
无线电追踪技术在地下啮齿动物研究中的应用
2016-10-20 09:50姬程鹏花立民杨思维周延山
草业科学 2016年9期
姬程鹏,花立民,杨思维,周延山,楚 彬
(甘肃农业大学草业学院,甘肃 兰州 730070)
无线电追踪技术在地下啮齿动物研究中的应用
姬程鹏,花立民,杨思维,周延山,楚 彬
(甘肃农业大学草业学院,甘肃 兰州 730070)
营地下生活的啮齿动物被誉为陆地生态系统的工程师,但其长期栖居于地下的生活习性难以被人们直接观察,给动物行为学和生态生物学等研究带来了挑战。随着科学技术的发展,无线电技术的小型化和轻量化为地下啮齿动物研究提供了可行性。国外学者已成功将无线电追踪技术应用于对地下啮齿动物洞道系统、活动节律、巢域面积变化以及社群制度等的研究。但国内将其应用于地下啮齿动物研究的较少。本文根据目前国内外研究现状,对无线追踪技术的研究内容和研究方法进行综述,以期为我国地下啮齿类研究提供参考。
无线电追踪;地下啮齿动物;行为学;生态学
地下啮齿动物(subterraneanrodents)是一类营地下生活的植食性小型哺乳动物[1-2]。在全世界,此类动物主要包括囊鼠科(Geomyidae)、八齿鼠科(Octodontidae)、栉鼠科(Ctenomyidae)、滨鼠科(Bathyergidae)等物种,广泛分布于非洲、美洲、欧亚大陆的热带草原、草原、干旱及半干旱灌丛[2-4]。在我国分布的主要以竹鼠亚科(Rhizomyinae)和鼢鼠亚科(Myospalacinae)为主[5]。地下啮齿动物活动包括挖掘、采食、贮食、交配、繁殖等,主要在其建造的地下洞道系统内完成[6]。在生态系统中,地下啮齿动物的挖掘活动一方面会导致与之相关的土壤类型、发育速率、营养可利用性以及微地形等非生命环境的变化[7]。另一方面,地下啮齿动物采食植物根茎,推土造丘影响植物地上和地下生物量,对植被覆盖、群落结构和植物多样性等方面有着重要影响[8-9]。因此,许多学者认为地下啮齿动物是陆地生态系统的“工程师”[10]。研究地下啮齿动物的生活习性和行为特征对保护生物多样性和科学控制其危害有重要意义。但是,由于地下啮齿动物几乎全在地下活动,难以观察,给研究者带来了很大的挑战。
无线电追踪技术(radiotracking)是一门新兴技术,是利用无线电波的发射和接收来确定动物的位置并进行追踪的动物学研究方法[11]。由于不需要与目标动物直接接触,无线电追踪研究可以更准确地提供动物在自然状态下的活动状况。对地下啮齿动物而言,由于其取食、交配、育幼等大多数活动都发生在自己建造的洞道系统里[12],无线电追踪技术可以更好地研究自然状态下地下啮齿动物的活动规律,尤其是空间利用特征和活动节律等方面[13],因而被广大研究者所青睐。加之无线电追踪技术研究动物更符合国际动物福利法规,因此,越来越多的国外学者将无线电追踪技术应用到地下啮齿动物研究中。但国内将无线电追踪技术用于地下啮齿动物的研究报道很少,从20世纪80年代至今,仅有周文扬和窦丰满[14]曾使用无线电追踪技术研究高原鼢鼠的活动节律。本文对近25年国内外利用无线电追踪技术对地下啮齿动物的相关研究与应用进行归类总结,以期为国内学者采用此项技术研究地下啮齿动物提供技术借鉴。
1 研究对象
全世界地下啮齿动物有4科4亚科,约300种[15]。无线电追踪技术最早在1958年被应用到半地下啮齿动物土拨鼠(Marmota monax)的研究[16]。根据1990-2014年的文献记载,目前应用无线追踪技术开展研究的地下啮齿动物有4科14种,约占到地下啮齿动物总种数的5%。从研究频率的高低来看,依次为栉鼠科、滨鼠科、八齿鼠科、鼹形鼠亚科、囊鼠科等(表1)。从研究对象的分布来看,主要集中在非洲和南美洲两地,研究对象多为当地的特有品种,如滨鼠科的研究主要集中在非洲东部[10,17];栉鼠科和八齿鼠科的研究集中在南美洲的南部[11,18-19]。从研究对象的科学价值来看,无线追踪技术首次用于濒危的平原囊鼠奥索卡亚种(Geomys bursarius ozarkensis),因为平原囊鼠的亚种被认为是“最应该得到保护的物种”[20]。Connior和Risch[20]通过研究它们的空间利用和生活史特点,为保护该物种提供了理论依据。
2 研究内容
2.1无线追踪技术在动物巢域研究中的应用
