
一株耐甲氧西林金黄色葡萄球菌噬菌体qdsa002的分离鉴定及其生理学性质研究
2016-11-08 07:16王静雪李梦哲毛相朝
食品工业科技 2016年16期
鞠 磊,王静雪,林 洪,李梦哲,毛相朝
(中国海洋大学食品科学与工程学院,食品安全实验室,山东青岛 266003)
一株耐甲氧西林金黄色葡萄球菌噬菌体qdsa002的分离鉴定及其生理学性质研究
鞠磊,王静雪*,林洪,李梦哲,毛相朝
(中国海洋大学食品科学与工程学院,食品安全实验室,山东青岛 266003)
为探究耐甲氧西林金黄色葡萄球菌的生物防治方法,从青岛市团岛污水处理厂采集到的污水中分离得到了一株噬菌体qdsa002。借助噬菌斑形态、酶切、电镜等技术对其进行了分类鉴定。研究了其最佳感染复数、一步生长曲线、pH稳定性和热稳定性等生理学特征。结果表明,qdsa002核酸属于线型双链DNA,具有正多面体的头部及较长的尾部,qdsa002头部直径约47 nm,尾长约120 nm,属肌尾噬菌体科。生理学特征研究结果表明,qdsa002在双层平板上能形成清晰透亮的噬菌斑。最佳感染复数为0.001,潜伏期40 min,爆发期180 min。在pH为5~9范围内能够保持良好的生理活性。在温度不高于50 ℃条件下裂解性能良好。该噬菌体可用于防治MRSA感染的生物制剂。
耐加氧西林金黄色葡萄球菌噬菌体,酶切,鉴定,生理学性质
耐甲氧西林金黄色葡萄球菌(methicillin-resistantStaphylococcusaureus,MRSA)自1961年在英国被首次发现之后[1],便成为临床感染最为重要的病原菌之一。MRSA引起的感染死亡率很高,造成了很严重的临床医学及公共卫生问题。目前对MRSA的治疗主要采用万古霉素,但是近几年的研究发现已有对万古霉素耐药、中介耐药及异质性耐药的MRSA菌株产生[2],这无疑对于治疗MRSA感染提出了一个新的难题与挑战。
噬菌体是一种感染细菌、真菌和放线菌的病毒,具有以下特点:专一性高、安全性好、增殖能力强和来源范围广等优点[3],是生物防治致病菌很好的一种方法。近年来噬菌体应用于腐败菌或食源性致病菌的相关研究已有很多,且取得了较好的控制作用。例如Suanne Guenther等[4]利用噬菌体对被沙门氏菌感染的巧克力牛奶、土耳其熟食肉品、水产食品及热狗进行抑菌处理后发现,温度为8 ℃时沙门氏菌不生长,温度为15 ℃时沙门氏菌数有3~5个数量级的下降。另外,Pilar Garcia等人用制备的混合噬菌体对奶酪中的金黄色葡萄球菌进行抑制,裂解性噬菌体可以在25 ℃的温度条件下作用4 h后,奶酪中的金黄色葡萄球菌可基本被消除,表明噬菌体在奶酪加工过程中能起到较好的生物防控作用[5];本实验通过分离MRSA噬菌体并对其生理学性质进行研究,为噬菌体治疗MRSA感染提供了实验依据。
1 材料与方法
1.1材料与仪器
耐加氧西林金黄色葡萄球菌43300(简称S.aureus43300)从美国典型微生物菌种保藏中心购买;UNIP-10柱式细菌基因组提取试剂盒上海生工;营养肉汤液体培养基、营养琼脂固体培养基北京陆桥技术有限责任公司;氯化钠、水合硫酸镁等试剂均为分析纯;半固体由体积比为1∶1的营养肉汤液体培养基和营养琼脂固体培养基配制而成;实验用水为新鲜纯水。
DYY-TC型电泳仪北京市六一仪器厂;JEM-1200EX透射式电子显微镜JEOL公司;XMT-152A电热恒温培养箱宁波机电工业研究设计院;HZQ-F100振荡培养箱哈尔滨市东联电子技术开发有限公司;SW-CJ-2FD超净工作台苏州安泰空气技术有限公司;SiGMa离心机北京思达兴业仪器有限公司;TDL-5M台式大容量冷冻离心机湘仪离心机仪器有限公司;BS2248电子天平北京赛多科斯仪器系统有限公司;Forma Scientific超低温冰箱东莞市昊昕仪器设备有限公司。
1.2实验方法
