
4种蕨类植物的孢子繁殖研究
2016-11-09 11:25朱晓凤金冬梅赵国华刘保东凌铁军严岳鸿
植物研究 2016年2期
朱晓凤 沈 慧 金冬梅 赵国华, 刘保东 凌铁军 严岳鸿
(1.上海辰山植物园(中国科学院上海辰山植物科学研究中心),上海 201602; 2.安徽农业大学省部共建茶树生物学与资源利用国家重点实验室,合肥 230036; 3.哈尔滨师范大学黑龙江省普通高等学校植物生物学重点实验室,哈尔滨 150025)
* 通信作者:E-mail:shenhui@sibs.ac.cn
4种蕨类植物的孢子繁殖研究
朱晓凤1,2沈 慧1*金冬梅1赵国华1,3刘保东3凌铁军2严岳鸿1
(1.上海辰山植物园(中国科学院上海辰山植物科学研究中心),上海 201602;2.安徽农业大学省部共建茶树生物学与资源利用国家重点实验室,合肥 230036;3.哈尔滨师范大学黑龙江省普通高等学校植物生物学重点实验室,哈尔滨 150025)
蕨类植物的资源开发越来越受到重视,但目前的孢子繁殖技术还不够成熟,已成为其市场扩大的制约因素。本研究对4种有代表性的资源型蕨类植物乌毛蕨(Blechnumorientale)、荚果蕨(Matteucciastruthiopteris)、假鞭叶铁线蕨(Adiantummalesianum)和水蕨(Ceratopteristhalictroides)进行了孢子繁殖研究,详细观察并记录了繁殖过程中配子体发育各阶段出现的特征和时间。其中以水蕨的发育周期最短,从播种到形成幼孢子体约33天;假鞭叶铁线蕨发育周期最长,约73天;乌毛蕨和荚果蕨的发育周期介于二者之间。结合过去已发表的相关观察资料,对4种蕨类的孢子萌发、原叶体、性器官以及幼孢子体形成等四个生长关键点所出现时间的培养条件进行了分析,结果表明:(1)光照时间越长,孢子萌发天数越短;(2)光照强度越大,原叶体发育天数越长;(3)配子体发育速率在20~25℃范围内与温度无显著相关性。本研究可为进一步探究蕨类植物适宜的繁殖技术体系和开发蕨类资源提供指导。
蕨类植物;孢子繁殖;资源开发;环境因子
蕨类植物是一类通过孢子进行繁殖,不形成花或种子的维管植物[1],在食用、药用、观赏以及工业等方面均得到了广泛的应用[2~7]。蕨类植物的资源开发不能仅从野外获取,而其分株繁殖等无性繁殖方式的繁殖效率低,生产成本高,均不利于资源开发。采用孢子繁殖是一个有效途径,其繁殖效率高,资源破坏程度小,已成为蕨类植物资源可持续开发利用的基础。据统计,目前国内已经完成34科、80属、157种蕨类植物的配子体发育或孢子繁殖研究[8]。然而不同的蕨类植物的孢子繁殖具有不同的繁殖特点,生产实践工作者难以掌握一般的孢子繁殖规律,对资源蕨类植物高效率的开发应用形成了制约。
