
兰州北山3个灌木优势种光合特性
2016-10-20 09:47徐春华
草业科学 2016年9期
张 华,徐春华
(1.西北师范大学地理与环境科学学院,甘肃 兰州 730070;2.兰州大学草地农业系统国家重点实验室,甘肃 兰州 730000)
兰州北山3个灌木优势种光合特性
张 华1,2,徐春华1
(1.西北师范大学地理与环境科学学院,甘肃 兰州 730070;2.兰州大学草地农业系统国家重点实验室,甘肃 兰州 730000)
本研究对兰州北山植被优势种柽柳(Tamarix austromongolica)、侧柏(Platycladus orientalis)和柠条(Caragana korshinskii)的光合特性和微气象环境进行了测定,分析了其日变化特点,并通过灰色关联度分析方法探讨了净光合速率(Pn)与生理因素及环境因素的关联程度。结果表明,1)自然条件下,3种植物的Pn日变化均为不对称的双峰曲线,存在明显的光合“午休”现象,这是由于午间高光强、高温构成光抑制现象。2)生理因素分别与3种植物Pn的关联度由大到小依次为,柽柳:胞间CO2浓度(Ci)>叶温(Tl)>蒸腾速率(Tr) >气孔导度(Gs)>饱和水汽压(VPD)>气孔限制值(Ls),侧柏:Tr>Tl>VPD>Ci>Ls>Gs,柠条:Tr>Tl>Ci>Gs>VPD>Ls;生理因素中Ci对柽柳Pn的影响最大,Tr对侧柏和柠条Pn的影响最大。3)环境因素分别与3种植物Pn的关联度由大到小依次为,柽柳:光合有效辐射(PAR)>相对湿度(RH)>土壤温度(Ts)>土壤湿度(SM)>大气温度(Ta)>大气CO2浓度(Ca),侧柏:PAR>SM>Ta>Ts>RH>Ca,柠条:RH>PAR>SM>Ta>Ca>Ts;环境因素中PAR对柽柳和侧柏Pn的影响最大,RH则对柠条Pn的影响最大。
兰州北山;光合特性;日变化;灰色关联度分析;生理参数;环境参数
兰州北山是祁连山东脉的延伸,植被稀疏[1],由于气候变化和人类活动的加剧,水土流失日益严重,已成为黄河的主要沙源区之一,影响着黄河流域生态系统的健康和兰州城市的人居环境。为恢复生态环境,国家采取了多种治理措施,加大了对兰州南北两山植被生态恢复造林工程的投资力度[2-3]。柽柳(Tamarix austromongolica)、侧柏(Platycladus orientalis)和柠条(Caragana korshinskii)以其耐旱、抗盐碱等特点成为兰州北山的主要绿化植物。植物的光合作用是植物生产过程中物质积累与生理代谢的基本过程,也是评价植物第一生产力的标准之一,受生理因素和环境因素的共同影响[4-5]。
灰色关联度分析方法是将因素之间发展的相异或相似程度作为衡量两个因素关联程度的一种方法,能很好地反映各个因素间的相对重要程度或优劣关系[6-8]。目前,关于植物光合特性的研究大多都运用对样本量有较高要求的相关分析、回归分析等方法分析光合作用与其影响因素间的关系[9-11],运用灰色关联度分析方法综合分析光合作用影响因素的研究较少。因此,本研究通过分析兰州北山柽柳、侧柏和柠条光合作用日变规律,运用灰色关联度分析方法综合分析生理因素和环境因素对3种植物的光合作用的影响,剖析3种植物的光合作用机理,进而为兰州北山绿化植被选种提供理论与实践参考。
1 研究区概况
兰州北山属于黄土高原沟壑区,位于黄河兰州段以北,东起城关区青白石乡张儿沟,西至西固区达川乡达家沟,南临黄河,林草覆被率低、植被稀疏。地势北高南低,由西北向东南方向倾斜,海拔1 560~2 067m,坡度大于30°。属于典型的温带大陆性气候区,年均气温8.9 ℃,≥10 ℃年积温为3 242.0 ℃·d,年均降水量328mm左右,集中于7-9月,无霜期190d。土壤以淡灰钙土为主,土壤pH值8.0~8.6,有机质含量0.5%。原生植被以荒漠草原植被为主,由多年旱、丛生的低矮禾草,旱生灌木和小半灌木组成,人工绿化植被主要有柽柳、侧柏、柠条、杨树(Populus)、紫穗槐(Amorpha fruticosa)、油松(Pinus tabuliformis)等[12]。
2 材料与方法
