
广谱抗性基因Pi9在黑龙江省水稻品种中的分布
2016-10-26 01:33邹德堂姜思达赵宏伟郭丽颖孙健刘化龙辛威
东北农业大学学报 2016年7期
邹德堂,姜思达,赵宏伟,郭丽颖,孙健,刘化龙,辛威
(东北农业大学农学院,哈尔滨 150030)
广谱抗性基因Pi9在黑龙江省水稻品种中的分布
邹德堂,姜思达,赵宏伟,郭丽颖,孙健,刘化龙,辛威
(东北农业大学农学院,哈尔滨150030)
2014年利用显性标记pB8对黑龙江省96份水稻主栽品种筛选Pi9基因,对供试材料作苗期和分蘖期抗病鉴定,明确Pi9基因在黑龙江省水稻品种中分布情况及应用价值。结果表明,供试材料中,有早熟青森、垦粳1等44个品种含有Pi9基因,苗期和分蘖期平均发病级别均为2.4级;青森5号、长白9号等52个品种不含Pi9基因,苗期和分蘖期平均发病级别分别为5.1和5.4级。研究结果可为黑龙江省水稻品种改良以及抗病资源合理利用奠定基础。
水稻;分子标记;辅助选择;稻瘟病;Pi9基因
网络出版时间2016-7-20 10:40:03[URL]http://www.cnki.net/kcms/detail/23.1391.S.20160720.1040.004.html
邹德堂,姜思达,赵宏伟,等.广谱抗性基因Pi9在黑龙江省水稻品种中的分布[J].东北农业大学学报,2016,47(7):1-8.
Zou Detang,Jiang Sida,Zhao Hongwei,et al.Distribution of broad-spectrum resistance gene-Pi9 in the rice cultivars of Heilongjiang Province[J].Journal of Northeast Agricultural University,2016,47(7):1-8.(in Chinese with English abstract)
水稻在世界粮食作物中有重要作用[1]。近年来,水稻育种成效显著[2]。传统水稻育种方式易受外界因素干扰,致使优良基因流失且效率较低。随着分子生物学发展,分子标记辅助选择技术日渐成熟[3]。分子标记辅助选择技术利用与目标基因紧密连锁的标记在DNA水平上筛选,不受等位基因显隐性干扰,大幅提高育种效率,在提高水稻抗性、品质等方面均起重要作用[4],为现代育种开辟新途径。目前,水稻等多个作物测序工作已完成,相关数据库建立,分子标记数目和类型不断增加,分子标记辅助选择育种广泛应用[5]。