针对乌毛蕨(Blechnumorientale)、荚果蕨(Matteucciastruthiopteris)、假鞭叶铁线蕨(Adiantummalesianum)和水蕨(Ceratopteristhalictroides)4种蕨类植物的孢子繁殖过去已开展了多项不同的研究,其孢子萌发时间在3~20 d、幼孢子体形成时间在40~98 d范围内,因培养条件的不同生长周期略有差异[9~38]。他们对4种蕨类配子体发育的阶段特征也做了大量显微观察及描述工作,但仍不够全面,需要进一步补充。过去的研究仅针对某一种蕨类植物探究光照、水分、培养基质等环境因子的影响[9~14],未将不同生境的不同蕨类植物进行比较繁殖来研究蕨类孢子繁殖中普适性的关键生态因子。
本研究选择四种具有代表性的资源型蕨类植物,包括幼叶可食、成熟植株可供观赏、根状茎可入药等具综合资源利用价值的大型丛生蕨类乌毛蕨、中型丛生蕨类荚果蕨、水生蕨类水蕨和具有较高观赏价值的小型丛生蕨类假鞭叶铁线蕨等进行孢子繁殖特征的观察和记录,结合已发表的研究资料,对这4种蕨类植物配子体发育各阶段出现的时间和环境因子进行分析。旨在通过探讨这4种蕨类孢子繁殖过程中各阶段出现关键特征的时间与培养条件的关系,摸索同类型蕨类植物适宜的孢子繁殖条件,提供基础资料并建立适宜的繁殖技术体系,将这些蕨类植物扩大种群数量并进一步引向市场,开发其综合利用价值。
1 材料与方法
1.1 孢子材料
乌毛蕨孢子于2014年10月采自广西桂林植物研究所;荚果蕨孢子于2014年11月采自上海辰山植物园蕨类植物区;水蕨孢子于2014年11月采自上海辰山植物科研中心人工气候室;假鞭叶铁线蕨孢子于2014年7月采自云南元江哀牢山。将具有成熟孢子的叶片剪下,放入洁净的硫酸纸袋中密封,置于干燥通风处,4~6 d后孢子自然散落,收集于离心管中在4℃冰箱内保存备用。
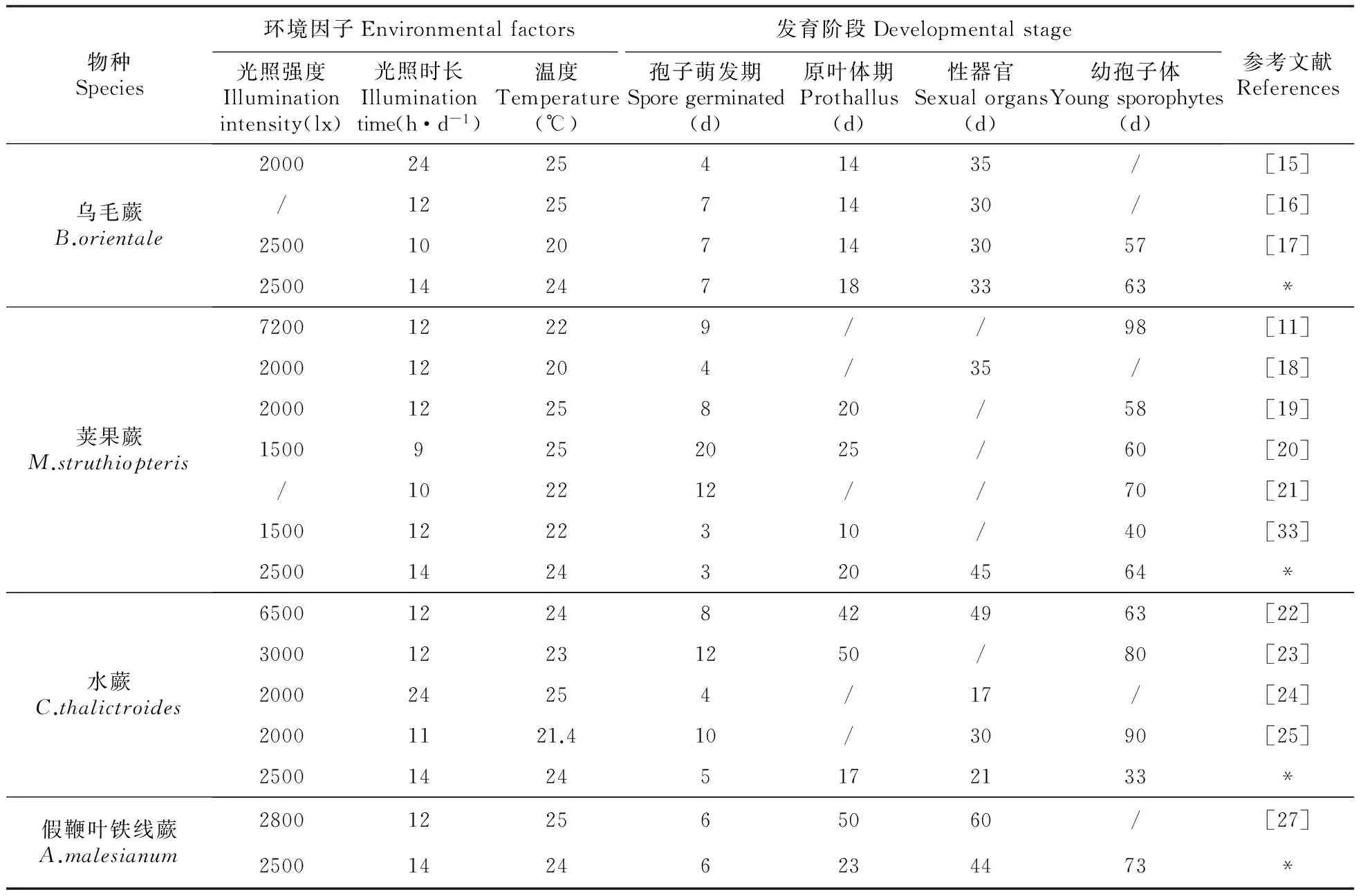
表1 环境因子和配子体发育各阶段出现时间对照表
注:*为4种蕨类孢子繁殖的相关数据
Note:*.Related data of spore propagation about 4 ferns.
