
黄土丘陵区刺槐人工林与乡土植物叶片和细根功能性状比较研究
2016-10-26 03:45王红霞温仲明高国雄戚德辉杨士梭
水土保持研究 2016年1期
王红霞, 温仲明,,, 高国雄,, 戚德辉, 郭 茹, 杨士梭
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学 资源环境学院,陕西 杨凌 712100; 3.中国科学院 教育部 水土保持与生态环境研究中心, 陕西 杨凌 712100)
黄土丘陵区刺槐人工林与乡土植物叶片和细根功能性状比较研究
王红霞1, 温仲明1,2,3, 高国雄1,2, 戚德辉2, 郭 茹3, 杨士梭1
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学 资源环境学院,陕西 杨凌 712100; 3.中国科学院 教育部 水土保持与生态环境研究中心, 陕西 杨凌 712100)
植物功能性状是表征植物与环境关系的重要媒介。不同种群间功能性状的差异直接决定其竞争优势,并决定群落的结构和功能,影响整个群落的演替与发展。以黄土丘陵区森林草原带不同立地环境下的刺槐林和乡土植物群落为研究对象,比较二者功能性状随立地环境的变化,进而阐明其对不同环境的适应策略。结果表明:(1) 同一坡向,刺槐林和乡土植物群落的叶N含量、根C含量、根N含量、比叶面积、叶组织密度及比根长之间有显著性差异(p<0.05),而根P含量、根组织密度只在阴坡上有显著性差异(p<0.05)。除叶组织密度、根组织密度、根P含量外,刺槐其他各功能性状值均高于乡土植物群落。在同一坡位,刺槐林叶C含量、叶N含量和根C含量、根N含量、比叶面积、比根长整体上显著高于乡土植物群落(p<0.05);(2) 刺槐林的土壤C含量、土壤P含量在阴阳坡上均低于乡土植物群落,阳坡土壤N含量高于乡土植物群落,阴坡上则相反;在不同坡位上,两者的土壤C含量有显著性差异(p<0.05);(3) 刺槐林和乡土植物群落5 m的土壤含水量变化基本一致。刺槐林下土壤含水量低于乡土植物群落。在干旱贫瘠的环境中,刺槐林具有较高的叶养分含量、根养分含量、比叶面积和比根长,具备较强的水分和养分竞争能力;而乡土植物群落具有较小的比叶面积、较大的叶组织密度和根组织密度,虽然可以较好地维持体内水分和养分的平衡,形成对环境的有效适应,但相比刺槐水分和养分竞争能力较弱。因此,在黄土丘陵区植物恢复重建中,要充分考虑物种对环境的适应性及其对生长的影响,要维持物种适应环境与健康生长间的平衡。
刺槐林; 乡土植物群落; 植物功能性状; 黄土丘陵区; 适应策略
植物功能性状(plant functional traits)是描述植物形态学、生理生态学、生物化学等属性的特性[1],是连接植物与环境的纽带,与生态系统功能密切相关[2-3],是表征生态系统功能变化的重要参数。基于植物功能性状的生态学研究近年来已成为生态学研究的前沿,并在植物功能性状及其与生态环境之间的关系研究取得了较大进展。在植物性状中,叶性状与植物对资源的获取、利用关系密切[4],如比叶面积的变化可反映叶片捕获光的能力和在不同强光下的自身保护能力[5],叶氮含量则直接影响植物的光合作用[6],而根系性状则影响植物吸收水分和养分的能力,如比根长可以表征植物对地下资源的竞争[7-8],根系生物量可反映植物对地下物质和能量的投资情况[9-10],这些信息对理解植物对土壤资源利用的效率具有重要的意义[11]。因此,植物叶片和细根性状在植被—环境关系研究中广泛采用,是分析不同物种生理生长以及对复杂环境响应适应策略的重要媒介。
近年来,越来越多的研究利用植物功能性状来分析外来物种与乡土物种的关系及其影响。研究表明,与乡土物种相比,能够在新生境中获得成功的外来物种往往具有较高的光合能力、资源利用效率、表型可塑性及较低的繁殖成本[12],如在贫瘠、干旱环境下的外来物种多具有具较强的水分和养分竞争能力[13]。通过与本地植物竞争水分、养分、光照及生存的空间等资源条件,外来物种会影响到生态系统的功能和结构,引起乡土物种数量的减少甚至灭绝。刺槐由于其生长速度快,耐干旱贫瘠,更新萌芽能力强,成为黄土高原营造水土保持林广泛采用的树种之一[14-15]。但是从20世纪90年代以来,黄土高原部分地区刺槐林出现了大面积的“小老树”现象,生长缓慢,林下植被更新缓慢,刺槐树梢枯死等[16]。而且林地土壤出现了严重的退化,深层土壤水分也发生了严重亏缺,严重影响了刺槐林的生长[17]。我们认为,“小老树”的出现,一方面可能与刺槐高效的土壤水分养分吸收利用机制有关,也与黄土高原独特的土壤条件有关。刺槐可能具有比乡土植物更具竞争能力的性状特征,可利用土壤中的前期储水迅速生长,并形成对乡土物种的竞争优势,但当前期储水过度消耗后,有限的降雨补充使得刺槐的生长变缓甚至停滞,从而形成“小老树”。