
不同氮效率油菜品种碳素累积转运差异及其对油分形成的影响
2016-10-18 11:46:44张晓龙张振华宋海星余佳玲官春云
中国农业科学 2016年18期
张晓龙,张振华,宋海星,余佳玲,官春云
(1湖南农业大学资源环境学院/南方粮油作物协同创新中心/土壤肥料资源高效利用国家工程实验室/农田污染控制与农业资源利用湖南省重点实验室/植物营养湖南省普通高等学校重点实验室,长沙 410128;2国家油料改良中心湖南分中心,长沙 410128)
不同氮效率油菜品种碳素累积转运差异及其对油分形成的影响
张晓龙1,张振华1,宋海星1,余佳玲1,官春云2
(1湖南农业大学资源环境学院/南方粮油作物协同创新中心/土壤肥料资源高效利用国家工程实验室/农田污染控制与农业资源利用湖南省重点实验室/植物营养湖南省普通高等学校重点实验室,长沙 410128;2国家油料改良中心湖南分中心,长沙 410128)
【目的】研究不同氮效率油菜品种碳素累积转运差异,为揭示氮高效品种协调籽粒碳氮代谢矛盾、促进油分形成的机理提供理论依据。【方法】采用土培试验,以不同氮效率油菜品种为供试材料,在正常供氮和氮胁迫条件下,研究不同生育期(抽薹期、开花期、角果发育期和收获期)碳素累积与器官分布的差异,并用13C标记技术测定营养器官碳素向生殖器官的再分配比例与再分配量,分析碳素累积转运对籽粒产量形成与油分累积的影响。【结果】氮高效品种的籽粒油分含量略高于氮低效品种,但2种供氮水平下品种间差异均未达到显著水平,而油分产量氮高效品种显著高于氮低效品种;与氮胁迫处理相比,正常供氮处理的油分含量略有降低,但油分产量显著增加。植株碳素累积量各生育期都表现为氮高效品种略高于氮低效品种,但品种间差异也均未达到显著水平;同一品种不同供氮水平处理之间的碳素累积量差异较大,正常供氮处理显著高于氮胁迫处理。不同氮效率油菜品种器官碳素分配比例存在差异,氮高效品种抽薹期和开花期叶片和根的碳素分配比例以及角果发育期和收获期角果与籽粒的碳素分配比例均大于氮低效品种,而全生育期茎中碳分配比例以及角果发育期和收获期根中的碳素分配比例却小于氮低效品种。与氮胁迫处理相比,正常供氮处理的抽薹期和开花期叶中碳素分配增多、根中碳素分配减少,收获期籽粒的碳素分配也是正常供氮处理高于氮胁迫处理。随着生殖生长进程,营养器官碳素向生殖器官的再分配比例和量逐渐增加,品种间差异也逐渐加大。开花期向花的再分配比例和量,氮胁迫条件下氮高效品种低于氮低效品种,正常供氮条件下则相反,但2种氮水平下的品种间差异均不显著;角果发育期向角果的再分配比例和量以及收获期向籽粒的再分配比例和量,2个氮水平均表现为氮高效品种高于氮低效品种,但只有正常供氮条件下差异显著;收获期向角果皮的再分配比例和量,氮胁迫条件下氮高效品种低于氮低效品种,正常供氮条件下则相反,但只有氮胁迫条件下差异显著。油菜收获时50%以上抽薹期累积碳素已离开营养器官,抽薹期累积的碳素减少比例与向生殖器官转运再分配的碳素比例具有相同的处理间变化趋势,但由于碳水化合物的呼吸消耗,碳素减少比例远大于碳素转运再分配比例。【结论】不同氮效率油菜品种各生育期碳素累积量并没有明显差异,但是氮高效品种生长后期有更多的营养器官碳素向生殖器官尤其是向籽粒转运,这是氮高效品种籽粒形成过程中争取更多碳源,缓解碳氮代谢矛盾,促进油分形成的重要机理之一。
油菜;碳素累积;碳素转运;油分形成;氮效率(1College of Resources and Environmental Sciences, Hunan Agricultural University/ Southern Regional Collaborative Innovation Center for Grain and Oil Crops in China/National Engineering Laboratory of Soil and Fertilizer Resources Efficient Utilization/ Hunan Provincial Key Laboratory of Farmland Pollution Control and Agricultural Resources Use/Hunan Provincial Key Laboratory of Plant Nutrition in Common University, Changsha 410128;2National Center of Oilseed Crops Improvement, Hunan Branch, Changsha 410128)