稻瘟病对全球水稻构成严重威胁[6],造成水稻减产高达15%~30%,严重制约水稻高产和稳产。稻瘟病生理小种繁多、变异速度快等特点增加稻瘟病防治困难。发掘抗病基因,培育抗病品种是防治稻瘟病经济有效方式。充分利用抗病基因,对栽培品种合理布局,发挥水稻自身优势解决病害问题起到重要作用[7]。
Pi9基因在小粒野生稻中发现并命名,被定位于水稻第6条染色体短臂上[8],是已克隆的抗稻瘟病主效基因之一,该基因能抵御来自多个国家和地区多数稻瘟病生理小种侵染,不留过敏病斑。雷财林等研究认为,Pi9基因在黑龙江省对稻瘟病菌株抗谱较广[9]。本研究通过对黑龙江省主栽的96个水稻品种分子标记辅助选择,检测Pi9基因在所选品种中分布情况。对所选品种在苗期和分蘖期作抗病性鉴定试验,将分子标记辅助选择结果与表型数据结合,明确Pi9基因在黑龙江省分布现状及其在提高水稻抗病性方面价值,为黑龙江省水稻合理布局及抗稻瘟病品种选育提供依据。
1 材料与方法
1.1实验材料
试验材料为96份黑龙江省主栽粳稻品种,75-1-127单基因(Pi9)系作为阳性对照;丽江新团黑谷为阴性对照。所有供试稻株均由东北农业大学水稻研究所提供。
1.2PCR检测
1.2.1DNA提取
采用CTAB法[10]提取供试材料DNA。
1.2.2供试引物
pB8引物序列见表1[11]。引物由上海生工生物工程技术服务有限公司合成。
1.2.3PCR扩增
PCR反应总体积为20 μL,其中DNA 3 μL(25 ng·μL-1),10×PCR缓冲液2 μL,25 mmol·L-1MgCl2溶液2 μL,10 mmol·L-1dNTP 0.2 μL,Taq酶0.3 μL,引物2 μL。加入ddH2O至20 μL。加入液体石蜡封盖体系。
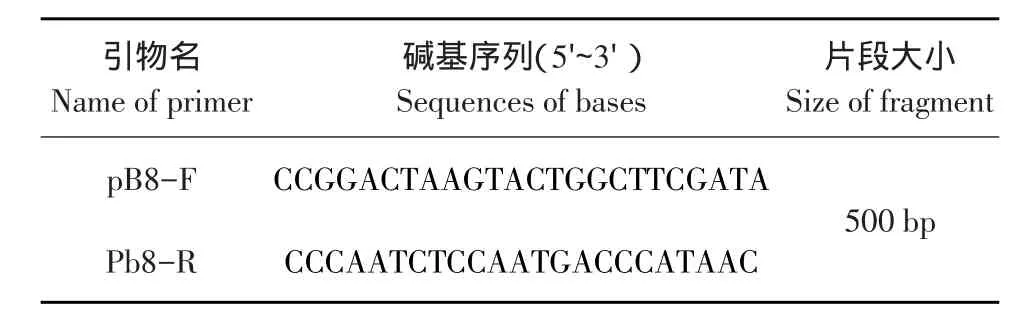
表1 引物序列与相关信息Table 1Primer sequences and related information
PCR扩增经94℃预变性6 min;94℃30 s,55℃30 s,72℃30 s,共包含35个循环;72℃延伸5 min[12],扩增产物于4℃条件下保存。所得PCR扩增产物经1%琼脂糖凝胶电泳,利用溴化乙锭染色后紫外灯下观察[13]。
1.3瘟病抗性接种鉴定
1.3.1供试菌株
供试菌株为ZA5,ZA5对黑龙江省主栽品种具有较强致病率(63.89%)[14],供试菌株由东北农业大学水稻研究所保存、提供。
1.3.2接种鉴定
苗期接菌鉴定于2014年在东北农业大学温室内完成,采用秧盘种植,选取饱满健康种子,每穴播种10粒,长至3叶1心时利用ZA5菌株孢子悬浮液喷雾,使液滴均匀分布于叶片及茎秆。喷雾接菌后,塑料薄膜覆盖保湿,遮光处理24 h后置于温室中培养,控制温度与湿度。于处理后7~10 d,发病情况稳定后调查发病级别。根据病斑形状、大小和数量,以国际水稻所叶瘟分级标准(见表2)[15]作为参照,调查水稻叶瘟发病级别,计算平均值。