但现有研究尚无法验证这一假设。虽然宋光[18]、李俊辉[19]等对不同立地条件下刺槐性状进行了研究,龚时慧[20]、杨士梭[21]等也对黄土丘陵区乡土自然植物群落的功能性状进行了研究,但少有研究对刺槐与乡土植物的功能性状进行比较。刺槐与乡土植物功能性状上是否有差异?差异是否显著?这些差异会对刺槐生长及土壤水分养分有何影响等问题仍需深入研究和探讨。为此,本研究试图通过对陕北黄土丘陵区森林草原过渡带刺槐林和乡土植物群落植物功能性状的调查和测定,比较二者的功能性状在不同地形条件下的变化特征,探讨相同生存环境下刺槐林与乡土植物群落功能性状的差异性,并分析其对土壤水分养分环境的影响,以验证假设。
1 材料和方法
1.1研究区概况
研究区位于陕西省安塞县境内北部,位于延河流域中上游(108°45′—110°28′E,36°23′—37°17′N),地貌类型属于典型黄土丘陵,地形条件复杂,水土流失严重,流域平均海拔1 371.9 m,平均坡度4.3‰,沟壑密度为4.7万条/km2。气候属大陆性半干旱季风气候带,年均气温8.8℃,年均降雨量500 mm,年日照时数2 395.6 h,日照百分率54%,全年无霜期157 d。研究区在植被区划上属典型草原区,自然分布的主要植物种有铁杆蒿(Artemisiasacrorum)、百里香(Thymusmongolicus)、长芒草(Stipabungeana)、白羊草(Bothriochloaischaemum)等,而刺槐(Robiniapseudoacacia)和柠条(Caraganaintermedia)是人工种植最广泛的两个物种[15]。流域内土壤类型主要有黄绵土、黏土、黑垆土等。
1.2样点布设和样品采集
样品采集在2014年8月进行。在研究区内选取林相整齐、林木分布均匀的人工刺槐林地为研究对象,布设2个完整的断面,每个断面包括阴坡和阳坡两个坡向,并按上、中、下坡位设置采样点进行采样,共设12个采样点,每个采样点设置2个10 m×10 m样方。同时在距离刺槐林采样区临近位置选择人为活动干扰较少、发育良好、无人工林覆盖的自然草本群落作为参照,按同样方法布设2个完整的断面,共12个采样点。每个样点设置1 m×1 m样方≥2个(详见表1)。对所有样方,进行群落和物种多度、盖度、群落及物种性状调查,同时记录每个样点的坡向和坡位,用手持GPS实际测量经纬度、海拔。
表1刺槐林和乡土植物群落样地信息表
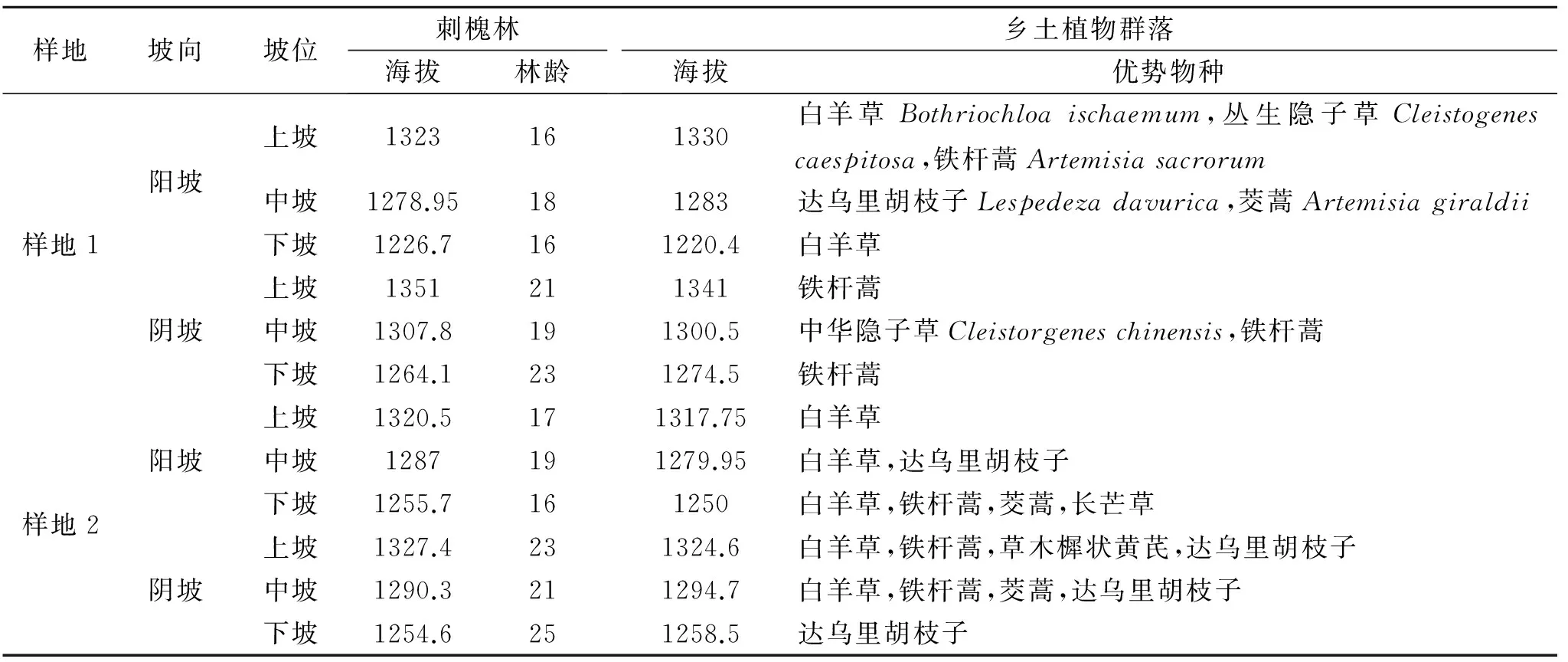
样地坡向坡位刺槐林海拔林龄乡土植物群落海拔优势物种阳坡上坡1323161330白羊草Bothriochloaischaemum,丛生隐子草Cleistogenescaespitosa,铁杆蒿Artemisiasacrorum中坡1278.95181283达乌里胡枝子Lespedezadavurica,茭蒿Artemisiagiraldii样地1下坡1226.7161220.4白羊草上坡1351211341铁杆蒿阴坡中坡1307.8191300.5中华隐子草Cleistorgeneschinensis,铁杆蒿下坡1264.1231274.5铁杆蒿上坡1320.5171317.75白羊草阳坡中坡1287191279.95白羊草,达乌里胡枝子下坡1255.7161250白羊草,铁杆蒿,茭蒿,长芒草样地2上坡1327.4231324.6白羊草,铁杆蒿,草木樨状黄芪,达乌里胡枝子阴坡中坡1290.3211294.7白羊草,铁杆蒿,茭蒿,达乌里胡枝子下坡1254.6251258.5达乌里胡枝子
1.3土壤因子和植物功能性状测定
1.3.1土壤性状的测定土壤的采集是在样方内采用“S”形路线选择5个点进行多点混合土样,用土钻分别采集0—5 cm,5—20 cm,20—40 cm三层土样,分层将5个点的土样混匀后用塑封袋封好带回实验室,经过自然风干后过0.149 mm土筛待用。土壤有机碳采用重铬酸钾—外加热法测定;土壤全氮的测定采用半微量凯式法消化,再用全自动凯氏定氮仪进行测定;土壤磷采用HClO4—H2SO4消煮,用钼锑抗比色法测定全磷,在测定土壤碳、氮、磷含量的同时测定土壤含水率。
1.3.2植物样品的采集刺槐林叶片和根系样品采集:在每个样方的左边、中部和右边各挑选3棵刺槐树,进行样品的采集。在树冠层中部沿东西南北四个方向采集完全展开、没有病虫害且未被遮光的叶片各20片,用于测定叶面积、叶组织密度,再在这些植株上摘取一些叶片混匀装入塑封袋内,带回实验室烘干粉碎封存待测叶片碳、氮、磷用;同时在这3棵刺槐树根部用铁锹挖取土壤剖面,采集一定数量的细根(直径<2 mm),用来测定比根长和根组织密度,将其他根系及附着土壤一起装进塑封袋带回室内清洗干净后烘干粉碎封存待测根碳、氮、磷用。