2.1试验方法
本研究于2014年5月上、中和下旬各选择晴朗无云的3d进行观测,时间为08:00-18:00。每天测定时选择生长状况相似的柽柳、侧柏和柠条各3株。每株中选取朝向相同、叶位相同、长势相近、无病虫害的叶片进行测定,同一叶片每次重复测量3次,共9次,每1h测定一轮,运用拍照法[13]计算所测部分的叶面积。
在自然条件下利用Ci-340便携式手持光合测量系统,所测参数包括净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)、饱和水汽压差(VPD)、叶温(Tl)等;WatchDog2000自动气象站,所测参数包括光合有效辐射(PAR)、大气CO2浓度(Ca)、大气温度(Ta)、土壤温度(Ts)、相对湿度(RH);WateraScoutSM100土壤湿度传感器测定了土壤湿度(SM)。根据所测参数计算水分利用效率(WUE=Pn/Tr)和气孔限制值(Ls=1-Ci/Ca)。
2.2灰色关联度分析方法
灰色关联度分析法是一种多因素统计分析方法,它以各因素的样本数据为依据,用灰色关联度来描述因素间关系的强弱、大小和次序[14]。该方法定量考虑多个因素的作用,得出具有可比性的综合性指标,从而提高综合评估的准确性和有效性,避免了人为评判的主观性。灰色关联度越大,说明比较项与参考项的发展趋势越接近,即参考数列受比较数列影响更大[15]。
2.3数据处理
先利用相关分析计算各微环境因素的相关系数,并进行显著性检验;然后运用灰色关联分析方法对Pn与其影响因素的关联度进行排序,得出Pn的影响因素排序。所有分析均在Excel2007和SPSS17.0中完成。
3 结果与分析
3.1主要环境因素日变化
环境因素主要包括PAR、Ca、Ta、Ts、RH和SM。PAR日变化呈现明显的单峰曲线,从08:00-13:00随着太阳高度的增大而增强,13:00左右达到最大,为2 035mol·(m2·s)-1,之后随着太阳的西落而迅速降低,17:00开始降幅变缓。Ca的日变化总体呈现下降趋势(图1)。随着PAR变化,Ta也表现为单峰曲线,最大值出现在14:00,为38.5 ℃(图1),与PAR相关性较为显著(r=0.732,P<0.05)(表1),并且有滞后性。
Ts日变化呈现不明显的单峰曲线,从08:00开始升高,在15:00达到最大,之后开始降低,增加幅度大于降低幅度,对Ta有1h的滞后效应,且Ts与Ca呈极显著负相关(r=-0.914,P<0.001)。RH的日变化总体呈下降趋势,在PAR达最大时降到最低,在14:00有小幅上升,RH与Ta呈负相关关系(r=-0.626,P<0.05),与Ts呈极显著负相关关系(r=-0.962,P<0.001)。SM的变化为先升后降,13:00最大,升降幅度均不大,与PAR呈极显著正相关关系(r=0.937,P<0.001)(图1,表1)。
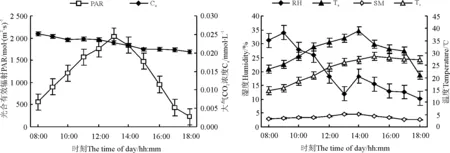
图1 环境因素日变化Fig.1 Diurnal changes of environmental factor
注:PAR,光合有效辐射;Ca,大气CO2浓度;RH,空气相对湿度;SM,土壤相对湿度;Ta,大气温度;Ts,土壤温度。下同。
Note:PAR,photosyntheticallyactiveradiation;Ca,atmosphericCO2concentration;RH,airrelativehumidity;SM,soilmoisture;Ta,airtemperature;Ts,soiltemperature.Thesamebelow.

表1 环境因素之间的相关系数Table 1 Correlation coefficient among environmental factors
注:*、**、***分别表示 在0.05、0.01和0.001水平上显著。
Note: *, **and***indicatesignificantcorrelationat0.05, 0.01and0.001level,respectively.