分蘖期接菌鉴定于同年在东北农业大学香坊实验实习基地试验田进行,4月16日播种,5月20日移栽,田间随机区组排列,每份材料种植1行,每行20株,株行距13.3 cm×30 cm,每穴1株,田间管理按常规进行。分蘖盛期从每行中选取3株,每株3个分蘖,采用注射接菌方式,离颈基部20 cm处人工注射ZA5生理小种,接菌量以心叶有水滴溢出为止,每稻杆约0.5 mL。对已注射过稻杆作标记。接种7~10 d后,按照国际水稻所叶瘟发病级别调查标准调查,统计发病级别并计算平均值。
1.3.3稻瘟病抗病级别及评价标准计算
将所选群体按照是否携带Pi9基因标准分为两部分,选取苗期和分蘖期两个时期,分别计算两部分平均发病级别,并统计发病区间。按照表2评价标准统计两部分品种各时期发病情况,得出两部分品种从高抗到高感各级别品种数量。同时计算两部分品种苗期和分蘖期抗病率。通过以上计算结果鉴定Pi9基因在黑龙江省主栽品种中的抗病性价值。
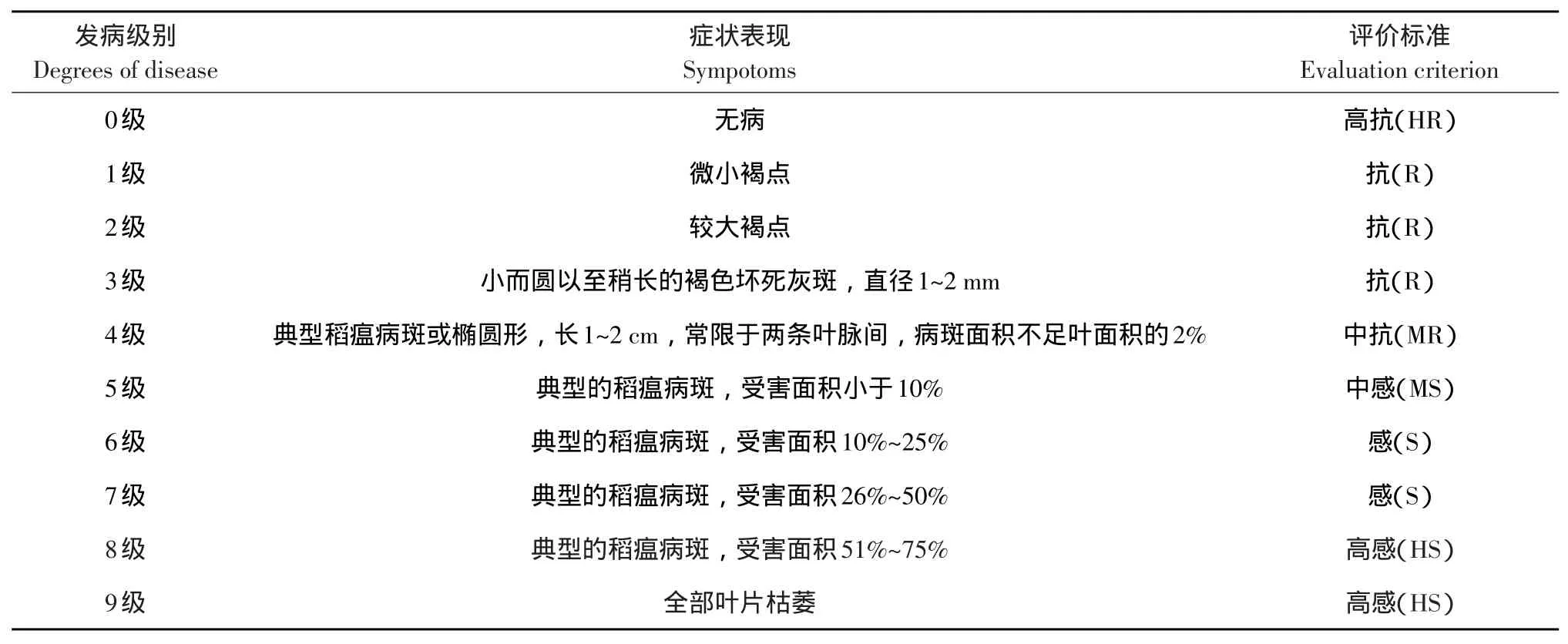
表2 国际水稻所叶瘟发病级别调查标准Table 2Evaluation criterion of rice leaf blast from the International Rice Research Institute
2 结果与分析
2.1广谱抗病基因Pi-9在黑龙江省主栽品种中分布情况
将选取的96份寒地粳稻品种以pB8为标记PCR扩增,以75-1-127为阳性对照,丽江新团黑谷为阴性对照,携带Pi9基因的品种500 bp处扩增出条带,结果见表3。从表中可知,所选96个品种中有44个品种扩增出目标条带,如早熟青森、垦粳1等,说明这44个品种携带Pi9基因;其余52个品种未扩增出目标条带,Pi9基因在所选品种中占有率达45.8%。图1为所选试验材料扩增结果。
2.2稻瘟病抗性鉴定
利用强致病力稻瘟病生理小种ZA5接种供试材料,结果见表3。由表3中结果经计算可得,75-1-127苗期和分蘖期病害级别分别为1.7和2.5级,表现为抗病;感病对照丽江新团黑谷苗期和分蘖期平均发病级别分别为8.1和9.0,均达到高感。
所选96份主栽品种中,对携带Pi9基因的供试材料抗病性结果分析可得,其苗期和分蘖期两时期发病较低,叶瘟整体发病级别在0~5.3级和0~4.2级,平均发病级别均为2.4级,表现为抗病。相同时期不含Pi9基因供试材料叶瘟发病级别在0~8.6级和0~9.0级,平均发病级别分别为5.1和5.4级,表现为中感。对44个品种分别在苗期和分蘖期进行抗病性评价标准鉴定,结果表明,苗期有高抗品种8个,抗病品种24个,中抗品种10个,中感品种2个;分蘖期有高抗品种3个,抗病品种34个,中抗品种7个。