1.2.1噬菌体的分离将采集的污水经5000 r/min 离心10 min后,取上清液5 mL加入到双倍浓缩的营养肉汤液体培养基中,同时加入100 μL对数期的S.aureus43300(S.aureus43300接种到营养肉汤培养基中,37 ℃条件下培养12 h可达到对数期),37 ℃下150 r/min振荡培养12 h。然后经5000 r/min离心10 min后,取上清液过0.22 μm的滤膜,得到噬菌体原液,于4 ℃冰箱中保存。
1.2.2噬菌体的纯化采用双层平板法[6],具体操作如下:用灭菌的SM缓冲液将噬菌体原液稀释1~6倍,取100 μL稀释的噬菌体液和100 μL对数期的S.aureus43300加入到5 mL半固体中,混合均匀后倒入固体培养基中,半固体凝固后放于37 ℃培养箱中倒置恒温培养12 h。
在上述出现噬菌斑的平板中,用接种丝挑取单个噬菌斑到1 mL SM缓冲液中混均,4 ℃保存12h。然后用无菌SM缓冲液对上述噬菌体液稀释1~6倍后,取100 μL稀释的噬菌体液和100 μL对数期的S.aureus43300加入到5 mL半固体培养基中,混合均匀后倒入固体培养基中,半固体凝固后放于37 ℃培养箱中倒置恒温培养12 h。重复上述操作5次以上,待得到的噬菌斑形态大小一致后即得到了纯噬菌体。
1.2.3噬菌体的增殖采用液体增殖法、固体增殖法[7]。液体增殖法:取100 μL对数期的S.aureus43300和100 μL噬菌体加入到营养肉汤培养基中,37 ℃条件下150 r/min培养8 h后,将上述液体在5000 r/min条件下离心10 min,取上清液用双层平板法测定效价,如果效价高(≥108pfu/mL),则将液体过0.22 μm滤膜得到高效价噬菌体,在4 ℃冰箱中保藏,如果效价低(<108pfu/mL),则继续将噬菌体进行增殖。
固体增殖法:取稀释1~6倍后的噬菌体液制成双层平板,于37 ℃恒温倒置培养12 h后,选取噬菌斑最多但未通透的平板,加入10 mL SM缓冲液,在4 ℃中过夜。然后将SM缓冲液及双层平板中的上层取到无菌离心管中,8000 r/min离心15 min,取上清液测定噬菌体的效价,如果效价高则将上清液过0.22 μm滤膜得到高效价噬菌体,在4 ℃冰箱中保藏。
1.2.4噬菌体核酸的提取和酶切鉴定取固体增殖后效价达109pfu/mL的噬菌体液1 mL在20000 r/min条件下离心1 h,将上清液去掉,采用UNIP-10柱式细菌基因组提取试剂盒提取噬菌体的核酸。
将噬菌体核酸与限制性内切酶HindⅢ 37 ℃酶切1 h,用1%琼脂糖凝胶电泳鉴定酶切结果,其中1%琼脂糖凝胶制备方法为称取0.3 g琼脂糖加入3 mL 5×TBE缓冲液和27 mL超纯水。电泳时电压设置为90 V,时间30min[8]。
1.2.5噬菌体的电镜观察采用磷钨酸负染法[9],取100 μL噬菌体液滴在石蜡上,将铜网置于噬菌体液上,5 min后将铜网取下放在滤纸片上自然晒干5 min,然后用2%的磷钨酸染色10 min后,取下铜网晒干,在透射电镜(JEM-1200EX)下进行观察。
1.2.6噬菌体最佳感染复数(MOI)的测定按照感染复数为10-7、10-6、10-5、10-4、10-3、10-2、10-1、100、101,将100 μL噬菌体和100 μLS.aureus43300加入到5 mL液体培养基中,37 ℃条件下150 r/min振荡培养8 h,将上述液体在8000 r/min离心15 min,取上清液用双层平板法测定噬菌体效价。效价最高的感染复数为噬菌体的最佳感染复数。
1.2.7噬菌体的一步生长曲线采用Lu等[10]方法进行改进,以感染复数为10,将1 mL噬菌体和1 mL对数期的S.aureus43300加入到100 mL液体培养基中,37 ℃培养5 min后,将上述液体12000 r/min离心30 s,弃上清。然后用营养肉汤液体培养基洗剂两次,除去未吸附的噬菌体,然后加入等量预热的营养肉汤液体培养基混匀,立即置于37 ℃条件下150 r/min振荡培养。同时开始计时,以0时刻和每隔10 min取样一次测定噬菌体效价。以时间为横坐标,噬菌体效价为纵坐标绘制一步生长曲线。