自然植物群落叶片和根系样品采集:在每个样方内左边、中部、右边各取2~3株长势较好的植株,每个植株上沿东西南北四个方向采取20~30片光照良好、完全伸展的健康叶片,用来测定叶面积、叶组织密度。另在这些植株上摘取一些叶片混匀装入塑封袋内,带回实验室烘干粉碎封存待测叶片碳、氮、磷用;采集过叶片的植株在根部用铁锹挖取土壤剖面,采集一定数量的细根,分成3组,用来测定比根长和根组织密度,将另外的根系装入塑封袋,带回实验室清洗后烘干粉碎封存待测根碳、氮、磷用。
1.3.3植物性状的测定比叶面积(specific leaf area, SLA)(mm2/g)=叶面积/叶干质量,叶组织密度(leaf tissue density,LTD)(mg/mm3)=叶干重(叶片烘干后质量)/叶体积,比根长(specific root length,SRL)(mm/g)=根长/根干质量,根组织密度(root tissue density,RTD)(g/ml)=根干质量/根体积[6]。
植物叶片、根系中有机碳采用重铬酸钾—外加热法测定;全氮采用H2SO4—H2O2消煮法,再用全自动凯氏定氮仪进行测定;全磷的采用H2SO4—H2O2消煮法,钒钼黄比色法测定。
1.4数据处理
1.4.1植物群落性状值计算本研究中的乡土植物是以群落(样点)为单元,用重要值加权平均计算群落性状值,利用采集到的样方内的单个物种的数据,分别计算出各个群落中每个物种的相对多度、相对盖度和相对生物量,再分别将一个群落内i物种的重要值用公式(1)算出,最后将该群落中i物种的平均j性状值用(3)算出。计算公式如下:
IVi=(相对多度+相对盖度+相对生物量)/3
(1)
(2)
(3)
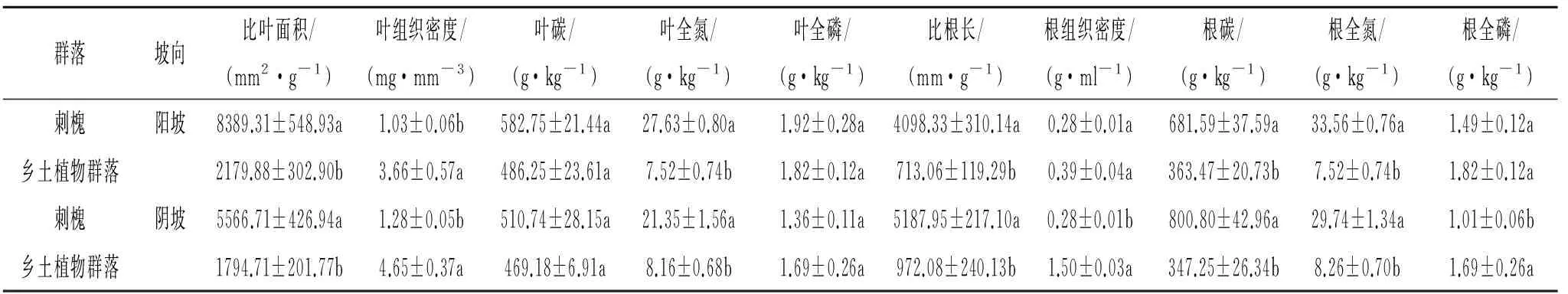
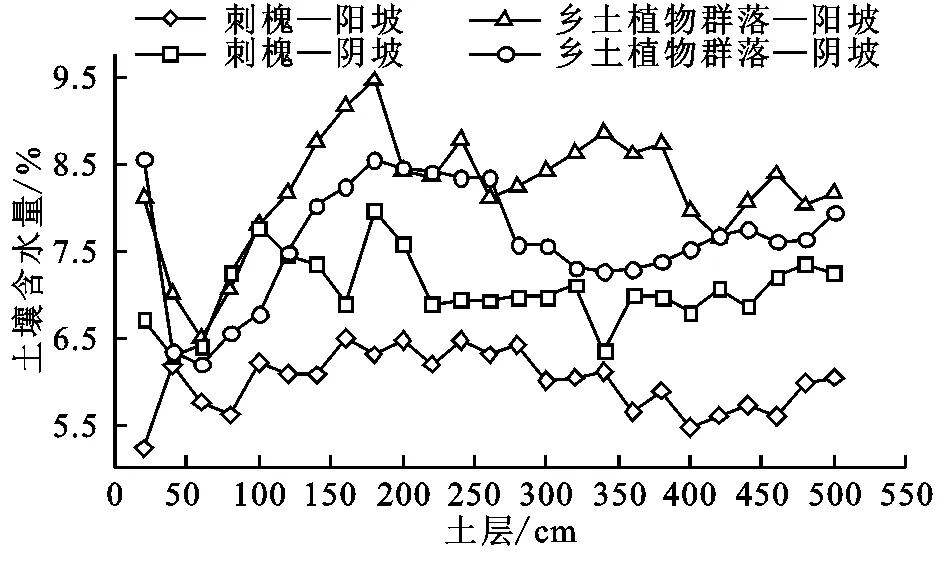
式中:IVi——i物种在该群落中的重要值(0 1.4.2数据处理采用Microsoft Excel 2007进行试验数据预整理;通过对数转换使数据符合正态分布,运用SPSS 16.0对刺槐林和乡土植物群落的功能性状及土壤因子在坡向和坡位分别做单因素方差分析,并用Duncan法对不同坡位各参数平均值进行显著性检验;应用SPSS 16.0对刺槐林和乡土植物群落的功能性状与对应的土壤因子之间进行Pearson相关检验;用Origin 9.0完成图形制作。 2.1刺槐和乡土植物群落功能性状在不同坡向上的变化比较 与乡土植物群落相比,刺槐林的叶C含量、叶N含量、叶P含量、根C含量、根N含量、比叶面积、比根长总体上高于乡土植物群落,根P含量、叶组织密度、根组织密度则较乡土植物群落小(表2)。在阳坡,刺槐叶N含量、根C含量、根N含量、比叶面积及比根长显著高于乡土植物群落(p<0.05),而叶组织密度则显著低于乡土植物群落(p<0.05);在阴坡,刺槐叶N含量、根C含量、根N含量、比叶面积、比根长显著高于乡土植物群落(p<0.05),叶组织密度、根组织密度均显著低于乡土植物群落(p<0.05)。 表2 不同坡向刺槐林与乡土植物群落功能性状比较 注:同列不同小写字母表示不同物种功能性状在同一坡向具有显著性差异(p<0.05)。 2.2刺槐与乡土植物群落功能性状在坡位上的变化比较 在不同坡位上(上、中、下坡位),刺槐叶N含量、根C含量、根N含量、比叶面积、比根长显著高于乡土植物群落(p<0.05),刺槐叶C含量在上坡位和下坡位显著高于乡土植物群落(p<0.05),而刺槐林的叶组织密度在3个坡位上均显著低于乡土植物群落(p<0.05),叶P含量、根P含量均在中坡位显著低于乡土植物群落(p<0.05)(表3)。 2.3刺槐与乡土植物群落土壤因子随地形的变化比较 刺槐林的土壤C含量在阴阳坡上均显著低于乡土植物群落(p<0.05),土壤N含量在阳坡上显著高于乡土植物群落(p<0.05),土壤P含量低于乡土植物群落,且仅在阴坡上差异显著(p<0.05)。在3个坡位上,乡土植物群落的土壤C含量显著高于刺槐林(p<0.05),土壤P含量高于刺槐林但差异不显著(p>0.05),土壤N含量在上坡位和中坡位上高于刺槐林,在下坡位上则低于刺槐林下土壤(图1)。 2.4刺槐林与乡土植物群落土壤水分变化比较 土壤水分是植物生长的主要限制因子之一,为了研究黄土丘陵区刺槐林和自然植物群落在生长过程中对土壤水分的利用情况,本研究对刺槐林下和乡土植物群落在不同坡向和坡位的土壤水分做了对比分析(图2和图3)。 表3 不同坡位刺槐林与乡土植物群落功能性状比较 注:同列不同小写字母表示不同物种功能性状在同一坡位具有显著性差异(p<0.05)。 