Burt[21]把地下啮齿动物的巢域定义为在主巢附近进行采食、交配和照顾幼仔的活动场所。巢域是地下啮齿动物进行各种生命活动的场所,它与繁殖、栖息地质量以及季节变化等有着密切关系[22],也是野生动物资源保护和鼠害防治的热点(图1)。根据无线电追踪数据可以计算出不同时段地下啮齿类两性之间活动巢域面积的大小,为巢域尺度的栖息地质量做出评估[23]。无线电追踪技术用于巢域研究通常是在地上建立坐标系或网格线,利用手持Yagi天线,通过无线电连续追踪佩戴有无线发射器的目标啮齿动物活动,记录其活动的位点,然后利用软件Calhome[24]、Ranges[18]和ArcView[25]等做出地下啮齿动物的活动区域。也可将Yagi天线固定在宽频的接收机上来确定地下啮齿动物的活动位点,如Cutrera等[22]标记了同域分布的10只蓝栉鼠和7只南方栉鼠,发现南方栉鼠的体重是蓝栉鼠的3倍,但利用无线追踪技术结合软件Calhome中的Theadaptivekernelmethod发现巢域面积却是蓝栉鼠的19倍,符合大个体生物往往需要更大的面积来采集食物以满足高的能量需求的特点[26],同时研究发现,蓝栉鼠巢域上的土壤紧实度是南方栉鼠的2.06倍,得出种间由于对土壤选择不一致导致在小时空尺度上的区域隔离,这个研究结果可能是因为个体之间洞道的结构不一样导致[27]。Cutrera等[13]利用无线追踪技术研究蓝栉鼠不同性别巢区,结合软件Calhome中100%minimumconvexpolygon(MCP)发现,蓝栉鼠为一夫多妻制,而且雄性巢域显著大于雌性巢域。这与周文扬和窦丰满[14]研究的高原鼢鼠巢区结论相似。并且Cutrera等[13]研究发现,不同性别巢域大小是由交配制度引起的,这与Fisher和Owens[28]对沙袋鼠(Macropod marsupials)的研究相似,同时还发现雄性巢域的空间分布是由它附近可以交配或者防御的伙伴所处的位置和数量决定的,与Komer和Brotherton[29]对哺乳动物一夫一妻制的研究结果类似。研究者们利用无线追踪技术,结合软件Ranges中的95%MCP做出了地下啮齿动物的巢域面积季节变化[17,30-32]。除Calhome和Ranges软件,利用ArcView软件研究巢域大小,可以同时做出每个个体90%和50%的活动区域,其中90%的区域被作为地下啮齿类的整个巢域范围,并且移除了10%的错误位点和偶然发生的袭击行为出现的位点[33],50%的区域被作为地下啮齿类最常用的核心区域[34]。地下啮齿动物巢域上方与非巢域上方的植被群落结构、地上生物量和地下根量、土壤的理化性状(紧实度、容重、含水量)不同,利用无线电追踪技术可以直观地研究影响地下啮齿动物巢域尺度栖息地选择和巢域面积季节变化的环境因素,为地下啮齿动物栖息地选择研究在巢域尺度提供了新方法和思路。

表1 无线电追踪技术研究的地下啮齿动物种类、地理分布及其发射器型号Table 1 The species, geographic distribution and transmitter model of subterranean rodents studied by radio tracking
注:研究频率指该种被研究的次数,以发表的文章数量统计为准。
Note:Researchfrequencymeansthetimesofthesespecieshavebeenstudiedaccordingtothepublishedpapers.
2.2无线追踪技术在活动节律研究中的应用
地下啮齿动物活动节律一方面是因动物自身内在机制与环境变量(光周期和热量条件)协调的结果[35-36],另一方面也与捕食的压力大小[37]以及外界的刺激特征有关[38]。活动节律研究可以提供地下啮齿类每天的活动规律,为进一步研究其繁殖和扩散等行为学特征提供理论依据,活动节律也一直是研究的热点之一(图1)。利用无线追踪技术研究活动节律,通常是在单位时间内连续监听并记录地下啮齿类所处的位点,同时计算两个连续位点之间的移动距离。为不影响地下啮齿类的正常活动规律,两次监测之间应保证有足够的时间间隔。Ebensperger等[18]通过2h监测一次,在每个活动位点插上标记,同时用手持GPS记录坐标,发现阿根廷胶鼠的活动主要集中在夜间,同时发现它们有相对较好的温度调节机制,可以忍受夜间很低的环境温度,同时避开白天的高温,这与Bozinovic和Contreras[39]对两种荒漠食草啮齿类的研究结果一致。一般在沙漠生境下营社群生活的物种适应这种活动规律[40-41],但在研究时段内发现,阿根廷胶鼠营独居生活,可能与研究的时间长短也有关系。klíba等[42]利用无线追踪技术与地温计测温结合研究发现,安塞尔鼹鼠的活动集中在地表洞道温度最高的时候进行。Cutrera等[13]通过无线电追踪监测发现,蓝栉鼠的活动节律具有多个高峰,Luna等[43]在半自然状态下研究栉鼠也得到了相同的结果,但是雌雄之间的活动节律无明显差异,可能与开展试验的时间有关。Urrejola和Lacey[44]通过每小时监测,发现野外生活的鼹八齿鼠的活动主要集中在白天,但是实验室研究捕获的鼹八齿鼠活动却集中在晚上,说明实验室条件下研究地下啮齿动物的活动节律不一定能反映其自然状态下活动节律。Kas和Edgar[45]、Kenagy等[46]研究实验室状态下和野外自由生活下的灌丛八齿鼠(Octodon degus)的活动节律也证实了这一点。因此,无线电追踪技术在研究自然状态下地下啮齿动物活动节律上有重要意义。