1.2.8噬菌体pH稳定性配制pH为2、3、4、5、6、7、8、9、10、11、12、13的营养肉汤液体培养基,分别取900 μL加入到无菌EP管中,放于37 ℃水浴锅中,当温度平衡后加入100 μL噬菌体,恒温水浴2 h后用双层平板法噬菌体效价。
1.2.9噬菌体的热稳定性将400 μL噬菌体加入到无菌EP管中,在40、50、60、70、80 ℃水浴作用20、40、60 min后,置于4 ℃冷却,然后用双层平板法测定噬菌体效价。
1.2.10数据处理使用作图软件Origin进行数据相关的处理。
2 结果与分析
2.1噬菌体的分离、纯化
从团岛污水处理厂采集到的污水中分离得到了一株噬菌体,编号为qdsa002,在双层平板上经过12 h培养后形成噬菌斑如图1所示。

图1 噬菌体qdsa002噬菌斑形态Fig.1 The bacteriophage plaque of phage qdsa002
可以看出,噬菌斑中间透亮且边缘清晰,呈现出烈性噬菌体的噬菌斑特征[11]。
噬菌体在自然界中广泛存在,且数量巨大,超过1031。噬菌体分为烈性噬菌体和温和性噬菌体,烈性是菌体进入宿主菌后能够裂解宿主菌,而温和性噬菌体却不能引起宿主菌的裂解[12]。噬菌体的分离与季节有较大关系,主要是温度能够影响到宿主菌的数量进而影响到噬菌体的数量。本研究中的噬菌体分离于2014年8月,属于秋季分离到的噬菌体,双层平板法培养24 h才能出现噬菌斑,与李雪玲等[13]分析的结果一致。
2.2噬菌体核酸的提取和酶切鉴定
噬菌体核酸提取和酶切鉴定结果如图2所示,可以看出提取的噬菌体浓度与纯度都较好,且噬菌体核酸能被HindⅢ酶切成三条片段,说明与HindⅢ酶有两个酶切位点,同时表明噬菌体核酸类型为双链DNA。

图2 噬菌体核酸噬类型的判断Fig.2 Determination of the nucleic acid type of bacteriophage注:M. λ-Hind Ⅲ digest DNA分子量标准;A. 噬菌体核酸;B. 噬菌体核酸Hind Ⅲ酶切。
本研究中使用了许多种限制性内切酶对qdsa002核酸进行酶切,只有HindⅢ酶切条带较为清晰可辨,而电泳条带的清晰度与很多因素有关如电压大小、琼脂糖凝胶浓度、电泳时间等[14]。具体实验条件需要具体分析和探索。
2.3噬菌体的电镜观察
噬菌体负染后经过透射电镜观察结果如图3所示。

图3 噬菌体qdsa002透射电镜照片(×128k)Fig.3 Electron micrograph of phage qdsa002(×128k)
可以看出,噬菌体为典型的T形噬菌体。头部呈正多面体结构,有细长的尾部。噬菌体全长168 nm,头部直径47 nm左右,尾部长度120 nm。
根据2005年国际病毒分类委员会(ICTV)第八次报告[15],细菌病毒被划为了五个级阶:群、目、科、属、种,细菌病毒有了系统的分类。本研究的噬菌体头部呈正多面体结构,全长168 nm,头部直径47 nm左右,尾部长度120 nm,为典型的肌尾噬菌体科与已报道的金黄色葡萄球菌噬菌体基本一致[16]。
2.4噬菌体最佳感染复数(MOI)的测定
qdsa 002的最佳感染复数测定结果见表1。从表1可以看出,感染复数从10到0.000001,噬菌体效价表现为先升高后降低的趋势,当感染复数为0.001时噬菌体效价最高为2.23×108,表明噬菌体qdsa 002最佳感染复数为0.001。

表1 噬菌体qdsa002最佳感染复数测定