1.2 土壤材料
采用上海辰山林下腐殖土,将其在日光下暴晒1周后压碎,过5号筛,装在聚丙烯罐中高温灭菌(121℃,40 min)后待用。
1.3 培养方法
把土壤放入事先灭过菌、底部铺有海绵的20 cm×25 cm的不锈钢盘中,厚约3 cm,润湿后将孢子均匀地播在土壤表面,盖上玻璃封住盘口,置于人工气候室内培养,温度为24±1.2℃,空气相对湿度约70%,日光灯照射,光照14 h·d-1,黑暗10 h·d-1,光照强度约2 500 lx。
幼孢子体苗长至1.5 cm高时进行分苗移栽,每簇2~3株,株行距2 cm×2 cm。移栽后先缓苗2~3 d,环境条件不变。苗高4 cm左右时进行移栽,移至口径10 cm的塑料花盆里,栽培基质为腐叶土,室内温度15~25℃,自然光照10~12 h·d-1,每天喷水一次,遮阴保湿,湿度约70%。幼苗高10 cm左右进行室外炼苗,30 d后可定植于自然环境下。室外温度0~32℃,自然光照,湿度大于30%。
1.4 观察方法
各发育阶段的形态特征在体式解剖镜(型号Nikon SMZ-1500)下观察拍照或做成临时装片在光学显微镜(型号Axio Scope A1)下观察拍照。随机选取20个个体,用ImageJ测量软件测量相关形态数据,计平均值。
时间统计方法:每3 d观察1次,记录孢子萌发时间、丝状体形成时间、片状体形成时间、原叶体形成时间、胚形成时间、幼孢子体形成时间。将不锈钢盘分成4 cm×5 cm大小的5个分区,以每个阶段标志性现象出现数量超过50%为限。共统计了60株,重复3次,计平均值。
1.5 作图方法
4种蕨类植物配子体各发育阶段特征图片用Photoshop进行组合;同时利用R3.2.0软件对环境因子和配子体发育各阶段出现时间(表1)作散点图,并标出线性回归的趋势[39~40]。
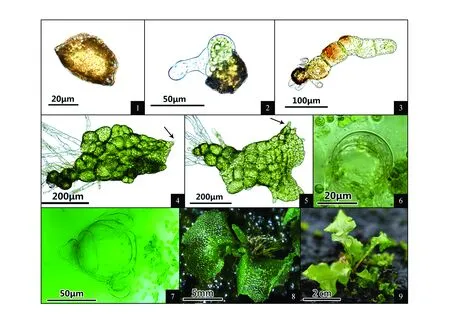
图版Ⅰ 乌毛蕨配子体各发育阶段特征 1.孢子形态;2.孢子萌发;3.丝状体;4.片状体;5.原叶体;6.精子器;7.颈卵器;8.幼孢子体;9.幼苗PlateⅠ Spore propagation characteristics of B.orientale at each stage 1.Spore morphology; 2.Spore germinated; 3.Filament; 4.Plate; 5.Prothallus; 6.Antheridium; 7.Archegonium; 8.Young sporophytes; 9.Seedling
2 结果
2.1 4种蕨类孢子繁殖各阶段的特征
2.1.1 乌毛蕨配子体及幼孢子体发育特征描述
(1)孢子及孢子萌发:孢子棕褐色,两面体型,单裂缝,孢子大小17 μm×29 μm(图版Ⅰ:1)。播种后7 d左右,孢子吸水膨胀,萌发形成初生假根和原始细胞(图版Ⅰ:2)。萌发类型为书带蕨型[29]。(2)丝状体:播种9 d左右出现第二个细胞,形成具有3~8个圆柱状细胞的丝状体(图版Ⅰ:3)。(3)片状体:无明显顶端细胞,前端边缘出现个别毛状体(图版Ⅰ:4箭头所示)。(4)原叶体:播种18 d左右形成心脏形,边缘有毛状体(图版Ⅰ:5箭头所示)。原叶体发育类型为三叉蕨型。(5)性器官:播种33 d左右性器官出现,精子器近圆球形,直径约34 μm(图版Ⅰ:6);颈卵器顶面观铜钱状,3~5个细胞高,直径约58 μm(图版Ⅰ:7)。(7)幼孢子体:播种后63 d左右形成幼孢子体(图版Ⅰ:8),直至长出2~3片叶子后原叶体枯死(图版Ⅰ:9)。
2.1.2 荚果蕨配子体及幼孢子体发育特征描述