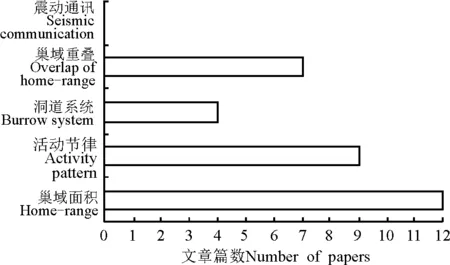
图1 无线电追踪技术在地下啮齿类研究内容的分布Fig.1 Distribution of research contents in the subterraneanrodents studied by radio tracking
2.3无线追踪在洞道系统研究中的应用
地下啮齿动物基本上完全营地下生活,且主要以植物的根系为食物,必须进行持续的挖掘活动才能获得足够的食物。由于这种特殊的习性,常可在土壤中形成庞大而复杂的洞道系统[36,47]。洞道系统是地下啮齿动物进行取食、交配、育幼等活动的主要场所[12]。在食物缺少的时候,地下啮齿动物会通过挖掘洞道来寻求食物资源,但并非每天都利用整个洞道系统。繁殖期的雄性巨型鼹鼠每天利用不足20%的巢域范围[17]。南方栉鼠每天只利用总巢域范围的9%,而蓝栉鼠每天利用的巢域范围只有南方栉鼠的一半[21]。由于地下啮齿动物活动的特殊性,即使利用无线追踪技术,也不能完整立体地绘制出地下啮齿动物的洞道系统。因为利用无线电追踪技术得出来的都是一系列散布的点,而洞道系统是复杂庞大的立体网络结构。因此,利用无线电追踪技术和传统的挖掘技术相结合成为了绘制地下啮齿动物完整洞道系统的有效方法[48]。klíba等[10]利用该方法绘制出了霜鼠完整的洞道系统,周文扬和窦丰满[14]也利用自制无线电追踪和连续挖掘绘制出了我国特有物种高原鼢鼠的洞道系统。
2.4无线追踪技术在巢域重叠研究中的应用
严格意义来讲,地下啮齿动物的生活习性有两类,即营独居生活和营群居生活[49]。巢域重叠为相邻的两只或若干只地下啮齿动物提供了相同的栖息地,重叠的区域可能是同个巢域子代的扩散,也可能是雌雄之间进行交配的场所。利用无线电追踪技术研究不同生活习性下地下啮齿动物巢域重叠也成为研究地下啮齿动物繁殖或者扩散行为的热点之一。巢域重叠的研究方法与巢域的研究方法基本一致,即利用电脑软件做出各自的巢域范围,然后分析相互之间的重叠区域。klíba等[10]研究独居行为的霜鼠发现两个连续的巢域中只有很低的巢域重叠,原因可能是地下啮齿动物的洞道系统有些是长期利用,有些只是临时利用。对栉鼠属不同种的研究发现它们各自巢域彼此相互临近,但均没有重叠区域,从而证明栉鼠属不同种是独居动物[13,22,50-51]。阿根廷胶鼠活动范围很大,但是相互重叠的区域很低,因此,营社群行为的阿根廷胶鼠也被证明是独立生活[17]。在雨季和旱季两个季节研究棘鼠,发现相邻巢域都有大量的重叠区域,巢域面积重叠的增大可能意味着繁殖活动的加剧[34]。因此,巢域重叠也成了研究地下啮齿动物社群结构和繁殖策略的间接方法之一。
2.5无线电追踪技术在震动通讯中的应用
地下啮齿动物常年生活在地下,视觉和听觉功能在洞道系统中受到了很大的限制[52],种内个体之间如何进行信息交流,如何在合适的时间进行交配繁殖活动也成了研究地下啮齿类的关键问题之一。声音在土壤里不能进行长距离的传播,因此很多地下哺乳动物依靠震动传递信号,用来求偶和标记领地[53-54]。利用无线电追踪技术把相邻的地下啮齿动物佩戴发射器,同时把地震波检波器放在两个巢域之间,录下它们发出的震动信号,结合室内试验,分析数据,即可得出代表不同内容的震动信号。Hrouzková等[55]通过两次试验,第1次,在实验室内将非洲鼹鼠(Tachyoryctes daemon)放在人工模拟洞道内录下它们产生的震动信号,第2次,在相邻近的洞道系统之间放上障碍物再次录下震动信号。结果发现两次都产生低频率的震动信号,但是当一只靠近另一只的情况下则产生了高频率的震动信号。李金刚等[54]也通过实验室试验发现甘肃鼢鼠(Myospalax cansus)具有震动通讯现象,雌鼠的敲击频次高于雄鼠的且持续时间更长,而且震动声的能量主要集中在低频区域。这两组试验是在实验室模拟的情况下做的,如果可以在实验室内把各种震动波的信号收集分析完整,在野外结合使用无线电追踪技术把相邻巢域之间佩戴发射器的地下啮齿类在自然状态下发出的不同类型的震动波收录,结合实验室数据就可以得出更能反映自然状态下和更有说服力的结果。如果破译出了不同种类的地下啮齿类的震动信号,就可以在繁殖季节有针对性地采取措施扰乱地下啮齿类之间的震动通讯信号,从而达到低成本、高效率的防治目的,且不会对人畜健康产生威胁。在自然状态下的很多地下啮齿类具有侵占行为[56],即地下啮齿类通过敲击洞壁发出震动波以确定邻居是否有回应,如果没有回应就会很快侵占对方巢域,随着技术的进步,可以把需要研究的地下啮齿类临近巢域的个体活捕并佩戴发射器,待其适应发射器后,把其中一只捕获掉,利用无线电追踪,观察周围的其它同种个体是否有侵占行为。侵占行为也为防治工作提供了新思路,在鼠害防治过程中,单一、小面积的捕杀活动后,相邻个体又会快速入侵,扩大自己的巢域,从而使种内竞争压力变小、环境舒适度提高、种群繁殖速度升高,进而导致种群数量增加。因此,将巢域侵占与种群数量变化结合研究可以为地下啮齿动物防治提供技术借鉴。