注:不同小写字母表示不同物种林下土壤在同一坡向(坡位)具有显著性差异(p<0.05)。 图1不同立地条件下刺槐与乡土植物群落的土壤因子比较 图2中,阴阳坡,刺槐林和乡土植物群落5 m土壤含水量变化基本一致。乡土植物群落土壤含水量阴坡高于阳坡,刺槐林也有相同的规律,且乡土植物群落的土壤含水量在阴阳坡均高于刺槐林下的土壤含水量。二者的耗水层主要集中在0—60 cm土层,60 cm之后乡土植物群落土壤含水量呈现明显的上升趋势,180 cm之后逐渐降低,最后趋于稳定状态;刺槐林土壤含水量在阴坡整体上呈现稳定的状态,在阳坡呈现下降的趋势。 图3中,三个坡位上,刺槐林和乡土植物群落5 m土壤含水量变化基本一致。乡土植物群落的土壤含水量表现为下坡>中坡>上坡,刺槐林有相同的规律,且乡土植物群落的土壤含水量均高于刺槐林下的土壤含水量。刺槐林和乡土植物群落土壤含水量整体呈现先上升再下降最后趋于稳定的状态。 图2 刺槐林和乡土植物群落土壤含水量随坡向的分布特征 图3 刺槐林和乡土植物群落土壤含水量随坡位的分布特征 本研究表明,刺槐作为外来物种,在功能性状方面(除叶C,P含量和根C含量外)与乡土植物群落有显著性差异。刺槐林的叶C,N,P含量、根C,N含量、比叶面积、比根长等较乡土植物群落高,根P含量、叶组织密度、根组织密度则表现为刺槐林高于乡土植物群落。刺槐作为豆科植物,具有较高的固氮能力[21],较高的叶片和根N含量有助于刺槐维持较高的光合能力与养分平衡能力[18]。宝乐[22]、王力[23]等的研究表明,比叶面积大的植物通常有较高的生长速率,可以较好地适应资源环境丰富的环境,而比叶面积较小的乡土植物群落对贫瘠的环境则具有更强的适应性。植物叶、根组织密度则可反映植物的防御能力和耐寒性[23]。本研究中乡土植物群落和刺槐林在根组织密度上无显著差异,但乡土植物群落具有较高的叶组织密度,表明乡土植物具有更好的耐旱力和较强的防御力[24]。植物的比根长表征植物根系收益和花费的关系,决定根系吸收水分和养分的能力[15],本研究中刺槐林的比根长显著高于乡土植物群落,表明刺槐相对乡土植物群落,具有更强的水分和养分竞争能力。 土壤是植被正常生长的基础,而植被又是土壤形成过程中的重要因子。例如,植被类型会影响土壤养分的累积、循环或分布[25],而土壤养分又反作用于植物的生长,特别是土壤中的C,N,P三种营养元素,对植物的正常生长影响作用显著[26]。本研究表明,乡土植物群落的土壤C,N,P含量在总体上高于刺槐林。乡土植物群落的土壤养分含量高于刺槐林,这可能与刺槐具有较强的水分和养分竞争能力有关,也可能与物种多样性有关。在本研究中,刺槐林的林下植被更新缓慢,多样性差,植被覆盖度较小,乡土植物群落的多样性较人工林丰富[22],加之刺槐有较强的水分和养分竞争能力,对土壤水分养分的消耗大于乡土植物,因此,乡土植物群落的土壤C,N,P含量在总体上高于刺槐林。在不同坡位上,土壤C,P含量均表现为乡土植物群落高于刺槐林,土壤N含量在下坡位表现为刺槐林高于植物群落,造成这种现象的原因可能是因为下坡位土壤水分条件较好,且可沉积中上坡的侵蚀土壤,土壤养分含量较高,可满足刺槐的正常生长[27],从而可通过较高的生物量形成和枯枝落叶回归土壤,提高土壤的N含量[28]。这表明,要使刺槐发挥较好的保持水土和改善土壤生态环境的作用,必须要注意营造刺槐林的地形条件,尽量避免在坡面的中上部营造,最好选择在沟坡下部。而在中上部养分较贫瘠的立地环境,乡土植物群落较刺槐有更好的土壤改善作用[29]。 本研究表明,黄土丘陵区刺槐林的需水量明显高于乡土植物群落。乡土植物群落的土壤含水量在阴坡和阳坡均高于刺槐林,而且刺槐林和乡土植物群落的土壤含水量均表现为阴坡高于阳坡。从下坡到上坡,乡土植物群落和刺槐林的土壤含水量表现为逐渐减少的趋势,且乡土植物群落的土壤含水量均高于刺槐林下的土壤含水量。阴坡和阳坡由于光照条件、水分蒸发情况、植被覆盖度等差异[30],阴坡土壤含水量高于阳坡。降雨后,土壤水分受降水再分配的影响,在重力的作用下,向下运移,从而造成了刺槐林和自然植物群落不同坡位土壤水分含量的差异[31]。Wright等[32]认为,高的叶养分浓度是生长在干旱地区的植物保持水分的选择性策略。在本研究中,刺槐林叶片C,N,P的浓度、比叶面积较乡土植物群落高,而刺槐林下土壤含水量较乡土植物群落低,说明刺槐林在干旱的环境中,通过提高叶片养分浓度,吸收土壤中水分来保持体内的含水量,维持自身的生长,而非降低比叶面积的策略。加之刺槐林根系具有比根长,具有较强的水分养分吸收能力,因此,刺槐林下土壤含水量较低,这与韦兰英等[33]的研究结果相一致。 综上所述,在资源贫瘠的环境下(中上坡位),刺槐林一方面会分配更多的养分给吸收和保持水分的器官或者组织来构建自身的防御系统,减少对刺槐林生长的物质投入,导致刺槐的生长速率降低。