感染复数是研究病毒感染与产出之间量效关系而提出的一个重要生物学指标。感染复数不同,子代噬菌体的产率也会有较大的差异,噬菌体产率最高的感染复数为最佳感染复数。这一生理学指标为后续噬菌体与细菌用量比提供了一个依据。在本研究中,噬菌体qdsa 002最佳感染复数为0.001,曾志良等[17]分离得到的金黄色葡萄球菌噬菌体最佳感染复数为0.01,李陇平等[18]分离得到的金黄色葡萄球菌噬菌体最佳感染复数也是0.01,与之相比较qdsa 002具有较强的裂解能力。
2.5噬菌体的一步生长曲线
噬菌体qdsa002的一步生长曲线见图4。从图4可以看出,前40 min内,噬菌体的数量并没有增加,表明噬菌体的潜伏期为40 min,在随后的180 min内,噬菌体的数量急剧增加,说明噬菌体的爆发期约180 min,之后噬菌体趋于稳定,进入稳定期。
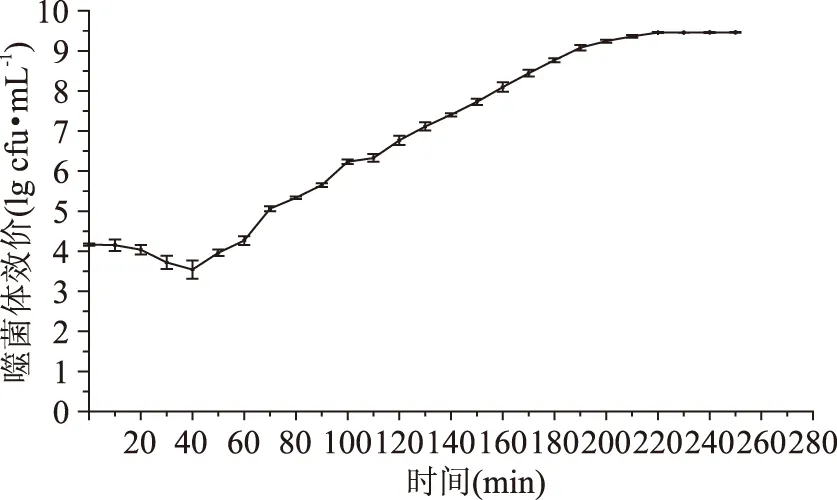
图4 噬菌体qdsa002一步生长曲线Fig.4 One-step growth curve of phage qdsa002
噬菌体一步生长曲线表现为潜伏期、裂解期和稳定期,不同噬菌体的潜伏期和裂解期也会存在一定的差异。噬菌体与宿主菌吸附开始潜伏期;噬菌体从宿主菌内释放,表明潜伏期结束,裂解期开始;宿主菌裂解死亡噬菌体数量保持平稳,说明裂解期结束进入稳定期。本研究的噬菌体qdsa002潜伏期为40 min,爆发期为180 min,与顾敬敏[19]分离得到的金黄色葡萄球菌噬菌体GH15相比(潜伏期为25 min),潜伏期明显加长。
2.6噬菌体pH稳定性
噬菌体pH稳定性测定结果见图5。可以看出,噬菌体qdsa002在5~9范围内噬菌体效价与初始效价相比没有明显差异,能够保持良好的裂解活性;当pH为4和10时,噬菌体效价能够保持在107pfu/mL以上,此时仍能保持噬菌体较好的活性;当pH=11时,噬菌体效价依然保持在106pfu/mL以上;当pH≤3和pH≥12时,噬菌体基本完全失活。

图5 噬菌体qdsa002 pH稳定性Fig.5 The pH stability of phage qdsa002
噬菌体对宿主菌的吸附要受到pH的影响,表现为噬菌体空间构象会受到pH的影响。本研究中,qdsa002在pH为5~9范围内均能保持良好的生理活性,即最适pH为5~9。与贾静静[20]研究的噬菌体最适pH6~8相比,qdsa002的最适pH范围更广。
2.7噬菌体的热稳定性
噬菌体qdsa002热稳定性测定结果见图6。可以看出,噬菌体qdsa002在40 ℃和50 ℃条件下作用1 h效价基本保持不变;噬菌体在60 ℃下作用20 min效价降到105pfu/mL,作用1 h后效价只有102pfu/mL;在70 ℃下作用20 min效价就降到不足10 pfu/mL,40 min后检测不到噬菌体效价;在80 ℃下作用20 min噬菌体就基本全部失活。说明噬菌体qdsa 002在50 ℃及其以下温度能够保持良好的活性。

图6 噬菌体qdsa002的热稳定性Fig.6 The thermal stability of phage qdsa002
温度也能影响到噬菌体对细菌的吸附,过高的温度会影响到噬菌体的空间构象使其失去裂解活性。在本研究中,噬菌体qdsa002在50 ℃及其以下的温度能够保持良好的活性,与蔡天舒研究的金黄色葡萄球菌噬菌体qdsa001相比,耐热性基本一致。
3 结论
以S.aureus43300为宿主菌,从青岛市团岛污水处理厂的污水中分离纯化出一株噬菌体qdsa002,并对其它生理生化特性进行了研究,得到的结论如下:
3.1电镜观察表明该噬菌体qdsa002头部为正多面体结构,有细长的尾部,呈蝌蚪形。头部直径大约为47 nm左右,尾部长约120 nm左右,属于典型的肌尾噬菌体。通过对噬菌体基因组提取和鉴定表明,其核酸类型为DNA。