(1)孢子及孢子萌发:孢子黑褐色,两面体型,单裂缝,大小44 μm×63 μm(图版Ⅱ:1)。播种后3 d左右,孢子吸水膨胀,萌发形成初生假根和原始细胞(图版Ⅱ:2)。萌发类型为紫萁型[29]。(2)丝状体:播种5 d左右出现第二个细胞。丝状体单列或双列,可形成3~14个细胞长(图版Ⅱ:3)。(3)片状体:丝状体顶端细胞不断分裂成片状体(图版Ⅱ:4)。(4)原叶体:播种20 d左右形成规整的心脏形原叶体(图版Ⅱ:5)。原叶体发育类型为铁线蕨型。(5)性器官:播种45 d左右性器官出现,精子器近圆球形,直径约39 μm(图版Ⅱ:6);颈卵器直径约62 μm,颈部由4~5层颈壁细胞组成(图版Ⅱ:7)。(6)幼孢子体:播种64 d左右形成幼孢子体(图版Ⅱ:8),直至幼苗长到7 cm左右原叶体才枯死(图版Ⅱ:9)。

图版Ⅱ 荚果蕨配子体各发育阶段特征 1.孢子形态;2.孢子萌发;3.丝状体;4.片状体;5.原叶体;6.精子器;7.颈卵器;8.幼孢子体;9.幼苗Plate Ⅱ Spore propagation characteristics of M.struthiopteris at each stage 1.Spore morphology; 2.Spore germinated; 3.Filament; 4.Plate; 5.Prothallus; 6.Antheridium; 7.Archegonium; 8.Young sporophytes; 9.Seedling
2.1.3 水蕨配子体及幼孢子体发育特征描述
(1)孢子及孢子萌发:孢子黄褐色,四面体型,三裂缝,外壁有肋条状纹饰,大小108 μm×120 μm(图版Ⅲ:1)。播种后5 d左右,孢子吸水膨胀,从三裂缝中伸出初生假根,萌发成原始细胞(图版Ⅲ:2)。萌发类型为书带蕨型[29]。(2)丝状体:播种8d左右出现第二个细胞(图版Ⅲ:3)。丝状体有2~7个细胞,单列或双列,呈圆锥状。(3)片状体:匙形,基部2~4列细胞宽(图版Ⅲ:4),无顶端细胞。(4)原叶体:播种17 d左右片状体一侧出现分生细胞,形成不对称的心脏形原叶体(图版Ⅲ:5)。原叶体发育类型为水蕨型。(5)性器官:播种21 d左右性器官出现,原叶体边缘的精子器铜钱状,直径约57.4 μm(图版Ⅲ:6);颈卵器直径约62.5 μm,颈部由4~5层颈壁细胞组成(图版Ⅲ:7)。(6)幼孢子体:播种33 d左右形成幼孢子体(图版Ⅲ:8),直至长出2~3片叶子后原叶体枯死(图版Ⅲ:9)。
2.1.4假鞭叶铁线蕨配子体及幼孢子体发育特征描述
(1)孢子及孢子萌发:孢子红棕色,四面体型,三裂缝,外壁有瘤状纹饰,大小30 μm×34 μm(图版Ⅳ:1)。播种后6 d左右,从三裂缝破壁,孢子第一次分裂形成一大一小两个细胞,小的伸长为假根,大的为原始细胞(图版Ⅳ:2)。萌发类型为紫萁型[29]。(2)丝状体:播种11 d左右出现第二个细胞(图版Ⅳ:3)。丝状体有2~5个细胞,单列或双列。(3)片状体:具顶端细胞,经一次纵裂后交替横裂形成片状体(图版Ⅳ:4)。(4)原叶体:播种23 d左右片状体前端内凹,形成心脏形原叶体(图版Ⅳ:5)。原叶体发育类型为铁线蕨型。(5)性器官:播种44 d左右性器官出现,精子器圆球形,直径约50 μm(图版Ⅳ:6);颈卵器顶面观铜钱状,4~5个细胞高,直径约58 μm(图版Ⅳ:7)。(6)幼孢子体:播种73 d左右形成幼孢子体(图版Ⅳ:8),直至长出2~3片叶子后原叶体枯死(图版Ⅳ:9)。