3 研究方法
无线电追踪技术研究地下啮齿动物生态生物学特性,首要任务是实现目标动物的无损伤活捕,以保证后期监测个体能够表现出与自然状态下其它个体一样的活动规律。活捕之后一般还需要进行麻醉-佩戴发射器-放回捕获点-在活动区域上方建立网格线-确定巢域等步骤。目前使用无线追踪技术的设备主要有4家公司,分别为英国的Biotrack公司、美国的AVM公司、WildlifeMaterial公司和AdvancedTelemetrySystem公司。我国目前有天津市浙海公司也生产发射器,但多用在鸟类或大型动物上。发射器的重量也有严格的要求,一般不能超过目标动物的4%[10,13],发射器的佩戴方法最常用的有3种:环颈式佩戴、斜跨式和粘贴式佩戴,考虑到动物有换毛特点,一般经常采用前两种佩戴方法[57]。无线追踪所用天线是非常重要的信号接收设备。有Yagi天线、双极(dipole)天线、H(H-adcock)型天线、环形(loop)天线之分。目前,Yagi天线能够很好地分辨真实信号和反射信号,因此被研究者所青睐[58]。手持Yagi天线可以较为精确地确定地下啮齿动物的活动位点。固定的Yagi天线系统配合宽频接收机可以连续地监测地下啮齿动物的活动,但由于距离较大,会影响确定位点的精确性。也有学者直接利用手持GPS来记录地下啮齿动物的活动坐标,这种方法简便易行,但对设备的精确度要求较高。对于地下啮齿类所处位点的判定通常采用三点定位法[59],将得到的散点通过软件计算得出地下啮齿动物的活动时间和活动范围的大小。
4 机遇和挑战
目前,国外将无线电追踪技术应用于大型动物[11,60]和地下啮齿动物研究方面相对比较成熟[17,20,61-62],国内有周文扬和窦丰满[13]在1990年利用自制无线电技术研究高原鼢鼠的活动规律和繁殖交配方式。周建伟等[57]于2013年在高寒草甸测试过3种发射器的佩戴方法及信号问题,但从1990年至今,利用无线电追踪技术在地下啮齿动物上的研究成果鲜见报道。目前啮齿动物对生态、生产的影响极大,如在黑龙江地区地上害鼠黑线姬鼠(Apodemus agrariusPallas)和花鼠(Eutamias sibiricus),地下害鼠中华鼢鼠(Myospalax fontanieri)造成黑龙江红松(Pinus koraiensis)、樟子松(Pinus sylvestrisvar. mongolica)和云杉(Picea asperata)等林业大面积受损[63]。在青藏高原草地生态系统中,高原鼢鼠是两大优势啮齿类之一,高原鼢鼠适度的挖掘活动对生态系统的营养循环、空间异质性和微量气体释放均有重要的有利影响,但当高原鼢鼠的密度超过一定数量后其取食和挖掘活动会加剧草地退化[64],因此研究高原鼢鼠的习性和活动规律对于控制其危害、保护草地生物多样性和实现草地畜牧业可持续发展具有重要意义[65]。目前,无线电发射器的小型化和轻量化为研究高原鼢鼠的生活习性带来了可能,同时利用无线电追踪技术与分子生物学方法的结合,研究高原鼢鼠扩散对象和扩散时间等问题,对草原鼠害防治和维持草地生物多样性都有重要意义[66]。
虽然无线追踪技术有很多优点,但是任何技术都处在不断完善的过程中。由于发射器一般是根据研究对象和研究需要定制。目前国内外学者遇到的问题有以下几点:第一,由于发射器小型化,因此电池有效寿命通常只有3~9个月,不便于长期监测[20]。第二,发射项圈易脱落。由于啮齿类生长速度较快,因此发射器项圈佩戴要松紧合适,如果发生脱落,就必须通过天线定位,并挖掘项圈,费时费力[67]。第三,无线电信号不稳定,一般无线电发射信号在地面接收的最佳范围是0~5m[57]。如果佩戴无线电发射器的地下啮齿动物活动距离过深,信号衰减严重,影响到监测效果[17]。同时无损伤活捕技术也是制约我国学者使用该技术来研究地下啮齿动物的主要原因,国外学者常用的活捕器械有Hickmantrap[68],美国TomahawkLiveTrap公司的产品[69],还有一些自制的活捕器械,如针对囊鼠设计的活捕笼[70],我国学者针对鼢鼠设计的无损伤活捕笼[71]。另外,关于麻醉技术,国际上也没有统一的标准和流程,这些因素都给国内外学者带来了很多挑战。
< class="emphasis_italic">References
:
[1]EllermanJR.Thesubterraneanmammalsoftheworld.TransactionsoftheRoyalSocietyofSouthAfrica,1956,35(1):11-20.