因此,“枯稍”“生长矮小”等刺槐不良表现,可能就是上述原因共同作用的结果。在黄土丘陵区植被恢复重建中,要充分考虑物种对环境的适应性及其对生长的影响(主要是养分和水分的消耗特点),要维持物种适应环境与健康生长间的平衡。当物种为适应环境而严重降低植物生长时,在这些生境中的植被恢复应剔除这些物种以促进黄土丘陵区植被恢复重建得以健康持续地发展。 [2]孟婷婷,倪健,王国宏.植物功能形状与环境和生态系统功能[J].植物生态学报,2007,31(1):150-165. [3]Lienin P, Kleyer M. Plant trait responses to the environment and effects on ecosystem properties[J]. Basic and Applied Ecology,2012,13(4):301-311. [4]Vendramini F, Diaz S, Gurvich D E, et al. leaf traits as indicators of resource-use strategy in floras with succulent species[J]. New Phytologist,2002,154(1):147-157. [5]张林,罗天祥.植物叶寿命及其相关叶性状的生态学研究进展[J].植物生态学报,2004,28(6):844-852. [6]Pérez-Harguindeguy N, Díaz S, Garnier E, et al. New handbook for standardised measurement of plant functional traits worldwide[J]. Australian Journal of Botany,2013,61(3):167-234. [7]Koike T, Kitao M, Quoreshi A M, Matasuura Y. Growth characteristics of root-shoot relations of three birch seedings raised under different water regimes[J]. Plant and Soil,2003,255(1):303-310. [8]Place G, Bowman D, Burton M, et al. Root penertration through a high bulk density soil layer:differential response of crop and species[J]. Plant and Soil,2008,307(1/2):179-190. [9]Markesteijn L, Poorter L. Seeding root morphology and biomass allocation of 62 tropical tree species in relation to drought-and shade-tolerance[J]. Journal of Ecology,2009,97(2):311-325. [10]Valdés M, Asbjornsen H, Gómez-Cárdenas M, et al. Drought effects on fine-root and ectomycorrhizal-root biomass in managedPinusoaxacanaMirov stands in Oaxaca, Mexico[J]. Mycorrhiza,2006,16(2):117-124. [11]Jose S, Williams R, Zamora D. Belowground ecological interactions in mixed-species forest plantations[J]. Forest Ecology and Management,2006,233(2/3):231-239. [12]Oliva M A, Kuki K N, Mielke M S, et al. Key leaf traits indicative of photosynthetic plasticity in tropical tree species[J]. Trees,2015,29(1):247-258. [13]Cornelissen J H C, Lavorel S, Garnier E, et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J]. Australian Journal of Botany,2003,51(4):335-380. [14]Wright I J, Reich P B, Westoby M. Strategy shifts in leaf physiology, structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats[J]. Functional Ecology,2001,15(4):423-434. [15]郑颖,温仲明,宋光,等.