3.2噬菌体对50 ℃以下的温度具有较高的耐受力,60 ℃温度作用20 min后,其效价仍在105pfu/mL以上,在70 ℃下作用20 min效价就降到不足10 pfu/mL,而在高于80 ℃以上的高温时,将会失去裂解细菌的活性。噬菌体qdsa002最适pH为5~9,当pH在3以下及12以上时,会完全失去感染宿主菌的能力。对噬菌体的感染复数的研究表明,噬菌体和宿主菌的浓度比为0.001时,所得到的子代噬菌体的效价最高。另外,其一步生长曲线显示噬菌体感染宿主菌的潜伏期为40 min,裂解期约为180 min。综上所述,噬菌体qdsa002具有较为良好的生物学特性,为应用提供了良好材料。
[1]Matouskova I,Janout V. Current knowledge of methicillin-resistantStaphylococcusaureusand community-associated methicillinresistantStaphylococcusaureus[J]. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub,2008,152(2):191-202.
[2]Liu C,Bayer A,Cosgrove SE,et al. Clinical Practice Guidelines by the Infectious Diseases Society of America for the Treatment of Methicillin-ResistantStaphylococcusaureus
Infections in Adults and Children[J].Clinical Infectious Diseases,2011,52(3):e18-e55.
[3]Li M,Han F,Lin H,et al. Isolation,identification and physiological characterization of lytic phage against bacteriumSalmonellasp[J]. Fisheries Science,2013,32(9):531-535.
[4]Guenther S,Herzig O,Fieseler L,et al. Biocontrol of Salmonella Typhimurium in RTE foods with the virulent bacteriophage FO1-E2[J]. Int J Food Microbiol,2012,154(1-2):66-72.
[5]Pilar Garcia,Carmen Madera,Beatriz Martinez,et al. Biocontrol ofStaphylococcusaureusin curd manufacturing processes using bacteriophages[J]. International Dairy Journal,2007,17:1232-1239.
[6]余茂効,司穉东. 噬菌体实验技术[M]. 北京:科学出版社,1991:10.
[7]丁云娟. 副溶血弧菌噬菌体qdvp001的分离鉴定及其在牡蛎净化中的初步应用[D]. 青岛:中国海洋大学,2012.
[8]李萌. 宽裂解谱沙门氏菌噬菌体的基因组学分析及其重组内溶素抑菌活性研究[D]. 青岛:中国海洋大学,2014.
[9]宁淑香,聂丽平,陆敏. VP10-一株海洋噬菌体的分离及特性研究[J]. 水产科学,2003,19(2):14-16.
[10]Lu Z,Breidt FJ,Fleming HP. Isolation and characterization of aLactobacillusplantarumbacteriopHage ΦJL21 from a cucumber fermentation[J]. Food Microbiol,2003,84(2):225-235.