[2]NevoA.Adaptiveconvergenceanddivergenceofsubterraneanmammals.AnnualReviewofEcologyandSystematics,1979,10:269-308.
[3]MielkeHW.Moundbuildingbypocketgophers(Geomyidae):TheirimpactonsoilandvegetationinNorthAmerica.JournalofBiogeography,1977,4(2):171-180.
[4]AndersenDC.Below-groundherbivoryinnaturalcommunities:Areviewemphasizingfossorialanimals.TheQuarterlyReviewofBiology,1987,62(3):261-286.
[5]蒋志刚,马勇,吴毅,王应祥,周开亚,刘少英,冯祚建.中国哺乳动物多样性及地理分布.北京:科学出版社,2015:343-349.
JiangZG,MaY,WuY,WangYX,ZhouKY,LiuSY,FengZJ.China’sMammalDiversityandGeographicDistribution.Beijing:SciencePress,2015:343-349.(inChinese)
[6]张堰铭,刘季科.地下鼠生物学特征及其在生态系统中的作用.兽类学报,2002,22(2):144-154.
ZhangYM,LiuJK.Thebiologicalcharacteristicsofsubterraneanrodentsandtheirrolesinecosystem.ActaTheriologicaSinica,2002,22(2):144-154.(inChinese)
[7]刘锦上.高原鼢鼠的洞道空间对高寒草甸植被性状的影响.兰州:兰州大学硕士学位论文,2009.
LiuJS.EffectsofburrowspaceofPlateauZokoronvegetationcharactersonalpinemeadows.MasterThesis.Lanzhou:LanzhouUniversity,2009.(inChinese)
[8]施大钊.鼠类的危害、利用及其控制.生物学通报,1996,31(1):11-14.
ShiDZ.Theharm,useandcontroloftherodents.BulletinofBiology,1996,31(1):11-14.(inChinese)
[9]韩天虎,张卫国,丁连生.高原鼢鼠栖息地的植被特征.草业学报,1999,8(2):43-49.
HanTH,ZhangWG,DingLS.FeaturesofvegetationinMyospalax baileyihabitat.ActaPrataculturaeSinica,1999,8(2):43-49.(inChinese)
[11]KenwardRE,WallsSS,HodderKH.Lifepathanalysis:Scalingindicatesprimingeffectsofsocialandhabitatfactorsondispersaldistances.JournalofAnimalEcology,2001,70(1):1-13.
[12]NevoE.MosaicEvolutionofSubterraneanMammals:Regression,ProgressionandGlobalConvergence.Oxford:OxfordUniversityPress,1999:88-91.
[13]CutreraAP,AntinuchiCD,MoraMS,VassalloAI.Home-rangeandactivitypatternsoftheSouthAmericansubterraneanrodentCtenomys talarum.JournalofMammalogy,2006,87(6):1183-1191.
[14]周文扬,窦丰满.高原鼢鼠活动和巢区的初步研究.兽类学报,1990,10(1):31-89.
ZhouWY,DouFM.Studiesonactivityandhomerangeofplateauzokor.ActaTheriologicaSinica,1990,10(1):31-89.(inChinese)
[15]WilsonDE,ReederDM.MammalSpeciesoftheWorld:ATaxonomicandGeographicReference.3rdrevisededition.TheUnitedStates:JohnsHopkinsUniversityPress,2005:859-1570.