3.23种植物光合特性日变化
3种植物的Pn日变化均呈明显不对称的双峰曲线(图2)。侧柏和柠条Pn的第1峰值在10:00左右出现,而柽柳Pn的第1峰值出现的时间为11:00,侧柏和柠条第1峰值出现的时间较柽柳提前了1h;3种植物的第2峰值都出现在14:00左右。12:00-13:00之间,过高的光强和温度对3种植物的光合作用产生了抑制,Pn均处于谷值。可以看出,3种植物均存在明显的光合“午休”现象[16-17],而且侧柏和柠条的午休强度大于柽柳。经过光合“午休”后,柽柳的光合作用有所增强,而侧柏和柠条的光合能力并未恢复到午休前的最高水平。
蒸腾作用是反映植物水分代谢的一个重要生理指标,Tr能够反映蒸腾作用的强弱。3种植物Tr的日变化曲线均呈双峰型,其中,柽柳Tr变化不明显(图2)。
3种植物的Ci日变化在波动中呈现下降趋势(图2),其中,柽柳和柠条Ci变化幅度较大,柽柳的Ci在13:00明显升高,柠条的Ci在14:00左右明显升高。3种植物的Ls日变化都呈双峰曲线(图2),在08:00时较高,然后下降,侧柏Ls从09:00开始上升,柽柳和柠条在10:00开始上升,均在12:00达到第1个峰值。随着PAR、Ta在全天呈现单峰曲线(图1),Tl与Ta变化大致相同(图2),其中,侧柏的Tl峰值出现在13:00左右,柽柳和柠条Tl的峰值在14:00左右出现。Ta升高引起环境饱和水汽压增大,进而加大叶片表面蒸发能力使叶片VPD增大(图2)。3种植物Gs日变化曲线大致呈“L”型(图2),其中08:00是全天的最大值点,08:00-11:00内下降趋势相对较大,11:00以后变化幅度相对减缓,并且午后均出现了相对较小的上升趋势。
水分利用效率WUE是反映植物耐旱性的一个有效指标,它显示植物有效利用水分的能力,即在相同条件下,水分利用效率高的植物的抗旱能力较强[18]。本研究中3种植物WUE的变化趋势和Pn的变化趋势大致相同一致,总体来看,柽柳的WUE明显高于侧柏和柠条,说明柽柳的抗旱能力强于侧柏和柠条(图2)。3种植物的WUE日均值表现为柽柳(2.56molCO2·mmol-1H2O)>柠条(1.13molCO2·mmol-1H2O)>侧柏(0.42molCO2·mmol-1H2O),这主要是由于3种植物在根系分布上的不同导致了不同的水分利用方式。
3.3Pn与生理因素和环境因素的灰色关联分析
灰色关联度计算结果表明(表2),生理因素对3种植物Pn的影响均较大,但影响程度存在差异。生理因素与3种植物Pn的关联度由大到小排序,柽柳表现为Ci>Tl>Tr>Gs>VPD>Ls;侧柏表现为Tr>Tl>VPD>Ci>Ls>Gs;柠条表现为Tr>Tl>Ci>Gs>VPD>Ls。可见,生理因素中,Ci对柽柳Pn的影响最大(关联度为0.859),是影响柽柳Pn的重要生理因素;Tr对侧柏和柠条Pn的影响最大(关联度分别为0.781、0.679),是影响侧柏和柠条Pn的重要生理因素。其它生理因素中,Tl对3种植物Pn的影响均较大(关联度分别为0.847、0.766、0.654)。
环境因素与3种植物Pn关联度从大到小的排列顺序(表3),柽柳表现为PAR>RH>Ts>SM>Ta>Ca;侧柏表现为PAR>SM>Ta>Ts>RH>Ca;柠条表现为RH>PAR>SM>Ta>Ca>Ts。其中,PAR与柽柳和侧柏Pn的关联度最大(分别为0.617、0.733),表明环境因素中,PAR对柽柳和侧柏光合作用的影响最大;RH与柠条Pn的关联度(0.663)较大,表明环境因素中RH对柠条光合作用的影响最大。其它环境因素中,RH对柽柳Pn的影响最大(关联度为0.551),是柽柳Pn的重要环境影响因素;SM对侧柏Pn的影响最大(关联度为0.624),是侧柏Pn的重要环境影响因素;PAR对柽柳Pn的影响最大(关联度为0.632),是柠条光合作用的重要环境影响因素。
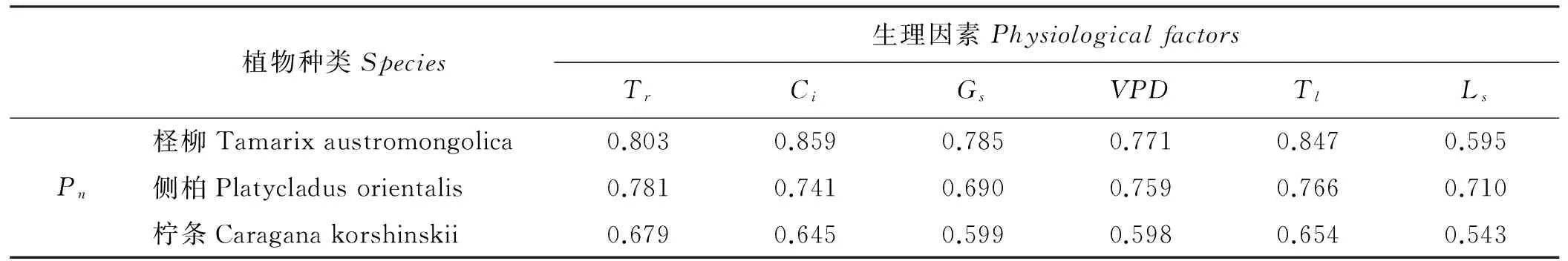
表2 Pn与生理因素的关联度Table 2 Relation Grades between Pn and physiological factors
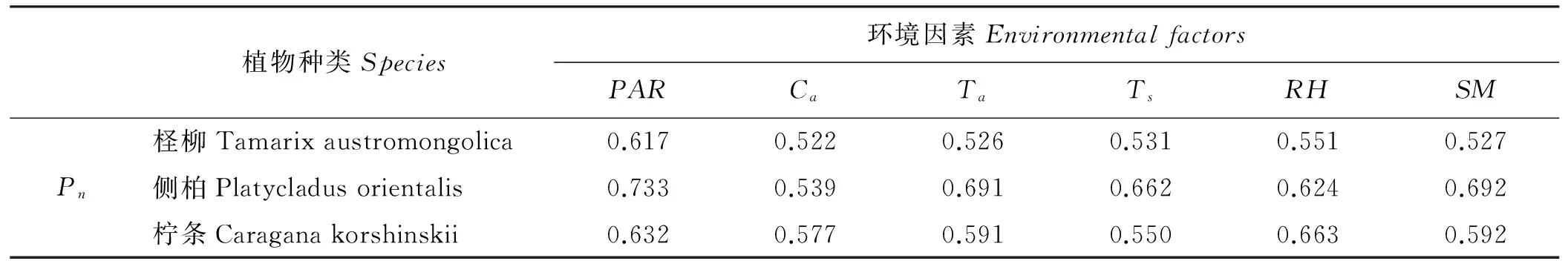