图版Ⅲ 水蕨配子体各发育阶段特征 1.孢子形态;2孢子萌发;3.丝状体;4.片状体;5.原叶体;6.精子器;7.颈卵器;8.幼孢子体;9.幼苗Plate Ⅲ Spore propagation characteristics of C.thalictroides at each stage 1.Spore morphology; 2.Spore germinated; 3.Filament; 4.Plate; 5.Prothallus; 6.Antheridium; 7.Archegonium; 8.Young sporophytes; 9.Seedling
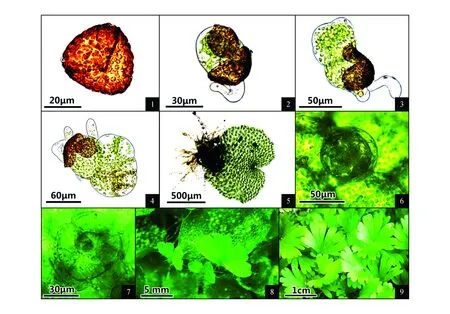
图版Ⅳ 假鞭叶铁线蕨配子体各发育阶段特征 1.孢子形态;2孢子萌发;3.丝状体;4.片状体;5.原叶体;6.精子器;7.颈卵器;8.幼孢子体;9.幼苗Plate Ⅳ Spore propagation characteristics of A.malesianumat each stage 1.Spore morphology; 2.Spore germinated; 3.Filament; 4.Plate; 5.Prothallus; 6.Antheridium; 7.Archegonium;8.Young sporophytes; 9.Seedling
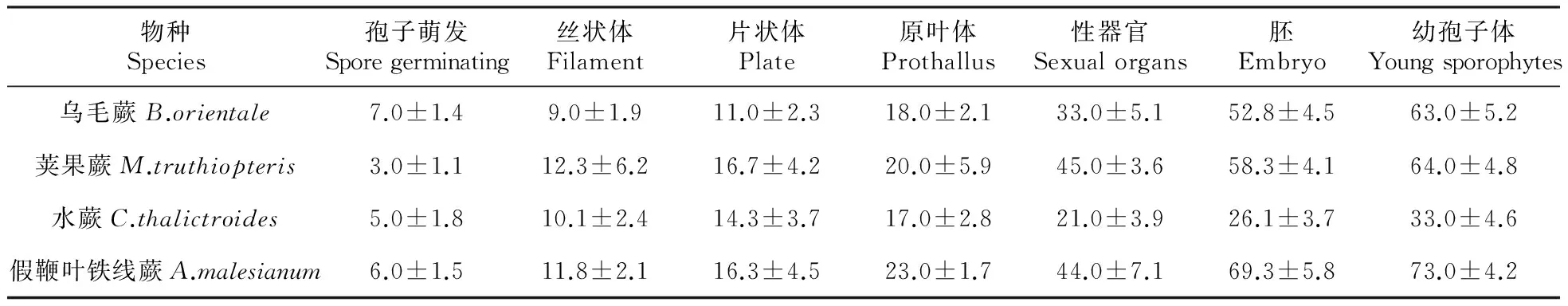
物种Species孢子萌发Sporegerminating丝状体Filament片状体Plate原叶体Prothallus性器官Sexualorgans胚Embryo幼孢子体Youngsporophytes乌毛蕨B.orientale7.0±1.49.0±1.911.0±2.318.0±2.133.0±5.152.8±4.563.0±5.2荚果蕨M.truthiopteris3.0±1.112.3±6.216.7±4.220.0±5.945.0±3.658.3±4.164.0±4.8水蕨C.thalictroides5.0±1.810.1±2.414.3±3.717.0±2.821.0±3.926.1±3.733.0±4.6假鞭叶铁线蕨A.malesianum6.0±1.511.8±2.116.3±4.523.0±1.744.0±7.169.3±5.873.0±4.2
2.2 4种蕨类孢子繁殖各阶段特征时间统计
4种观赏蕨类孢子繁殖各阶段的时间统计结果如表2所示,可以看出,本文研究的4种蕨类孢子萌发时间在接种后3~7 d;丝状体期在9~12 d;片状体期在11~17 d;原叶体期在17~23 d;性器官形成时间在21~45 d;胚形成时间在26~69 d;幼孢子体形成时间在33~73 d。其中以水蕨的发育周期最短,从播种到形成幼孢子体约33 d;假鞭叶铁线蕨发育周期最长,约73 d;乌毛蕨和荚果蕨的发育周期介于二者之间。
2.34种蕨类植物配子体各时期发育天数与环境因子的关系
2.3.14种蕨类植物配子体各时期发育天数与光照时长的关系
在4种蕨类的孢子萌发期(图1:A)、水蕨的幼孢子体期(图1:D),线性回归呈现负相关的趋势,表明以上阶段的发育受光照时长的影响较为显著,即光照时间越长,4种蕨类孢子萌发越快、水蕨的幼孢子体形成越快。
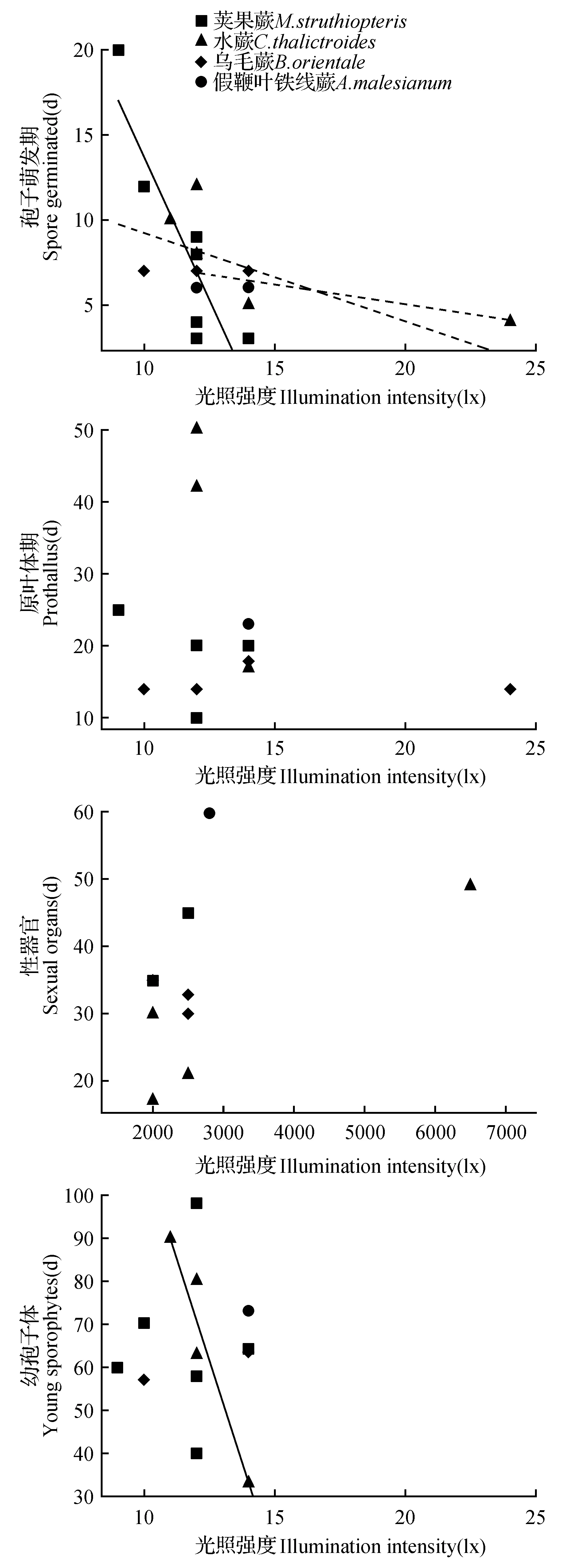
图1 配子体各时期发育天数与光照时长关系的散点图实线表示回归线P<0.05,虚线表示回归线P<0.1,下同。Fig.1 Scatter diagram between gametophyte development time at each stage and illumination time