[16]LeMunyanCD,WhiteW,NybergE,ChristianJJ.Designofaminiatureradiotransmitterforuseinanimalstudies.JournalofWildlifeManagement,1959,23(1):107-110.
[17]LövyM,klíbaJ,umberaR.Spatialandtemporalactivitypatternsofthefree-livinggiantmole-rat(Fukomysmechowii),thelargestsocialbathyergid.PlosOne,2013,8(1):1-9.
[18]EbenspergeraLA,SobreroR,CamposV,GiannoniSM.Activity,rangeareas,andnestingpatternsintheviscacharat,Octomy smimax.JournalofAridEnvironments,2008,72:1174-1183.
[19]FrancescoliG.Tuco-tucos’vocalizationoutputvariesseasonally(Ctenomys pearsoni;Rodentia,Ctenomyidae):Implicationsforreproductivesignaling.ActaEthologica,2011,14(1):1-6.
[20]ConniorMB,RischTS.HomerangeandsurvivaloftheOzarkpocketgopher(Geomys bursarius ozarkensis)inArkansas.AmericanMidlandNaturalist,2010,164(1):80-90.
[21]BurtWH.Territorialityandhomerangeconceptsasappliedtomammals.JournalofMammalogy,1943,24:346-352.
[22]CutreraAP,MoraMS,AntenucciCD,VassalloAI.Intra-andinterspecificvariationinhome-rangesizeinsympatrictuco-tucos,Ctenomys australisandC.talarum.JournalofMammalogy,2010,91(6):1425-1434.
[23]AebischerNJ,RobertsonPA,KenwardRE.Compositionalanalysisofhabitatusefromanimalradio-trackingdata.Ecology,1993,74(5):1313-1325.
[24]KieJG,BaldwinJA.Calhome:Aprogramforestimatinganimalhomeranges.WildlifeSocietyBulletin,1996,24(2):342-344.
[25]HoogePN,EichenlaubB,SolomonEK.AnimalMovementextensiontoArcView,version2.04beta.AlaskaScienceCenter-BiologicalScienceOffice.USGeologicalSurvey,Anchorage,Alaska.http://www.absc.usgs.gov/glba/gistools,1999.
[26]McNabBK.Bioenergeticsandthedeterminationofhome-rangesize.AmericanNaturalist,1963,894:133-140.
[27]LunaF,AntinuchiCD.Energyanddistributioninsubterraneanrodents:SympatrybetweentwospeciesofthegenusCtenomys.ComparativeBiochemistryandPhysiology,2007,147(4):948-954.
[28]FisherDO,OwensIPF.Femalehomerangesizeandtheevolutionofsocialorganizationinmacropodmarsupials.JournalofAnimalEcology,2000,69:1083-1098.
[29]KomersPE,BrothertonPNM.Femalespaceuseisthebestpredictorofmonogamyinmammals.ProceedingsoftheRoyalSocietyB:BiologicalSciences,1997,264:1261-1270.
[30]EbenspergerLA,BlumsteinDT.Socialityinnewworldhystricognathrodentsislinkedtopredatorsandburrowdigging.BehavioralEcology,2006,17:410-418.
[31]KenwardRE,SouthAB,WallsSS.Ranges6,Version1.2.2003.Wareham,UnitedKingdom:AnatrackLtd.,2003.
[32]HayesLD,CheshA,EbenspergerLA.Ecologicalpredictorsofrangeareasanduseofburrowsystemsinthediurnalrodent,Octodon degus.Ethology,2007,113:155-165.
[33]AndreassenHP,ImsRA,StensethNC,YoccozNG.Investigatingspaceusebymeansofradiotelemetryandothermethods:Amethodologicalguide.In:StensethNC,ImsRA.(eds).TheBiologyofthelemmings.London:AcademicPress,1993:589-618.
[34]EndriesMJ,AdlerGH.Spacingpatternsofatropicalforestrodent,thespinyrat(Proechimys semispinosus)inPanama.JournalofZoology,2005,265(2):147-155.
[35]ArgamasoSM,FroehlichAC,MccallMA,NevoE,ProvencioI,FosterRG.Photopigmentsandcircadiansystemsofvertebrates.BiophysicalChemistry,1995,56:3-11.
[36]RezendeEL,CortésA,BacigalupeLD,NespolodRF,BozinovicF.Ambienttemperaturelimitsabove-groundactivityofthesubterraneanrodentSpalacopus cyanus.JournalofAridEnvironments,2003,55(1):63-74.
[37]HalleS,StensethNC.ActivityPatternsinSmallMammals:AnEcologicalApproach.Berlin:Springer-Verlag,2000:67-90.
[38]BennettNC.Thelocomotoractivitypatterninafunctionallycompletecolonyofthecommonmole-ratCryptomys hottentotusfromSouthAfrica.JournalofZoology(London),1992,228:435-443.