表3 Pn与环境因素的关联度Table 3 Relation Grades between Pn and environmental factors
4 讨论与结论
4.1讨论
植物净光合速率日变化曲线主要有双峰型或单峰型两种[19-20]。本研究中3种植物净光合速率的日变化曲线均为不对称的双峰曲线,存在明显的光合“午休”现象,这与大多数植物的光合特性一致[21]。光合“午休”现象也是C3植物的共同特征[22]。Farquhar和Sharkey[23]认为,影响净光合速率下降有气孔限制和非气孔限制两方面的因素,如果净光合速率下降的同时胞间CO2浓度降低,气孔导度有所增大,此时净光合速率降低主要是由气孔关闭引起的;而如果净光合速率下降的同时胞间CO2浓度上升,但气孔导度下降,表明净光合速率降低主要是由非气孔因素引起的。本研究中, 12:00-14:00期间,侧柏净光合速率降低,胞间CO2浓度也降低,气孔导度也随之降低;随着净光合速率降低,柽柳和柠条的胞间CO2浓度有所上升,但气孔导度有所降低。这主要是由于午间高温、强辐射使叶片蒸腾加强,造成暂时的水分亏缺,叶温升高,刺激了气孔,气孔导度降低, 气孔限制值明显升高,从而防止了水分的过分流失,同时使CO2吸收量减少,导致光合速率降低,而非气孔因素所致,这和王珊珊等[24]、刘建峰等[25]、胡文超等[26]的研究结果一致。
在自然条件下,内部生理因素和外界环境因素对植物光合作用的影响不是单一和孤立的,而是相互联系、相互制约、综合影响的[27-28]。张锦春等[29]认为,由于环境因素的不同引起植物光合特性的多样性变化,环境因素起着主导作用。各环境因素之间也相互影响,相互制约。强烈的太阳辐射是引起一天中温度、湿度等一系列环境因素变化的根本原因[30]。光合有效辐射在13:00达到最大,而大气温度在14:00达到最大,大气温度对光合有效辐射有1h的滞后效应。午间大气CO2浓度和相对湿度持续降低,因为午间过高的大气温度引起空气密度降低,进而使得午间相对湿度和大气CO2浓度降低。此外,植物光合作用吸收CO2也是大气CO2浓度降低的一个重要原因。在影响植物光合作用的环境因素中,光合有效辐射是驱动因素,其变化驱动了大气温度和土壤温度的变化,而相对湿度、土壤湿度与大气CO2浓度的变化受大气温度的影响较大。
土壤水分状况是决定植物光合作用的关键因素,对植物的生存和演替有重要作用。植物内部因素如叶片气孔导度、气孔阻力等是植物长期适应多变的环境和土壤而形成的一类自身调节因素,主要通过环境因素而间接影响植物的光合生理变化。通过灰色关联度分析发现,环境因素中,光合有效辐射与柽柳和侧柏净光合速率的关联度最大,是影响柽柳和侧柏光合作用的重要环境因素。生理因素中,蒸腾速率与侧柏和柠条净光合速率的关联度最大,是影响它们光合作用的重要生理因素。植物的蒸腾作用是通过气孔完成的。在干旱环境下,当午间光合有效辐射增大时,气孔变大,蒸腾速率随之增强,导致植物单位面积叶片蒸腾失水增多,从而降低净光合速率。
植物叶片温度在一定程度上取决于空气温度,影响光合作用的生化反应和饱和水汽压[31-32],从而影响蒸腾和叶片失水、气孔开闭,以至于影响CO2和水汽的进出、光合磷酸化酶活性、RuBp羧化酶活性、RuBp的再生[33-34]。本研究分析发现,午间叶温均较高,3种植物的净光合速率均下降,且叶温与3种植物净光合速率的关联度均排在第二,对3种植物光合作用的影响也较大。
除上述因素外,土壤水分是影响植物体内水分状况的重要因素,从而直接影响植物光合作用的进行。在自然条件下,植物体内的水分状况随土壤水分的变化而异,从而引起光合活性的变化。水分胁迫导致光合作用下降是干旱条件下作物减产的重要原因,土壤水分亏缺对光合作用的影响可以通过气孔导度进行,也可以直接影响到叶肉细胞的光合能力[35]。本研究通过灰色关联分析发现,土壤湿度与侧柏净光合速率 的关联度排在第二,表明土壤湿度对侧柏光合作用的影响也较大。
灰色关联度分析法在农业科学研究中被广泛应用,该方法具有直观、准确和便捷等特点。但灰色关联度分析法仅能反映各性状问的关联紧密程度,不能同时反映各因素间的作用方向和直接、间接效应。因此,进一步的分析还应结合多元回归分析和相关分析等方法。
4.2结论
各环境因素紧密相关。光合有效辐射变化直接引起大气温度的变化,而大气温度对大气CO2浓度和相对湿度有明显影响。大气温度对光合有效辐射,大气CO2浓度和相对湿度对大气温度分别有1h滞后效应,在光合有效辐射的驱动下各环境因子对净光合速率共同起作用。
