一种细胞周期依赖性激酶抑制因子,参与AD的发生、发展。Wang等观察AD模型小鼠并与年龄匹配的对照组相比发现,AD模型小鼠microRNA-222表达下调,与p27kip1蛋白水平升高相关,从而指出microRNA-222正是通过影响AD的细胞周期失调及p27kip1的表达而参与发病[26]。Zhang等[27]通过原位杂交与免疫组化检测,发现microRNA-222高表达的样本中有81%出现了PUMA(p53 up-regulated modulator of apoptosis)的表达水平降低(P<0.001),与此同时在microRNA-222下调的样本中有79%检测出了PUMA的高水平表达。PUMA是2001年由Yu等人发现的一个新bcl-2家族成员,其中PUMA-α和PUMA-β因含有BH3结构域而受到了广泛的研究,该结构域是PUMA蛋白与bcl-2家族其它成员相互作用,发挥促进神经细胞凋亡作用的关键因素,调控bcl-2基因的表达可参与AD的发病。已有的研究也表明,PUMA诱导细胞凋亡包括了p53依赖性凋亡通路和p53非依赖性凋亡通路,两条通路最终都通过诱导线粒体膜电位改变,引起细胞色素C等凋亡信号因子的释放,进而激活caspase-3和caspase-9最终导致细胞凋亡,caspase-3和caspase-9水平增加增强表达,从而导致AD的发生。Ok等的研究证实,caspase介导的APP切割产物与Aβ共定位于AD老年斑中,caspase在调节神经毒性Aβ的生成及在AD神经元最终的凋亡中有着极其重要的作用。microRNA-222参与AD的发病也与其在海马区域中的含量相比于其他区域特异性要高有关。结合本研究结果,血清中microRNA-107、microRNA-222可能间接反映脑组织内的microRNA-107、microRNA-222表达变化,但AD患者血清中表达水平下降的原因和机制尚需进一步研究。
综上所述,本研究结果显示microRNA-107、microRNA-222在AD患者中的表达量显著降低,可能作为AD的一种潜在的生物标志物,并且可能成为治疗靶点,但microRNA-107、microRNA-222表达水平与AD的诊断是否具有特异性,尚需进一步扩大样本量进行验证。
[1]Galimberti D,Scarpini E.Progress in Alzheimer’s disease[J].J Neurol,2012,259(2):201-211.
[2]Delay C,Mandemakers W,Hebert SS.MicroRNAs in Alzheimer’s disease[J].Neurobiol Dis,2012,46(2):285-290.
[3]Taki T.An approach to glycobiology from glycolipidomics:ganglioside molecular scanning in the brains of patients with Alzheimer’s disease by TLC-blot/matrix assisted laser desorption/ionization-time of flight MS[J].Biol Phmxn Bu,2012,35(10):1642-1647.
[4]Yang MH,Yang YH,Lu CY,et al.Activity-dependent neuroprotector homeobox protein:a candidate protein identified in serum as diagnostic biomarker for Alzheimer’s Disease[J].J Proteomics,2012,75(12):3617-3629.
[5]Czech C,Bemdt P,Busch K,et al.Metabolite profiling of Alzheimer’s disease cerebrospinal fluid[J].PLoS One,2012,7(2):e31501.
[6]Hu ZP,Browne ER,Liu T,et al.Metabonomic profiling of TASTPM transgenie Alzheimer’s disease mouse model[J].J Proteome Res,2012,11(12):5903-5913.
[7]Han X,Rozen S,Bovle SH,et al.Metabolomics in early Alzheimer’s disease:identification of altered plasma sphingolipidome using shotgun lipidomits[J].PLoS One,2011,6(7):e21643.
[8]Krol J,Loedige I,Filipowicz W.The widespread regulation of microRNA biogenesis,function and decay[J].Nat Rev Genet,2010,11:597-610.
[9]Valencia-Sanchez MA, Liu J, Hannon GJ,et al.Control of teanslation and microRNA degradation by microRNA and siRNAs[J].Genes Dev,2006,20:515-524.
[10]Cohen JE,Lee PR,Chen S,et al.MicroRNA regulation of homeostatic synaptic plasticity[J].Proc Natl Acad Sci USA,2011,108(28):11650-11655.
[11]Can J,Wang WY,Mao YW,et al.A novel pathway regulates memory and plasticity via SIRT1 and miR-134[J].Nature,2010,466(7310):1105-1109.
[12]Mo M,Chen L,Fu Y,et al.Cell-free Circulating miRNA biomarkers in cancer[J].J Cancer,2012,3(4):432-434.
[13]Resnik KE,Alder H,Hagan JP,et al.The detection of differentially expressed microRNAs from the serum of ovarian cancer patients using a novel real-time PCR platform[J].Gynecol Oncol,2009,112(1):55-59.
[14]Junn E,Mouradian MM.MicroRNAs in neurodegenemtive diseases and their therapeutic potential[J].Pharmacol Ther,2012,133(2):142-150.