[39]BozinovicF,ContrerasLC.Basalrateofmetabolismandtemperatureregulationoftwodesertherbivorousoctodontidrodents:Octomys mimaxandTympanoctomys barrerae.Oecologia,1990,84:567-570.
[40]RandallJA.Convergencesanddivergencesincommunicationandsocialorganizationofdesertrodents.AustralianJournalofZoology,1994,42(4):405-433.
[41]RandallJA.Environmentalconstraintsandtheevolutionofsocialityinsemifossorialdesertrodents.In:WolffJO,ShermanPW.(eds).RodentSocieties:AnEcologicalandEvolutionaryPerspective.Chicago:UniversityofChicagoPress,2007:368-379.
[43]LunaF,AntinuchiCD,BuschC.RhythmsoflocomotoractivityandburrowuseunderseminaturalconditionsinCtenomys talarum(Rodentia,Octodontidae).RevistaChilenadeHistoriaNatural,2000,73(1):39-46.
[44]UrrejolaD,LaceyEA.Dailyactivitypatternsoffree-livingcururos(Spalacopus cyanus).JournalofMammalogy,2005,86(2):302-308.
[45]KasMJH,EdgarDM.Anonphoticstimulusinvertsthediurnal-nocturnalphasepreferenceinOctodon degus.JournalofNeuroscience,1999,19(1):328-333.
[46]KenagyGJ,NespoloRF,VásquezRA.Dailyandseasonallimitsoftimeandtemperaturetoactivityofdegus.RevistaChilenadeHistoriaNatural,2002,75(3):567-581.
[47]RadoR,ShanasU,ZuriI,TerkelJ.Seasonalactivityintheblindmolerat(Spalaxehrenbergi).CanadianJournalofZoology,1993,71(9):1733-1737.
[48]HickmanGC.Adaptivenessoftunnelsystemfeaturesinsubterraneanmammalburrows.In:NevoE,ReigOA.(eds).EvolutionofSubterraneanMammalsattheOrganismalandMolecularLevels.NewYork:Wiley-Liss,1990,335:185-210.
[49]LaceyEA,PattonJL,CameronGN.Spatialandsocialsystemsofsubterraneanrodents.In:LaceyEA,PattonJL,CameronGN.(eds).LifeUnderground:TheBiologyofSubterraneanRodents.Chicago:UniversityofChicagoPress,2000:255-296.
[50]LaceyEA,BraudeSH,WieczorekJR.SolitaryburrowusebyadultPatagoniantuco-tucos(Ctenomys haigi).JournalofMammalogy,1998,79(3):986-991.
[51]TassinoB,EstevanI,GarberoRP,AltesorP,LaceyEA.SpaceusebyRíoNegrotuco-tucos(Ctenomys rionegrensis):Excursionsandspatialoverlap.MammalianBiology,2010,76(2):143-147.
[52]BurdaH,BrunsV,MüllerM.Sensoryadaptationsinsubterraneanmammals.ProgressinClinicalandBiologicalResearch,1990,335:269-293.
[53]NarinsPM,LewisER,JarvisJJ,OriainJ.TheuseofseismicsignalsbyfossorialsouthernAfricanmammals:Aneuroethologicalgoldmine.BrainResearchBulletin,1997,44(5):641-646.
[54]李金钢,王廷正,何建平,闵一建.甘肃鼢鼠的震动通讯.兽类学报,2001,21(2):153-154.
LiJG,WangYZ,HeJP,MinYJ.SeismiccommunicationinsubterraneanGansuzokor(Myospalax cansus).ActaTheriologicaSinica,2001,21(2):153-154. (inChinese)
[56]NevoE,HethG,PrattH.Seismiccommunicationinablindsubterraneanmammal:Amajorsomatosensorymechanisminadaptiveevolutionunderground.PNAS,1991,88(4):1256-1260.
[57]周建伟,花立民,左松涛,纪维红.3种无线发射器在高原鼢鼠生态学研究中的效果测试.中国媒介生物学及控制,2013,24(6):486-490.
ZhouJW,HuaLM,ZuoST,JiWH.FieldevaluationofthreetypesofradiotransmittersinecologicalstudyonplateauzokorMyospalax baileyi.ChineseJournalofVectorBiologyandControl,2013,24(6):486-490.(inChinese)
[58]孙岳,张雁云.无线电遥测技术在动物学研究中的应用.北京师范大学学报:自然科学版,2009,45(3):268-274.
SunY,ZhangYY.Radiotrackinginzoologicalresearch.JournalofBeijingNormalUniversity:NaturalScienceEdition,2009,45(3):268-274.(inChinese)
[59]KenwardRE.AManualforWildlifeRadioTagging.London:AcademicPress,2001:211-243.
[60]WiebeKL.Delayedtimingasastrategytoavoidnest-sitecompetition:Testingamodelusingdatafromstarlingsandflickers.Oikos,2003,100(2):291-298.
[62]TheuerkaufJ,RouysS,ChatreauC.Mortalityofradio-trackedwildratsinrelationtotransmitterweightandresilienceoftransmittersinrelationtotheirdesign.RoyalSocietyofNewZealand,2007,37(3):85-90.