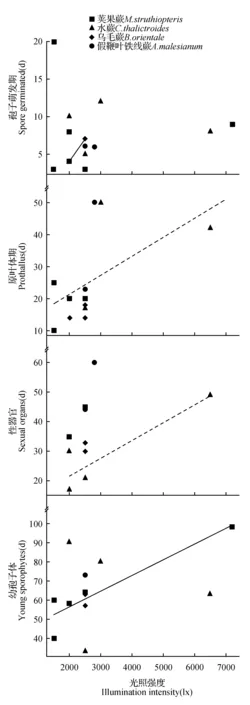
图2 配子体各时期发育天数与光照强度关系的散点图Fig.2 Scatter diagram between gametophyte development time at each stage and illumination intensity
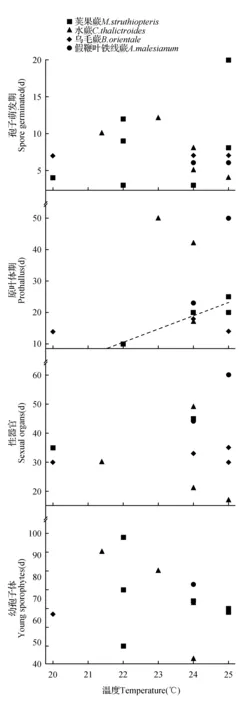
图3 配子体各时期发育天数与温度关系的散点图Fig.3 Scatter diagram between gametophyte development time at each stage and temperature
2.3.24种蕨类植物配子体各时期发育天数与光照强度的关系
在乌毛蕨的孢子萌发期(图2:A)、4种蕨类的原叶体期(图2:B)、水蕨的性器官期(图2:C)和荚果蕨的幼孢子体期(图2:D),线性回归均呈现正相关的趋势,表明以上阶段的发育受光照强度的影响较为显著,即光照强度越大,发育天数越长。
2.3.34种蕨类植物配子体各时期发育天数与温度的关系
在20~25℃范围内,4种蕨类植物配子体各时期发育天数与温度相关性不明显,仅在荚果蕨原叶体期呈现较弱的正相关趋势(图3:B),即温度越高,荚果蕨原叶体发育天数越长。
3 讨论
本研究结果中,4种蕨类植物形态特征方面无变异情况,均为正常发育。乌毛蕨、荚果蕨、水蕨和假鞭叶铁线蕨完成配子体发育史的时间分别约63、64、33和73 d,与文献报道[15~28]有一定的差异。不同种类的蕨类植物因其生境迥异在不同的培养条件下生长速度有差异,其中光照时长、光照强度、温度等3个环境因子的影响较大[33~37,41~48]。
3.1光照时长对蕨类植物配子体发育时间差异的影响
本文研究结果表明光照时间越长,4种蕨类孢子萌发越快、水蕨的幼孢子体形成越快。戴锡玲[15,24]采用全天光照培养水蕨和乌毛蕨,二者均可正常生长,据此我们推测进一步延长光照时间可能不会阻碍蕨类植物的生长发育。
3.2光照强度对蕨类植物配子体发育时间差异的影响
无论是大型丛生蕨类(乌毛蕨)、中型丛生蕨类(荚果蕨)、水生蕨类(水蕨)还是小型丛生蕨类(假鞭叶铁线蕨)在原叶体期发育天数上均与光照强度表现出呈现较强的正相关性,即光照强度越大,原叶体发育越慢。已有报道表明,蕨类植物配子体在黑暗条件或者高于4 800 lx光强的条件下无法正常发育[9,41~44]。原因是在配子体发育阶段,黑暗条件下其叶绿体无法形成;但较弱的光照强度就足以使配子体光形态建成,如果光照强度过大则会抑制细胞的分裂,使配子体的发育停滞[47~48]。蕨类植物配子体总体上在光照强度较低时发育较快,可能与蕨类植物长期适应林下阴暗环境有关。
3.3 温度对蕨类植物配子体发育时间差异的影响
本文中用于分析的温度在20~25℃内,梯度较小,因而不能准确地反映温度和配子体发育速率之间的相关性。有研究表明,蕨类植物配子体在低于15℃或者高于30℃的温度下无法正常发育[9,41~46]。蕨类植物常生于林下、溪边等较阴凉的地方,当植株处于温度胁迫时,细胞质膜相对透性增强,造成溶质外渗增加,使植株膜系统受到明显伤害而无法正常生长[47~48],因此20~25℃是较为适宜的生长温度。
综上所述,我们推断适宜的温度,较弱的光照强度以及较长的光照时间有利于蕨类植物的配子体发育,建立适宜的繁殖技术体系可从这几个方面着手,这在实际生产中对快速获得种苗具有指导意义。
1.陆树刚.蕨类植物学[M].北京:高等教育出版社,2007
2.石雷.观赏蕨类[M].北京:中国林业出版社,2002.