对不含Pi9基因供试材料分析可得,其苗期和分蘖期两个时期叶瘟发病级别在0~8.6级和0~9.0级,平均发病级别分别为5.1和5.4级,表现为中感。对这52个品种分别在苗期和分蘖期进行抗病性评价标准鉴定,结果表明,苗期有高抗品种1个,中抗品种5个,抗病品种11个,中感品种11个,感病品种17个,高感品种7个;分蘖期有高抗品种3个,抗病品种9个,中抗品种4个,中感品种6个,感病品种20个,高感品种10个。
计算所选材料2014年抗病率[16],携带Pi9基因的44个品种苗期抗病率(%)=42/44×100%= 95.45%,分蘖期抗病率(%)=52/52×100%=100%;不含Pi9基因的52个品种苗期抗病率(%)=6/52× 100%=11.54%,分蘖期抗病率(%)=16/52×100%= 30.77%,携带Pi9基因的品种抗病率明显高于不含Pi9基因品种。
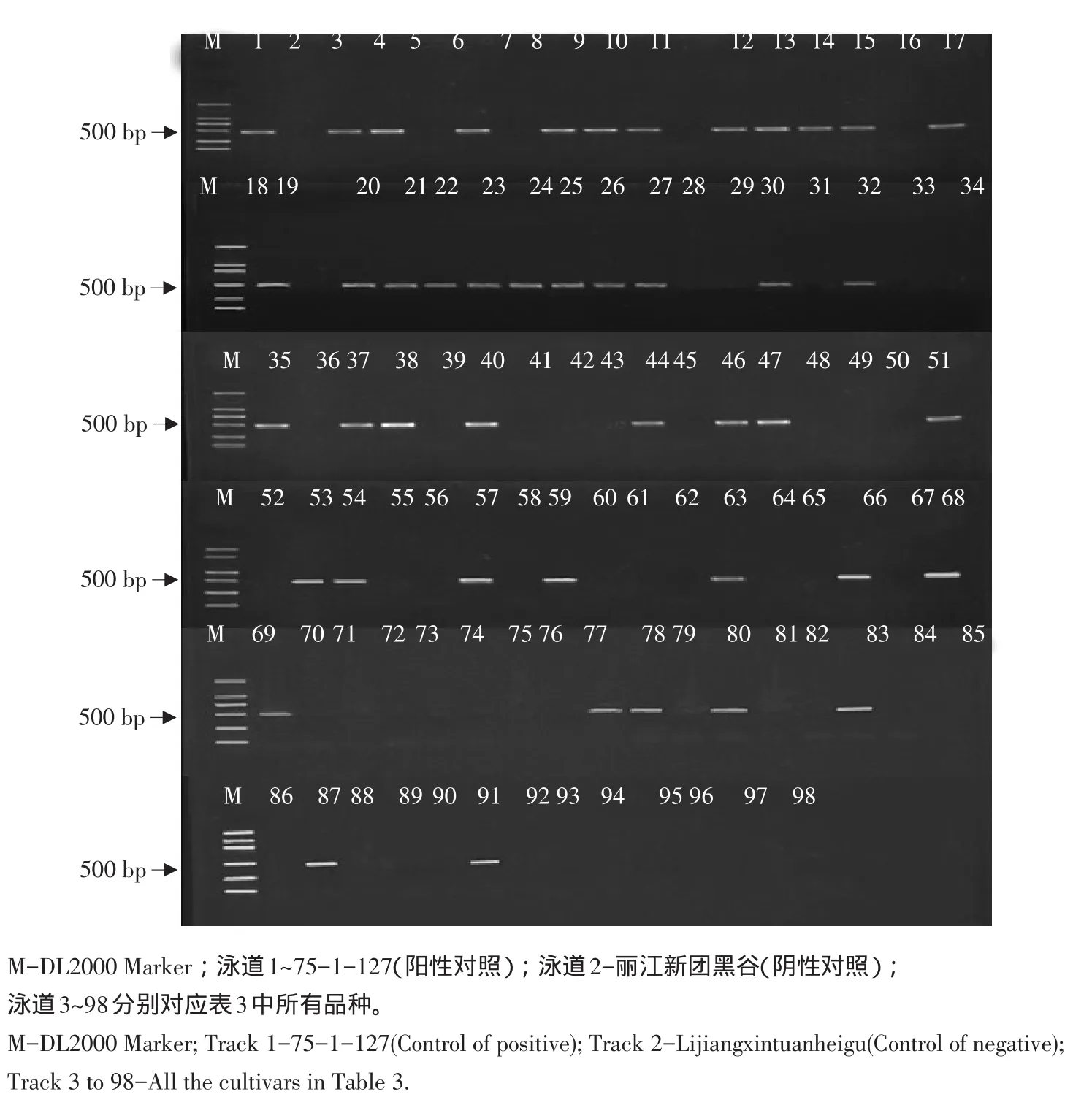
图1 所有品种以pB8为引物扩增结果Fig.1PCR amplifications of pB8 specific markers for all the cultivars

表3 黑龙江省主栽品种Pi9基因分布及抗病性表现Table 3Distribution of Pi9 gene and disease resistance of major cultivars from Heilongjiang Province

续表

注:√代表含有Pi9;×代表不含有Pi9;75-1-127为阳性对照;丽江新团黑谷为阴性对照。Note:√indicates that the cultivars contain the gene of Pi9;×indicates that the cultivars do not contain the gene of Pi9;75-1-127 is control of positive;Lijiangxintuanheigu is control of negative.
3 讨论
分子标记辅助选择在育种方面应用广泛[17],刘华招等筛选2009年黑龙江省种植面积较广的36份品种,最终筛选出4个含有Pib抗性基因和6个含有Pi-ta抗性基因品种[18]。李洪亮等将水稻品种BL6中携带的抗病基因Pi1和Pi2聚合到空育131中,利用已建立的显性分子标记RM144和AP22分别对Pi1和Pi2基因进行分子标记辅助选择,大幅拓宽空育131品种抗谱[19]。