[63]梁东升.黑龙江垦区森林鼠害的发生原因与防治措施.现代化农业,2006,(6):42-43.
LiangDS.ThecausesandpreventionmeasuresofrodentinHeilongjiangreclamationarea.ModernizingAgriculture,2006(6):42-43.(inChinese)
[64]张兴禄,李广.高原鼠兔和高原鼢鼠在高寒草甸生态系统的作用.草业科学,2015,32(5):816-822.
ZhangXL,LiG.Effectsofrodentsactivitiesongrazinglandandecosysteminalpinemeadow.PrataculturalScience,2015,32(5):816-822.(inChinese)
[65]周延山,花立民,纪维红,楚彬,刘丽.鼢鼠亚科繁殖特性研究进展.草业科学,32(6):1010-1016.
ZhouYS,HuaLM,JiWH,ChuB,LiuL.Areviewaboutreproductivecharacteristicsofzokors.PrataculturalScience,32(6):1010-1016.(inChinese)
[66]刘丽,纪维红,花立民,楚彬,周延山.分子标记在地下啮齿类动物扩散中的应用.草业科学,2015,32(4):612-619.
LiuL,JiWH,HuaLM,ChuB,ZhouYS.Areviewofmolecularmarkersofsubterraneanrodentsdispersal.PrataculturalScience,2015,32(4):612-619.(inChinese)
[67]KoivulaM,KorpimäkiE.Doscentmarksincreasepredationriskofmicrotinerodents?Oikos,2001,95:275-281.
[68]HickmanGC.Alive-trapandtrappingtechniqueforfossorialmammals.SouthAfricanJournalofZoology,1979,14:9-12.
[69]EmsensWJ,SusdbeekL,HirschBT,KaysR,WinkelhagenAJS,JansenPA.Effectsoffoodavailabilityonspaceandrefugeusebyaneotropicalscatterhoardingrodent.Biotropica,2013,45(1):88-93.
[70]ConniorMB,RischTS.Livetrapforpocketgophers.SouthwestNaturalist,2009,54:100-103.
[71]HuaLM,JiWH,ZhouJW,ZuoST.Alivetrapandtrappingtechniqueforsubterraneanzokors(Rodentia).Mammalia,2015,79(4):487-490.
(责任编辑苟燕妮)
Researchprogressofradiotrackingtechnologyinsubterraneanrodents
JiCheng-peng,HuaLi-min,YangSi-wei,ZhouYan-shan,ChuBin
(CollegeofRangelandScienceofGansuAgricultureUniversity,Lanzhou730070,China)
Thesubterraneanrodentsareengineersofterrestrialecosystem.However,itisverydifficulttodirectlystudythesubterraneanrodentsbecausetheyliveunderground,whichtakesomechallengesforstudyingtheirbehaviorandbioecology.Withthedevelopmentofradiotechnology,theminiandlightradiotransmitterhavemadeitpossibleforstudyingthesubterraneanrodents.Manyforeignerscientistshavesuccessfullyusedtheradiotrackingontheborrowingsystems,dailymovement,homerangedynamicandsocialstructureofsubterraneanrodents.However,thistechnologywaslessusedonthesubterraneanrodentsinChina.Thispapersummarizedtheresearchprogressofradiotrackingonsomerodentspecieswhichlivedunderground.ThepurposeistoprovidesomereferencesforstudyingthesubterraneanrodentsinChinainthefuture.
radiotracking;subterraneanrodents;behavior;ecology
HuaLi-minE-mail:hualm@gsau.edu.cn
10.11829/j.issn.1001-0629.2015-0598
2015-11-04接受日期:2016-04-06
国家自然基金项目(31460635);农业部公益性行业科研专项(201203041)
姬程鹏(1990-),男,河南洛阳人,在读硕士生,主要从事草地保护研究。E-mail:jichengp321@163.com
花立民(1971-),男,甘肃临洮人,教授,博士,主要从事草地保护研究。E-mail:hualm@gsau.edu.cn
动物生产层
S443;S12
A
1001-0629(2016)9-1859-09*
姬程鹏,花立民,杨思维,周延山,楚彬.无线电追踪技术在地下啮齿动物研究中的应用.草业科学,2016,33(9):1859-1867.
JiCP,HuaLM,YangSW,ZhouYS,ChuB.Researchprogressofradiotrackingtechnologyinsubterraneanrodents.PrataculturalScience,2016,33(9):1859-1867.
猜你喜欢
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
海外星云(2021年6期)2021-10-14
无线互联科技(2021年4期)2021-04-21
兽医导刊(2020年3期)2020-12-30
动漫星空(兴趣百科)(2020年5期)2020-06-10
小猕猴智力画刊(2019年3期)2019-04-19
防护林科技(2019年7期)2019-03-25
电子制作(2018年23期)2018-12-26
电子制作(2018年19期)2018-11-14