3.夏冰.民族植物学和药用植物[M].南京:东南大学出版社,2006.
4.王俊丽.中国民族地区药用植物资源利用与生物技术[M].北京:科学出版社,2012.
5.王振宇.食用植物资源学:食用植物开发利用原理[M].哈尔滨:东北林业大学出版社,2003.
6.高愿军.中国野生植物开发与加工利用[M].北京:中国轻工业出版社,1995.
7.曾汉元,丁炳扬.蕨类植物孢子萌发及原叶体发育的观察[J].植物科学学报,2004,22(4):368-371.
8.严岳鸿,石雷.蕨类植物迁地保护的方法与实践[M].北京:中国林业出版社,2014.
9.李晓.荚果蕨生态适应性调查与孢子繁殖技术研究[D].长春:吉林农业大学,2011.
10.魏琳.不同栽培条件对荚果蕨萌发与生长及其生理变化的影响[D].哈尔滨:东北林业大学,2012.
11.魏琳,赵雨森,高德武,等.不同水分处理对荚果蕨幼苗生长和生理特性的影响[J].东北林业大学学报,2012,40(7):64-67.
12.徐金忠,高德武,孙雪文,等.不同有机肥处理对荚果蕨幼苗生长及光合特性的影响[J].中国农学通报,2013,29(13):192-196.
13.徐洪亮,高德武,李日新,等.不同遮阴措施对荚果蕨幼苗生长和生理特性的影响[J].水土保持应用技术,2013(2):1-3.
14.陈蔚辉,刘俊祥.生态因子对水蕨生长的影响[J].湖南农业大学学报:自然科学版,2009,35(5):493-496.
15.戴锡玲,王全喜,李金国.液体培养基条件下乌毛蕨配子体发育的研究[J].武汉植物学研究,2007,25(2):205-208.
16.张开梅,石雷,李东.乌毛蕨配子体发育的研究[J].热带亚热带植物学报,2005,13(5):419- 422.
17.王玥,赵金博,王金娟等.乌毛蕨科3种植物配子体发育的研究[J].植物研究,2007,27(3):269-274.
18.包文美,敖志文,刘保东,等.东北蕨类植物配子体发育的研究—Ⅴ.球子蕨科[J].植物研究,1988,8(3):139-208.
19.董丽,苏雪痕.荚果蕨MatteucciastruthiopterisTodaro孢子繁殖的研究[J].园艺学报,1993,20(3):274-278.
20.陈龙清,季华,袁芳亭.荚果蕨孢子的无菌培养[J].植物生理学通讯,2000,36(5):436-437.
21.高德武,石长金,李日新,等.荚果蕨人工繁育技术研究[J].中国农学通报.2008,24(2):303-307.
22.Nayar B K,Kaur S.A reinvestigation of the morphology of the gametophyte and juvenile sporophyte ofCeratopteristhalictroides[J].Canadian Journal of Botany,1969,47(3):395-404.
23.黄碧光,陈庆山,林文雄.水蕨的农田生境调查和人工繁育探索[J].中国生态农业学报,2012,20(6):810-812.
24.戴锡玲,金沁,王全喜.水蕨配子体发育的研究[J].植物研究,2006,25(3):274-276.
25.陈蔚辉,肖卫彬.水蕨配子体和孢子体发育的研究[J].湖南农业大学学报:自然科学版,2008,34(3):306-310.
26.包文美,敖志文,刘保东.东北蕨类植物配子体发育的研究—Ⅳ.铁线蕨科[J].植物研究,1987,7(4):54-62.
27.罗顺元,王任翔.假鞭叶铁线蕨孢子的组织培养[J].植物生理学通讯,2007,43(1):131-132.
28.罗顺元.假鞭叶铁线蕨配子体发育的研究[J].热带亚热带植物学报.2009,17(2):164-168.
29.Nayar B K,Kaur S.Gametophytes of homosporous ferns [J].The Botanical Review,1971,37(3):295-396.
30.曾汉元,丁炳扬.蕨类植物孢子萌发及原叶体发育的观察[J].武汉植物学研究.2004,22(4):368-371.
31.曾汉元,丁炳扬.蕨类植物配子体发育的研究[J].植物研究.2003,23(2):154-158.
32.郝丽珍,邓俊玲.蕨菜孢子萌发及生育进程与诸因素关系研究[J].内蒙古农牧学院学报,1998,19(2):75-80.