环境及遗传背景对延河流域植物叶片和细根功能性状变异的影响[J].生态学报,2014,34(10):2682-2692. [16]Craine J M, Lee W G. Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand[J]. Oecologia,2003,134(4):471-478. [17]徐冰,程雨曦,甘慧洁,等.内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联[J].植物生态学报,2010,34(1):29-38. [18]宋光,温仲明,郑颖,等.陕北黄土高原刺槐植物功能性状与气象因子的关系[J].水土保持研究,2013,20(3):125-130. [19]李俊辉,李秧秧.立地条件和树龄对刺槐叶形态及生理特性的影响[J].水土保持研究,2012,19(4):176-181. [20]龚时慧,温仲明,施宇.延河流域植物群落功能性状对环境梯度的响应[J].生态学报,2011,31(20):6088-6097. [21]杨士梭,温仲明,苗连朋,等.黄土丘陵区植物功能性状对微地形变化的响应[J].应用生态学报,2014,25(12):3413-3419. [22]宝乐,刘艳红.东灵山地区不同森林群落叶功能性状比较[J].生态学报,2009,29(7):3692-3703. [23]王力,邵明安,李裕元.陕北黄土高原人工刺槐林生长与土壤干化的关系研究[J].林业科学,2004,40(1):84-91. [24]施宇,温仲明,龚时慧.黄土丘陵区植物叶片与细根功能性状关系及其变化[J].生态学报,2011,31(22):6805-6814. [25]McQuilkin W E. The natural establishment of pine in abandoned fields in the Piedmont Plateau region[J]. Ecology,1940,21(2):135-147. [26]侯庆春,黄旭,韩仕峰,等.黄土高原地区小老树成因及其改造途径的研究[J].水土保持学报,1991,5(1):64-72. [27]马玉珠,钟全林,靳冰洁,等.中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子[J].植物生态学报,2015,39(2):159-166. [28]Wright I J, Reich P B, Comelissen J H C. Assessing the generality of global leaf trait relationship[J]. New Phytologist,2005,166(2):485-496. [29]成向荣,赵忠,郭满才,等.刺槐人工林细根垂直分布模型的研究[J].林业科学,2006,42(6):40-48. [30]王春红,王治国,铁梅,等.河沟流域土壤水分空间变化及制备分布于生物量研究[J].中国水土保持科学,2004,2(2):18-23. [31]潘占兵,余峰,王占军,等.宁南黄土丘陵区坡向、坡位对苜蓿地土壤含水量时空变异的影响[J].水土保持研究,2010,17(2):141-144. [32]Wright I J, Westoby M. Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species[J]. Functional Ecology,2003,17(1):10-19. [33]韦兰英,上官周平.黄土高原白羊草、沙棘和辽东栎细根比根长特性[J].