3种植物的水分利用效率日均值表现为柽柳(2.56molCO2·mmol-1H2O)>柠条(1.13molCO2·mmol-1H2O)>侧柏(0.42molCO2·mmol-1H2O),柽柳的水分利用效率明显高于侧柏和柠条,说明柽柳的抗旱能力强于侧柏和柠条。 3种植物光合“午休”现象明显,且均非气孔因素所致。
生理因素中,胞间CO2浓度与蒸腾速率对柽柳净光合速率的影响较大,蒸腾速率和叶温对侧柏和柠条净光合速率的影响较大;环境因素中,光合有效辐射和相对湿度对柽柳和柠条净光合速率的影响较大,光合有效辐射和土壤湿度对侧柏净光合速率的影响较大。
< class="emphasis_italic">References
:
[1]孙宏义,钟芳,李芳,董海涛.油蒿在兰州南北两山生态环境建设中的适应性评价研究.冰川冻土,2012,34(6):1517-1525.
SunHY,ZhongF,LiF,DongHT.EvaluationoftheadaptabilityofArtemisia ordosicacultivatedinLanzhou’sNorthSouthHills.JournalofGlaciologyandGeocryology,2012,34(6):1517-1525.(inChinese)
[2]谭明亮,段争虎,陈小红,张瑞军.半干旱区城市人工森林生态系统服务价值评估——以兰州市南北两山环境绿化工程区为例.中国沙漠,2012,32(1):219-225.
TanML,DuanZH,ChenXH,ZhangRJ.Ecologicalservicevalueassessmentofurbanartificialforestecosyteminsemi-aridregions:AcasestudyinLanzhouCity,China.JournalofDesertResearch,2012,32(1):219-225.(inChinese)
[3]段翰晨,颜长珍,马如兰,庞国锦,蒋小荣.兰州市南北两山生态建设效应的遥感监测.中国沙漠,2011,31(2):456-463.
DuanHC,YanCZ,MaRL,PangGJ,JiangXR.EcosystemconstructioneffectsinSouthernandNorthernMountainsofLanzhoubyremotesensingmonitoring.JournalofDesertResearch,2011,31(2):456-463.(inChinese)
[4]李林锋.4种湿地植物光合作用特性的比较研究.西北植物学报,2008,28(10):2094-2102.
LiLF.Comparativestudyonphotosyntheticcharacteristicsofhydrophytesinconstructedwetland.ActaBotanicaBoreali-OccidentaliaSinica,2008,28(10):2094-2102.(inChinese)
[5]TuittilaES,VasanderH,LaineL.SensitivityofCsequestrationinreintroducedsphagumtowater-levelvariationinacutawaypeatland.RestorationEcology,2004,12(4):483-493.
[6]林培松,尚志海.韩江流域典型区主要森林类型土壤肥力的灰色关联度分析.生态与农村环境学报,2009,25(3):55-58.
LinPS,ShangZH.GreyrelationalanalysisofsoilfertilityofmainforestsoilsintypicaldistrictofHanjiangRiverBasin.JournalofEcologyandRuralEnvironment,2009,25(3):55-58.(inChinese)
[7]杨秀芳,陈玲玲,乌艳红,娜日苏,吕宁,梁庆伟.应用灰色关联度综合评价26个青贮玉米的生产性能.草业科学,2012,29(1):105-111.