本研究分析Pi9基因在黑龙江省种质资源分布情况,筛选出44份携带Pi9基因的种质资源。为今后育种工作中优良品种选育提供依据,亲本选择时,合理选择携带Pi9基因品种,可提高配置的杂交后代抗病性。对不含Pi9基因,但具有较强抗病性的品种,如东农426、松粳8号等深入研究,加大力度发掘、定位抗病基因,探索抗病机理。在所选材料中存在苗期和分蘖期抗病性均优于阳性对照75-1-127的品种,比如龙联1号,北稻5号等。此类品种可能携带除Pi9之外的抗稻瘟病基因,因此加强对此类品种抗病基因发掘与研究,对黑龙江省水稻抗病资源拓展有重要意义。
本研究可知,Pi9基因在黑龙江省水稻抗病育种中具有很高应用价值,在黑龙江省乃至全国范围内加强对Pi9基因推广和应用意义重大。但有些不含Pi9基因的供试材料如东农426、松粳8号等表现较强稻瘟病抗性,说明这些品种中可能含有其他抗性基因,有待进一步发掘和利用。同时,试验材料中存在苗期和分蘖期抗病性均较强的品种,如龙庆稻1号、龙粳14号、东农426等。加大以上品种开发力度,发掘其中抗病基因将会为黑龙江省水稻抗病育种带来突破性进展。
通过比较苗期和分蘖期稻瘟病抗性发现,供试材料中一部分苗期和分蘖期抗病水平不一致,如石狩白毛苗期发病为5.3级,表现为中感,而分蘖期表现高抗(0级)。梧农71也有类似表现;东农421苗期发病为0.9级,表现为高抗,而分蘖期表现高感(8.2级)。由此看出,水稻苗期和分蘖期稻瘟病抗性差异显著,结果表明,相同抗病基因不同生育期表达差异显著。从表3中可以看出,相同生育时期,含有Pi9基因的供试材料其稻瘟病抗性差异显著,如苗期的龙盾106和龙粳香1号,平均抗病级别均相差2级,分别表现为高抗和抗性;分蘖期龙稻12号和垦粳1号,平均抗病级别均相差2.6级,分别表现为抗病和中感。结果表明,Pi9基因在不同遗传背景中抗病效果不同。
所选材料中存在含有Pi9基因,是对稻瘟病抵抗能力较弱品种。说明虽然Pi9基因对稻瘟病具有较强抵抗能力,但外界环境等因素影响,稻瘟病生理小种繁多等特点会限制抗性基因作用[20]。Pi9基因抗谱广,但不能对所有菌株产生抗病效果。杨婷婷等曾证明Pi9基因对来自韩国的ROR1和日本的KOH两个菌株无抗性[21]。因此在水稻生产中,不能仅依赖单一抗病基因或优势,培养并选育具有综合优良性状品种。考虑多个基因综合应用,利用多个优良基因互补作用拓宽抗谱,培育具有持久抗性品种。陈学伟等将Pid(t)1、Pi-b、Pi-ta2聚合到G46B品种中,使该品种抗病性得到有效改善[22];陈圣等将X23、Pi9和Bt基因聚合到同一株系中,这三种基因分别对水稻白叶枯病、稻瘟病和水稻螟虫具有很高抗性,聚合结果显示,所得株系能同时抵抗这三种逆境[23];阳海宁等将抗水稻白叶枯病基因Xa23和抗稻飞虱基因Bph3导入优良杂交品种保持系中,获得双基因聚合品种明显改善两种病害抗性[24]。