33.韩玉林,贺善安,顾姻,等.环境条件对荚果蕨孢子繁殖的影响[J].黑龙江八一农垦大学学报,1992,1(3):27-32.
34.Nayar B K.comparative study of the spore morphology of Ceratopteris,Anemia and Mohria,and its bearing on the relationships of the Parkeriaceae[J].Indian Bot Soc J,1968,47(1):246-256.
35.Schedlbauer M D.Fern gametophyte development:controls of dimorphism inCeratopteristhalictroides[J].American Journal of Botany,1976,63(8):1080-1087.
36.Sayers A,Hamilton R G.The effect of neighbors on gametophyte development inCeratopterisrichardii[J].American Fern Journal,1995,85(2):47-53.
37.Banks J A.Sex-determining genes in the homosporous fernCeratopteris[J].Development,1994,120(7):1949-1958.
38.Kachroo R,Nayar B K.Studies in Pteridaceae observations on gametophytes of some species ofAdiantumL.[J].Phytomorphogy,1953,3(2):240-248.
39.(美)麦特洛夫(Matloff N)著.陈堰平等,译.R语言编程艺术[M].北京:机械工业出版社,2013.
40.Maindonald J,Braun J.Data analysis and graphics using R:an example-based approach[M].Cambridge University Press,2006.
41.Suo J,Chen S,Zhao Q,et al.Fern spore germination in response to environmental factors[J].Frontiers in Biology,2015:1-19.
42.Nondorf S L,Dooley M A,Palmieri M,et al.The effects of pH,temperature,light intensity,light quality,and moisture levels on spore germination in Cheilanthes feei of Southeast Missouri[J].American Fern Journal,2003,93(2):56-69.
43.Miller J H,Miller P M.The effect of different light conditions and sucrose on the growth and development of the gametophyte of the fern,Onocleasensibilis[J].American Journal of Botany,1961:154-159.
44.Ospina K R,Briones O,Pérez-García B.Spore Germination of Three Tree Fern Species in Response to Light,Water Potential,and Canopy Openness[J].American Fern Journal,2015,105(2):59-72.
45.Kodama Y,Tsuboi H,Kagawa T,et al.Low temperature-induced chloroplast relocation mediated by a blue light receptor,phototropin 2,in fern gametophytes[J].Journal of plant research,2008,121(4):441-448.
46.张正修,戴绍军.环境因子对蕨类植物孢子萌发的影响[J].生态学报,2010(7):1882-1893.
47.Whittier D P.The effect of light and other factors on spore germination inBotrychiumdissectum[J].Canadian Journal of Botany,1973,51(10):1791-1794.
48.Pérez-García B,Mendoza-Ruiz A,Sánchez-Coronado M E,et al.Effect of light and temperature on germination of spores of four tropical fern species[J].Acta Oecologica,2007,32(2):172-178.
SporePropagationofFourFerns
ZHU Xiao-Feng1,2SHEN Hui1*JIN Dong-Mei1ZHAO Guo-Hua1,3LIU Bao-Dong3LING Tie-Jun2YAN Yue-Hong1
(1.Shanghai Chenshan Plant Science Research Center,Chinese Academy of Sciences,Shanghai 201602;2.State Key Laboratory of Tea Plant Biology and Utilization,Anhui Agricultural University,Hefei 230036;3.Key Laboratory of Plant Biology,Harbin Normal University,College of Heilongjiang Province,Harbin 150025)
The development of fern resources is attracting more attention in recent years, however, current spore reproductive technology is immature, and has become a bottleneck. We cultured the spores of four typical ferns, includingBlechnumorientale,Matteucciastruthiopteris,CeratopteristhalictroidesandAdiantummalesianum, observed spore propagation characteristics and time of the gametophyte development at each stage. By compiling the reported data, we analyzed the culture conditions, such as light and temperature, in four key development stages, including spore germination, and the formation of prothalli, sexual organs, and juvenile sporophyte. Longer illumination time may promote spore germination. Higher light intensity may prolong the formation of prothallus. Growth rate and temperature had no correlation during gametophyte formation in 20-25℃. Our research is useful for establishing the system of fern propagation and may promote the exploration of fern resources.
pteridophyte;spore propagation;resource development;environmental factor
国家自然科学基金(31370234);中国科学院战略生物资源科技支撑体系运行专项(CZBZX-1);上海绿化局科技攻关项目(F122421)
朱晓凤(1990—),女,硕士研究生,主要从事蕨类植物孢子繁殖方面的研究。
2015-09-29
S682.35
A
10.7525/j.issn.1673-5102.2016.02.003
猜你喜欢
渔业研究(2022年4期)2022-08-27
花生学报(2022年1期)2022-08-16
草地学报(2019年2期)2019-05-31
农机化研究(2019年9期)2019-05-24
渔业研究(2019年1期)2019-03-04
西北植物学报(2018年6期)2018-07-19
飞碟探索(2016年9期)2016-09-21
中国民族医药杂志(2016年2期)2016-05-14
中国民族医药杂志(2016年2期)2016-05-14
中国民族医药杂志(2016年3期)2016-05-09