[15]Hebert SS,Horre K,Nicolai L,et al.MicroRNA regulation of Alzheimer’s Amyloid precursor protein expression[J].Neurobiol Dis,2009,33(3):422-428.
[16]Long JM,Ray B,Lahiri DK.MicroRNA-153 physiologically inhibits expression of amyloid-beta precursor protein in cultured human fetal brain cells and is dysregulated in a subset of Alzheimer disease patients[J].J Biol Chem,2012,287(37):31298-31310.
[17]Long JM,Lahiri DK.MicroRNA-101 downregulates Alzheimer’s amyloid-β precursor protein levels in human cell cultures and is differentially expressed[J].Biochem Biophys Res Commun,2011,404(4):889-895.
[18]Fang M,Wang J,Zhang X,et al.The miR-124 regulates the expression of BACE1/B-secretase correlated with cell death in Alzheimer’s disease[J].Toxicol Lett,2012,209(1):94-105.
[19]Sohonrook N,Ke YD,Humphreys D,et al.Neuronal mircoRNA deregulation in response to Alzheimer’s disease amyloid-beta[J].PLoS One,2010,5(6):e11070.
[20]Banzhaf-Strathmann J,Benito E,May S,et al.MicroRNA-125b induces tau hyperphosphorylation and cognitive deficits in Alzheimer’s disease[J].EMBO J,2014,33(15):1667-1680.
[21]Miiller M,Kuiperij HB,Claassen JA,et al.MicroRNAs in Alzheimer's disease:differential expression in hippocampus and cell-free cerebrospinal fluid[J].Neurobiol Aging,2014,35(1):152-158.
[22]Nelson PT,Wang WX.MiR-107 is reduced in A1zheiIner’s diseasebrain neocortex:validation study[J].J Alzheimers Dis,20l0,2l(1):75-79.
[23]Wang WX,Rajeev BW,Stromberg AJ,et al.The expression of microRNA miR-107 decreases early in Alzheimer’s disease and may accelerate disease progression through regulation of betasite amyloid precursor protein-cleaving enzyme1[J].J Neurosci,2008,28(5):1213-1223.
[24]Feng L,Xie Y,Zhang H,et al.MiR-107 targets cyclin-dependent kinase 6 expression,induces cell cycle G1 arrest and inhibits invasion in gastric cancer cells[J].Med Oncol,2012,29(2):856-863.
[25]Bonda DJ,Lee HP,Kudo W,et al.Pathological implications of cell cycle re-entry in Alzheimer’s disease[J].Expert Rev Mol Med,2010,12:e19.
[26]Wang X,Xu Y,Zhu H,et al.Downregulated microRNA-222 is correlated with increased p27Kip1 expression in a double transgenic mouse model of Alzheimer’s disease[J].Mol Med Rep,2015,12 (5):7687-7692.
[27]Zhang CZ,Zhang JX,Zhang AL,et al.MiR-221 and MiR-222 target PUMA to induce cell survival in glioblastoma[J].Mol Cancer,2010,9(1):229.
Study on microRNAs expression in serum of patients with Alzheimer’s disease
ZENGQinghong,LIUXia,ZHOUFang,etal.
(DepartmentofNeurology,TheSecondPeople’sHospitalofLianyungang,Lianyungang22206,China)
ObjectiveTo explore the relationship between microRNAs and Alzheimer’s disease in serum of patients with Alzheimer’s disease,in order to find an early biomarker for diagnosis of AD.MethodsThe study subjects were divided into AD group and control group.MicroRNAs was extracted from serum.The expression of microRNAs was detected by microRNA gene chip,and the method was verified by fluorescence quantitative PCR.Results43 AD related genes were selected to screen the differentially expressed microRNAs gene (P<0.05),and the expression level of microRNA-107 and microRNA-222 was significantly lower in AD patients than in control group (P<0.01),and the results were consistent with gene chipQT-PCR.ConclusionMicroRNA-107 and microRNA-222 may influence the occurrence and development of AD in many ways,which is expected to be an early biomarker for the diagnosis of AD.
Alzheimer’s disease;microRNAs;microRNA-107;microRNA-222
1003-2754(2016)02-0123-05
2015-12-15;
2016-01-29
连云港市科技局课题基金(SH1224)
(1.连云港市第二人民医院神经内科,江苏 连云港 222006;2.连云港市第二人民医院检验科,江苏 连云港 222006)
庄爱霞,E-mail:aixiazhuang1965@sina.com
R749.1
A
猜你喜欢
中老年保健(2022年2期)2022-08-24 03:21:24
中老年保健(2022年1期)2022-08-17 06:14:36
中老年保健(2021年3期)2021-08-22 06:50:04
天津医科大学学报(2021年2期)2021-03-29 05:31:08
现代临床医学(2021年1期)2021-01-26 00:56:02
奥秘(2018年9期)2018-09-25 03:49:56
发明与创新(2016年23期)2016-10-13 02:16:14
湖北工业大学学报(2016年5期)2016-02-27 13:14:51
医学研究杂志(2015年4期)2015-06-10 06:42:43
河南科技(2014年10期)2014-02-27 14:09:18