生态学报,2006,26(12):4164-4170. Comparative Studies on Plant Functional Traits of Leaf and Fine Root BetweenRobiniapseudoacaciaand Local Plant Community in Hilly-Gully Region of the Loess Plateau, China WANG Hongxia1, WEN Zhongming1,2,3, GAO Guoxiong1,2, QI Dehui2, GUO Ru3, YANG Shisuo1 (1.InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling,Shaanxi712100,China; 2.CollegeofNaturalResourcesandEnvironment,NorthwestA&FUniversity,Yangling,Shaanxi712100,China; 3.ResearchCenterofSoilandWaterConservationandEcologicalEnvironment,ChineseAcademyofSciences,Yangling,Shaanxi712100,China) Plant functional trait is an important medium to express the relationships between plants and environment. The difference in functional traits among different species decides their competitiveness in a community, affecting the structure, function and development of the community. We selected artificial forest ofRobiniapseudoacaciaand local plant community in forest-steppe zone in hilly-gully region in Loess Plateau, and compared the differences of the functional traits betweenRobiniapseudoacaciaand local plant community under different site conditions and clarified their adaptive strategies to local environment. The results showed that: (1) within the same slope aspect, there was a significant difference in the nitrogen content of leaf, carbon and nitrogen contents of root, specific leaf area, leaf tissue density and specific root length betweenRobiniapseudoacaciaand local plant community (p<0.05) while the phosphorous content of root and the root tissue density showed significant difference only in the north-facing slope (p<0.05), the function of man-made forest ofRobiniapseudoacaciacharacter values was higher than the local plant community (except leaf tissue density, root tissue density and phosphorus content of root, under the same slope position, the carbon and nitrogen contents, specific leaf area and specific root length ofRobiniapseudoacaciawere significantly higher than those of the local plant community; (2) the soil carbon and nitrogen contents ofRobiniapseudoacaciaplantation were lower than those of the local plant community in south- and north-facing slopes, and the soil phosphorus content