水稻基因组研究迅速发展,在现有基础上加强对新抗源发掘,有针对性将优良基因导入、聚合到各地区主栽品种中,可为水稻育种开辟新途径。
[1]李永宾.水稻稻瘟病的发生特点及防治技术[J].现代农业科学,2013,44(14):147-150.
[2]Qu S,Liu G,Zhou B,et al.The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice[J].Genetics,2006,172:1901-1914.
[3]王麟.分子标记辅助选择及其在水稻育种中应用与展望[J].黑龙江农业科学,2009(1):140-143.
[4]冯建成.分子标记辅助选择技术在水稻育种上的应用[J].中国农学通报,2006,22(2):43-47.
[5]朱玉君,樊叶杨,黄得润,等.分子标记辅助选择在水稻育种中的应用[J].核农学报,2012,26(5):756-761.
[6]刘摇浩,陈志强,王加峰.水稻NBS-LRR类抗稻瘟病基因研究进展[J].江苏农业学报,2014(3):664-670.
[7]谢培,邓其明,王世全,等.水稻稻瘟病抗性基因研究进展[J].湖南农业科学,2011(5):79-81.
[8]张羽,李厚华,王保军,等.稻瘟病抗性基因Pi9的功能标记的开发[J].云南农业大学学报:自然科学版,2015,30(4):528-534.
[9]雷财林,张国民,程治军,等.黑龙江省稻瘟病菌生理小种毒力基因分析与抗病育种策略[J].作物学报,2011,37(1):18-27.
[10]Doyle J J,Doyle J L.Isolation of plant DNA from fresh tissue[J]. Focus,1990,12:149-151.
[11]张羽,冯志峰,张晗,等.陕西省水稻种质资源中Pi9基因的分布状况[J].四川农业大学学报,2013,31(2):115-121.
[12]刘欣,郑文静,张少斌.水稻SSR-PCR技术反应体系的优化[J].湖北农业科学,2009,48(7):1540-1542.
[13]陈锋,张士永,朱文银.分子标记辅助选择改良圣稻13和圣稻14的条纹叶枯病抗性[J].中国农业科学,2010,43(16):3271-3279.