YangXF,ChenLL,WuYH,Narisu,LyuN,LiangQW.Acomprehensiveevaluationofthegrowthperformanceof26silagemaizegenotypesusinggreycorrelativedegreeanalysis.PrataculturalScience,2012,29(1):105-111.(inChinese)
[8]史永良,杜英,张爱宁,张建韬.灰色关联度分析的甘肃马铃薯淀粉产业竞争力研究.浙江农业学报,2012,24(1):162-168.
ShiYL,DuY,ZhangAN,ZhangJT.CompetitivenessstudyofpotatostarchindustrybasedongraycorrelationanalysisinGansuProvince.ActaAgriculturaeZhejianggensis,2012,24(1):162-168.(inChinese)
[9]吴统贵,曾广泉,肖杨根,姚婕,虞木奎,成向荣,王臣.湿地松林下6种光合光合日变化及其与环境因素的关系.南京农业大学学报,2011,35(5):135-138.
WuTG,ZengGQ,XiaoYG,YaoJ,YuMK,ChengXR,WangC.DailyvariationofphotosynthesisofsixtreespeciesunderPinus elliotiiforestandtheirrelationswithenvironmentalfactors.JournalofNanjingForestryUniversity:NaturalScienceEdition,2011,35(5):135-138.(inChinese)
[10]池永宽,熊康宁,王元素,张锦华,董颖苹.贵州石漠化地区灰绿藜和鹅肠菜光合日动态.草业科学,2014,31(11):2119-2124.
ChiYK,XoingKN,WangYS,ZhangJH,DongYP.PhotosyntheticdailydynamicsofChenopodium glaucumandMyosoton aquaticuminGuizhourockydesertificationarea.PrataculturalScience,2014,31(11):2119-2124.(inChinese)
[11]康雅茸,张华.兰州银滩黄河湿地不同植物夏季光合日变化.生态学杂志,2014,33(9):2336-2342.
KangYR,ZhangH.DiurnalvariationofphotosynthesisofdifferentplantsinsummerinYintanwetlandofLanzhousectionofYellowRiver.ChineseJournalofEcology,2014,33(9):2336-2342.(inChinese)
[12]武利玉,苏世平,王蕙.兰州南北两山绿化区植物与植被类型初查.中国沙漠,2006,26(4):564-568.
WuLY,SuSP,WangH.PreliminaryinvestigationintoplantandvegetationtypesinafforestationregioninSouthernandNorthernMountainsofLanzhouCity.JournalofDesertResearch,2006,26(4):564-568.(inChinese)
[13]王忠芝,张金瑞.基于图像处理的叶面积测量方法.微计算机应用,2010,31(5):69-72.
WangZZ,ZhangJR.Ameasurementapproachofleafareabaseddigitalimageprocessing.MicrocomputerApplications,2010,31(5):69-72.(inChinese)
[14]孙玉刚.灰色关联度分析及其应用研究.南京:南京航空航天大学硕士论文,2007.
SunYG.Researchongreyrelationalanalysisanditsapplications.MasterThesis.Nanjing:NanjingUniversityofAeronauticsandAstronautics,2007.(inChinese)
[15]王士强,胡银岗,佘奎军,周琳璘,孟凡磊.小麦抗旱相关农艺性状和生理生化性状的灰色关联度分析.中国农业科学,2007,40(11):2452-2459.
WangSQ,HuYG,SheKJ,ZhouLL,MengFL.Grayrelationalgradeanalysisofagronomicalandphys-biochemicaltraitsrelatedtodroughttoleranceinwheat.ScientiaAgriculturaSinica,2007,40(11):2452-2459.(inChinese)
[16]许大全,李德耀,沈允钢,梁国安.田间小麦叶片光合作用“午睡”现象的研究.植物生理学报,1984,10(3):269-276.
XuDQ,LiDY,ShenYG,LiangGA.Onmiddaydepressionofphotosynthesisofwheatleafunderfieldconditions.ActaPhytophysiologiaSinica,1984,10(3):269-276.(inChinese)
[17]耿显胜,肖世奇,葛晓改.植物的光合午休.生物学教学,2010,35(12):59.
GengXS,XiaoSQ,GeXG.Middaydepressionofplants.BiologyTeaching,2010,35(12):59.(inChinese)
[18]曹生奎,冯起,司建华,常宗强,卓玛错,席海洋,苏永红.植物叶片利用效率研究综述.生态学报,2009,29(7):3882-3892.
CaoSK,FengQ,SiJH,ChangZQ,Zhuomacuo,XiHY,SuYH.Summaryontheplantwateruseefficiencyatleaflevel.ActaEcologicaSinica,2009,29(7):3882-3892.(inChinese)
[19]SobradoMA.Relationofwatertransporttoleafgasexchangepropertiesinthreemangrovespecies.Trees,2000,14(5):258-262.