was higher than that of the local plant communities in the south-facing slope, and opposite in north-facing slope, the soil carbon, nitrogen, phosphorus contents ofRobiniapseudoacaciaplantation and local plant community between slope positions were not significantly different(p>0.05), under different slope positions, only soil carbon content ofRobiniapseudoacaciaplantation and local plant community showed significant difference (p<0.05); (3) soil moisture content ofRobiniapseudoacaciaplantation is lower than local plant community. In the arid environment,Robiniapseudoacaciahad the higher leaf and root nutrients, and higher SLA and SRL, showing a higher competitive ability for soil water and nutrient use. Local plant community had smaller specific leaf area, larger leaf and root tissue density to maintain the balance of water and nutrients, forming adaptive strategies to local environment. However, local plant community had the lower competitiveness for water and nutrients compared withRobiniapseudoacacia. Therefore, we should fully consider the adaptability of species to the environment and effect of the adaptability on species growth, the balance between the species adaptability to environment and their health growth should be maintained in vegetation restoration in the loess hilly-gully region. plant functional traits;Robiniapseudoacacia; local plant community; hilly Loess Plateau; adaptive strategies 2015-09-17 2015-09-30 国家自然科学资助项目(41271297);中国科学院重要方向项目(KZCX2-EW-406) 王红霞(1988—),女,山西省临汾人,硕士研究生,主要从事植物功能生态学方面的研究。E-mail:1096352712@qq.com 高国雄(1969—),男,陕西省清涧县人,博士,副教授,主要从事水土保持工程方面的研究。E-mail:ggx@nwsuaf.ed.cn 温仲明(1969—),男,陕西省定边人,博士,副研究员,主要从事植物生态恢复方面的研究。E-mail:zmwen@ms.iswc.ac.cn Q948 A 1005-3409(2016)01-0001-072 结果与分析



3 结论与讨论
猜你喜欢
现代园艺(2020年2期)2020-12-22
小溪流(故事作文)(2018年10期)2018-12-28
森林工程(2018年4期)2018-08-04
时代农机(2018年11期)2018-03-17
现代园艺(2018年2期)2018-03-15
绿色科技(2017年3期)2017-03-14
绿色科技(2017年1期)2017-03-01
湖南林业科技(2017年1期)2017-02-06
故事作文·低年级(2016年10期)2016-09-10
电源技术(2016年9期)2016-02-27