[14]郭丽颖,赵宏伟,王敬国,等.黑龙江省稻瘟病菌生理小种鉴定和主栽水稻品种抗病性及遗传多样性分析[J].核农学报,2015,29(8):1444-1454.
[15]刘洋,徐培洲,张红宇,等.水稻抗稻瘟病Pib基因的分子标记辅助选择与应用[J].中国农业科学,2008,41(1):9-14.
[16]张其蓉,宋发菊,田进山,等.长江中下游稻区水稻区域试验品种抗稻瘟病鉴定与评价[J].江苏农业科学,2013(4):90-91.
[17]Bryan G T,Wu K S,Farrall L,et al.A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta[J].Plant Cell,2000,12(11):2033-2046.
[18]刘华招,刘延,刘化龙,等.黑龙江省种植品种中稻瘟病抗性基因Pib和Pita的分布[J].东北农业大学学报,2011,42(4):27-31.
[19]李洪亮,李荣田.稻瘟病抗性基因Pi1和Pi2的聚合及其育种价值分析[J].北方水稻,2010,40(5):5-12.
[20]文婷,梁毅,江南,等.利用Pi9基因序列标记辅助选择改良籼稻稻瘟病抗性[J].湖南农业大学学报,2012,38(3):262-266.
[21]杨婷婷,刘雄伦,谭令辞,等.水稻Pi9基因序列标记的开发及其抗瘟育种应用[J].作物研究,2014,28(3):231-235.
[22]陈学伟,李仕贵,马玉清,等.水稻抗稻瘟病基因Pid(t)1、Pi-b、Pi-ta2的聚合及分子标记选择[J].生物工程学报,2004,20(5): 708-714.
[23]陈圣,倪大虎,陆徐忠,等.分子标记辅助选择聚合X23,Pi9和Bt基因[J].生物学杂志,2009,26(3):7-9.
[24]阳海宁,韦绍丽,李孝琼,等.标记辅助培育水稻抗稻褐飞虱和稻白叶枯病基因聚合系[J].分子植物育种,2010,8(1):11-19.
Distribution of broad-spectrum resistance gene-Pi9 in the rice cultivars of Heilongjiang Province
ZOU Detang,JIANG Sida,ZHAO Hongwei,GUO Liying,SUN Jian,LIU Hualong,XIN wei
(School of Agriculture,Northeast Agricultural University,Harbin 150030, China)
The dominant molecular pB8 was used to identify the presence ofPi9 gene in 96 rice leading cultivars of Heilongjiang Province.Meanwhile,assays for resistance identification were performed among rice seedling stage and tillering stage.In order to realize the distribution and application value ofPi9 gene in the rice cultivars of Heilongjiang Province.The results showed that the Pi9 gene was found in 44 rice cultivars among the materials in this study,such as Zaoshuqingsen, Kendao1,whose average disease degrees of rice blast at seedling stage and tillering stage were both 2.4;the other 55 rice cultivars such as Qingsen5,Changbai9,whose average disease degrees of rice blast at seedling stage and tillering stage were 5.1 and 5.4,did not contain thePi9 gene.These results lay a foundation for the improvement of rice cultivars and rational utilization of rice disease resistance resources in Heilongjiang Province.
rice;molecular markers;marker-assisted seletion;rice blast;Pi9 gene
S511
A
1005-9369(2016)07-0001-08
2016-01-04
黑龙江省重大科技招标项目(GA14B102);“十二五”农村领域国家科技计划课题(2013BAD20B04);东北农业大学校级创新课题(yjscx14049)
邹德堂(1965-),男,教授,博士,博士生导师,研究方向为水稻遗传育种。E-mail:zoudt@163.com
猜你喜欢
今日农业(2022年4期)2022-06-01
中国农业科学(2022年8期)2022-05-16
作物学报(2022年6期)2022-04-08
吉林农业·下半月(2017年7期)2017-07-15
浙江农业学报(2017年1期)2017-05-17
农民致富之友(2017年6期)2017-05-10
上海农业学报(2017年3期)2017-04-10
现代农业(2016年5期)2016-02-28
现代农业(2015年1期)2015-02-28
江苏农业科学(2014年8期)2014-10-23