[20]刘小文,王秋霞,郭美霞,白洁,赵云,李小艳,曹坳程,何月秋,肖春,吴国星.紫茎泽兰生殖期光合特性日变化.草业科学,2011,28(1):105-109.
LiuXW,WangQX,GuoMX,BaiJ,ZhaoY,LiXY,CaoAC,HeYQ,XiaoC,WuGX.CharacteristicsofphotosyntheticdiurnalvariationatthereproductiveofEupatorium adenophorum.PrataculturalScience,2011,28(1):105-109.(inChinese)
[21]唐源江,阮志平.董棕和短穗鱼尾葵光合效率日变化及其与生理生态因子的关系.西北植物学报,2012,32(5):995-1001.
TangYJ,RuanZP.Photosyntheticdynamicvariationanditsrelationshipwiththeeco-physiologicalfactorsinCaryota urensandCaryota mitis.ActaBotanicaBoreali-OccidentaliaSinica,2012,32(5):995-1001.(inChinese)
[22]张颖,呼天明.普那菊苣夏季光合速率日变化及其影响因子的研究.西北农业学报,2007,16(5):184-187.
ZhangY,HuTM.StudyonthediurnalchangesofnetphotosyntheticrateandtheimpactfactorsofPunaChicoryinsummer.ActaAgricultureBoreali-OccidentalisSinica,2007,16(5):184-187.(inChinese)
[23]FarquharD,SharkeyD.Stomatalconductanceandphotosynthesis.AnnualReviewofPlantPhysiology,1982,33(1):317-345.
[24]王珊珊,陈曦,王权,包安明,曹晓明,李平衡.新疆古尔班通古特沙漠南缘多枝柽柳光合作用及水分利用的生态适应性.生态学报,2011,31(11):3082-3089.
WangSS,ChenX,WangQ,BaoAM,CaoXM,LiPH.EcologicaladaptabilityofphotosynthesisandwateruseforTamarix ramosissimainthesouthernperipheryofGurbantunggutDesert,Xinjiang.ActaEcologicaSinica,2011,31(11):3082-3089.(inChinese)
[25]刘建峰,杨文娟,史胜青,郭泉水,江泽平.崖柏与侧柏光合特性和叶绿素荧光参数的比较研究.西北植物学报,2011,31(10): 2071-2077.
LiuJF,YangWJ,ShiSQ,GuoQS,JiangZP.ComparisononphotosyntheticcharacteristicsandchlorophyllfluorescenceparametersbetweenThuja sutchuenensisandPlatycladus orientalis.ActaBotanicaBoreali-OccidentaliaSinica,2011,31(10):2071-2077.(inChinese)
[26]胡文超.西北地区18种常见灌木光合特性和光合固碳潜力的比较研究.银川:宁夏大学硕士学位论文,2013.
HuWC.Comparativestudyofthephotosyntheticcharacteristicsandphotosyntheticcarbonsequestrationof18commonshrubsinnorthwestareaofChina.MasterThesis.Yinchuan:NingxiaUniversity,2013.(inChinese)
[27]刘玉华,史纪安,贾志宽,韩清芳.旱作条件下紫花苜蓿光合蒸腾日变化与环境因子的关系.生态学报,2006,26(5):1468-1477.
LiuYH,ShiJA,JiaZK,HanQF.Diurnaldynamicsofphotosyntheticandtranspirationratesofalfalfaunderdryfarmingandtheirrelationshipswithenvironmentalfactors.ActaEcologicaSinica,2006,26(5):1468-1477.(inChinese)
[28]韩忠明,王云贺,韩梅,杨利民.药用植物防风(Saposhnikovia divaricata)不同时期光合作用特性比较.生态学报,2009,29(3):1382-1390.
HanZM,WangYH,HanM,YangLM.ComparativestudyonphotosynthesisofmedicinalplantSaposhnikovia divaricataindifferentperiods.ActaEcologicaSinica,2009,29(3):1382-1390.(inChinese)
[29]张锦春,赵明,张应昌,徐延双.灌溉植被梭梭、白刺光合蒸腾特性及影响因素研究.西北植物学报,2005,25(1):0070-0076.
ZhangJS,ZhaoM,ZhangYC,XuYS.Aresearchbetweenphotosynthetic,transpirationcharacteristicsandimpactofirrigatedvegetationofHaloxylon ammodendronandNitraria tangutorum.ActaBotanicaBoreali-OccidentaliaSinica,2005,25(1):0070-0076.(inChinese)
[30]谢田玲,沈禹颖,邵新庆,高崇岳.黄土高原4 种豆科牧草的净光合速率和蒸腾速率日动态及水分利用效率.生态学报,2004,24(8):1679-1686.
XieTL,ShenYY,ShaoXQ,GaoCY.DifferencesinphotosyntheticratetranspirationrateandwateruseefficiencybetweenfourlegumespeciesontheLoessPlateau.ActaEcologicaSinica,2004,24(8):1679-1686.(inChinese)
[31]MottKA.DostomatarespondtoCO2concentrationsotherthanintercellular.PlantPhysiology,1988,88(1):200-213.