[11]汤芳,陆承平. 猪链球菌的烈性噬菌体和前噬菌体[J]. 微生物学报,2015,55(4):389-394.
[12]彭勇. 副溶血弧菌噬菌体的分离鉴定、裂解性能及其在副溶血弧菌快速检测中的初步应用[D]. 青岛:中国海洋大学,2013.
[13]李雪玲,冯惠玲,李锡平,等. 金黄色葡萄球菌噬菌体的分离筛选[J]. 食品工业科技,2013,15:158-161,165.
[14]刘永平,罗志勇. 琼脂糖凝胶电泳实验技巧[J]. 实用预防医学. 2006,13(4):1068-1096.
[15]洪健,周雪平. ICTV 第八次报告的最新病毒分类系统. 中国病毒学,2006,21(1):84-96.
[16]蔡天舒. 金黄色葡萄球菌噬菌体qdsa001的生理学特性及初步应用[D]. 青岛:中国海洋大学,2013.
[17]曾志良,伍绮文,赵晓亚,等. 一株金黄色葡萄球菌噬菌体的分离及其裂解酶的原核表达[A]. 中国畜牧兽医学会动.物微生态学分会:2014:1.
[18]李陇平,张智英. 金黄色葡萄球菌烈性噬菌体的分离鉴定和最佳保存方法研究[J]. 中国畜牧兽医,2011(6):141-146.
[19]顾敬敏. 金黄色葡萄球菌噬菌体GH15及其裂解酶三维结构与分子作用机制研究[D]. 长春:吉林大学,2014.
[20]贾静静. 金黄色葡萄球菌噬菌体的分离及其裂解性能的研究[D]. 无锡:江南大学,2012.
Identification and biological properties of a methicillin-resistantStaphylococcusaureusphage qdsa002
JU Lei,WANG Jing-xue*,LIN Hong,LI Meng-zhe,MAO Xiang-zhao
(Food Safety Laboratory,College of Food Science and Engineering,Ocean University of China,Qingdao 266003,China)
In order to develop an effective bio-controlling method against methicillin-resistantStaphylococcusaureus,one phage qdsa002 was isolated from sewage water gathered from Tuandao sewage treatment plant of Qingdao. Bacteriophage was identified by plaque shapes,restriction endonucleases analysis,electronic microscope technique. The physiological characteristics of the optimum MOI value,the growth curve,the pH value and the thermal stability were assayed. Results showed that the phage qdsa002 had linear double stand DNA and regular polyhedron symmetric head. The qdsa002 had a head of about 47 nm in diameter and tails of 120 nm,belonging to family Siphoviridae. The physiological characteristics showed plaques with transparent center in double agar plate. The optimum MOI value of qdsa002 was 0.001,one-step growth experiment showed that the latent time and burst time were 40 min and 180 min,respectively. The qdsa002 was stable over a wide range of pH(5~9)and temperature(≤50 ℃). The phage can be used in the prevention and treatment of MRSA infection.
methicillin-resistantStaphylococcusaureusphage;restriction endonucleases analysis;identification;physiological characteristics
2015-12-25
鞠磊(1989-),男,硕士研究生,主要从事食品质量与安全方面的研究,E-mail:juleiouc@163.com。
王静雪(1976-),教授,主要从事食品质量与安全、应用微生物方面的研究,E-mail:snow@ouc.edu.cn。
“十二五”农村领域国家科技支撑计划(2015BAD16B0902);山东省科技重大专项(2014ZZCX02703)。
TS201.3
A
1002-0306(2016)16-0230-05
10.13386/j.issn1002-0306.2016.16.037
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年2期)2022-03-29
中学生数理化(高中版.高考数学)(2021年11期)2021-12-21
中国动物传染病学报(2021年3期)2021-07-21
中学生数理化(高中版.高二数学)(2021年4期)2021-07-20
中学生数理化(高中版.高二数学)(2021年4期)2021-07-20
科学24小时(2020年4期)2020-05-14
新世纪智能(数学备考)(2020年12期)2020-03-29
实用口腔医学杂志(2017年6期)2017-09-19
中华老年口腔医学杂志(2016年5期)2016-03-01