[32]MottKA,ParkhurstDF.Stomatalresponsestohumidityinairandhelox.Plant,Cell&Environment,1991,14(5):509-515.
[33]KhairiMA,HallAE.Temperatureandhumidityeffectonnetphotosynthesisandtranspirationofcitrus.PhysiologialPlantarum,1976,36(2):29-34.
[34]TenhunenJD,SerraAS,HarleyPC,DoughertyRL,ReynoldsJF.FactorsinfluencingcarbonfixationandwaterusebyMediterraneanchlorophyllshrubsduringsummerdrought.Oecologia,1990,82(3):381-393.
[35]陈建.4种灌木植物光合效率对土壤水分的响应过程与机制.济南:山东农业大学硕士学位论文,2008.
ChenJ.Thersponseprocessesandmechanismoffourshrub’sphotosyntheticefficiencyparameterstosoilwater.MasterThesis.Ji’nan:ShandongAgriculturalUniversity,2008.
(责任编辑王芳)
ResearchonphotosynthesischaracteristicsofthreetreebushesinBeishanMountainofLanzhouCity
ZhangHua1,2,XuChun-hua1
(1.CollegeofGeographicandEnvironmentalSciences,NorthwestNormalUniversity,Lanzhou730070,China;2.StateKeyLaboratoryofGrasslandPastoralAgriculturalEcosystem,LanzhouUniversity,Lanzhou730070,China)
ThisstudymainlymeasuredthediurnalchangeofphotosynthesischaracteristicsofTamarix austromongolica, Platycladus orientalisandCaragana korshinskiiintheBeishanMountainofLanzhou,aswellasenvironmentalfactors.Byusingtherelationalgradeanalysisofgreysystemtheory,therelationalgradebetweenNetphotosyntheticrate(Pn),physiologicalandenvironmentalfactorswerecalculated.Theresultsshowed:ThediurnalvariationofPnofthreespeciesshowedasymmetricbimodalpeakcurve,witharemarkablephotosynthetic“middaydepressions”.Itiscausedbythemiddayhighlightintensityandtemperature,ratherthanstomatalfactors.TherelationgradesbetweenPnandphysiologicalfactorswasIntercellularCO2concentration(Ci) >Leaftemperature(Tl) >TranspirationrateTr>Stomatalconductance(Gs) >Saturationvaporpressuredifferencefoliage(VPD) >Stomatallimitatingvalue(Ls)forT. austromongolica,Tr>Tl>VPD>Ci>Ls>GsforP. orientalisandTr>Tl>Ci>Gs>VPD>LsforC. korshinskii.Amongthem,CihasaobviouseffectonPnofT. austromongolicaanditisTrforP. orientalisandC. korshinskii.TherelationgradesbetweenPnanenvironmentalfactorswasPhotosyntheticallyactiveradiation(PAR)>Airrelativehumidity(RH) >Soiltemperature(Ts) >Soilmoisture(SM) >Airtemperature(Ta) >AtmosphericCO2concentration(Ca)forT. austromongolica,PAR>SM>Ta>Ts>RH>CaforP. orientalisandRH>PAR>SM>Ta>Ca>TsforC. korshinskii.Amongthem,PAR,hasadeeplyeffectonPnforP. orientalisandT. austromongolica,whileitisRHforC. korshinskii.
BeishanMountainofLanzhouCity;photosynthesischaracteristics;diurnalchange;greyrelationalanalysis;physiologicalfactors;environmentalfactors
ZhangHuaE-mail:zhanghua2402@163.com
10.11829/j.issn.1001-0629.2015-0599
2015-11-04接受日期:2015-02-26
国家自然科学基金项目( 41461011)
张华(1978-),女,甘肃兰州人,副教授,硕导,博士,主要从事干旱区植被与生态恢复研究。E-mail:zhanghua2402@163.com
Q945.11
A
1001-0629(2016)9-1809-09*
张华,徐春华.兰州北山3个灌木优势种光合特性.草业科学,2016,33(9):1809-1817.
ZhangH,XuCH.ResearchonphotosynthesischaracteristicsofthreetreebushesinBeishanMountainofLanzhouCity.PrataculturalScience,2016,33(9):1809-1817.
猜你喜欢
新农民(2022年22期)2022-11-07
现代农村科技(2021年5期)2021-12-07
乡村科技(2021年16期)2021-10-21
星星·诗歌原创(2021年3期)2021-06-01
中国煤炭工业(2020年1期)2020-12-28
河北林业科技(2020年3期)2020-03-23
当代陕西(2019年7期)2019-04-25
扬子江(2018年1期)2018-01-26
花卉(2017年7期)2017-11-15
花卉(2017年7期)2017-07-20