0 引言
【研究意义】油菜是需氮量多、籽粒油分与蛋白质含量之间矛盾比较突出的作物,加强氮素营养,可提高油菜产量,但也会因蛋白质累积增多而降低油分含量。因此,在提高油菜氮素利用效率的同时,合理调节碳氮代谢,对协调籽粒油分与蛋白质含量的矛盾具有重要的意义。【前人研究进展】籽粒中的碳素,一方面来自生殖器官本身的光合作用,另一方面来自营养器官碳素的再分配,促进作物生长后期碳素的再分配,可协调碳氮代谢对碳骨架的竞争,从而实现作物产量与品质双赢。目前,不同栽培措施对作物生长后期碳素转运再分配的影响以及产量对其的响应方面有较多研究报道[1-3],并提出了水稻碳素再分配模型[4],碳素再分配在保障逆境条件下作物稳产的作用也得到了证实[5-6]。但是,碳素再分配对籽粒品质的影响或对缓解产量与品质之间矛盾的贡献方面仅有少量研究报道。BENDEVIS等[7]研究表明,碳氮化合物的累积与再分配有利于提高藜麦产量和改善品质。CLAUDIA等[8]指出小麦生长后期促进营养器官碳水化合物向籽粒的再分配,可在增加产量的同时不降低籽粒蛋白质含量;高蛋白小麦品种花后碳水化合物的转运量高于低蛋白品种[9]。【本研究切入点】ZHANG等[10]研究表明与低氮效率油菜品种相比,高氮效率油菜品种虽然生长后期氮素再利用能力强、籽粒氮素累积量高,但籽粒油分含量并没有下降反而还高,说明氮高效油菜品种籽粒形成过程的碳骨架来源相对充足,而其中除了氮高效油菜品种角果发育期具有更强的光合能力之外,是否还存在更高的碳水化合物再分配机理,目前尚不清楚。【拟解决的关键问题】本文采用13C标记技术,研究不同氮效率油菜品种生长后期碳素转运再分配差异及其对油分累积的影响,以期揭示高氮效率油菜品种协调籽粒油分与蛋白质含量之间矛盾的生理机制,为实现油菜的优质高产提供重要参考。
1 材料与方法
1.1试验材料
供试油菜品种为课题组经过大田试验筛选出的相对氮高效品种27号和相对氮低效品种6号[11-12],分别用H和L表示。其中,氮效率指单位介质氮素所形成的产量,筛选时以参比品种的生育期相近为前提,按以下公式进行计算,氮效率=产量/介质氮素(包括肥料氮素和土壤氮素)[13]。2个油菜品种的籽粒产量及氮素吸收利用特性见表1所示。

表1 2个油菜品种的籽粒产量及氮素利用特性比较Table 1 Comparison of grain yield and nitrogen utilization characteristics of two Brassica napus
供试土壤为河流冲积物发育的河潮土,其化学性质为有机质22.14 g·kg-1、全氮1.42 g·kg-1、碱解氮105.31 mg·kg-1、全磷1.03 g·kg-1、速效磷49.71 mg·kg-1、全钾19.93 g·kg-1和速效钾137.02 mg·kg-1,pH5.13。供试肥料为氮肥用尿素(含氮46%),磷肥用过磷酸钙(含P2O512%),钾肥用氯化钾(含K2O 60%),硼肥施用硼砂(含硼11%)。用丰度为99%的13CO2气体(上海上化院天乐实业有限公司提供)进行13C标记。
1.2试验设计
用高25.7 cm,内径19.2 cm白色瓷钵,装土6.25 kg,进行土培试验,于2014年9月至2015年5月期间,在湖南农业大学农业资源与环境试验基地玻璃顶网室进行。采用育苗移栽方法,2014年9月24日播种、10月24日移栽,每钵一株,于2015年5月10日收获。
试验设品种和氮素供应两水平两因子,品种选氮低效品种和氮高效品种,氮素供应水平设正常供氮和氮胁迫2个水平,共4个处理,即氮低效品种氮胁迫处理(L)、氮高效品种氮胁迫处理(H)、氮低效品种正常供氮处理(L-N)和氮高效品种正常供氮处理(H-N),每个处理布置20钵,共80钵。正常供氮处理施肥量为0.2 g N·kg-1土、0.1 g P2O5·kg-1土、0.15 g K2O·kg-1土、2.0 mg B·kg-1土,氮胁迫处理除不施氮肥外,其他肥料用量与正常供氮处理一致,所有肥料均作底肥一次施入。
1.3植株13C标记
13C标记过程在可密闭的塑料棚进行,塑料棚长14 m、宽4 m、高1.5 m。油菜抽薹初期(2月7—10日)将盆钵移到塑料大棚内,棚内每天释放1.5 L丰度为99%的13CO2气体,气体释放时间为9:00—15:00,大棚封闭时间为9:00—17:00,连续标记4 d,标记结束后放回玻璃顶网室继续培养。
1.4取样与测定方法
分别于标记结束后2 d(2月12日)、开花期(3月12日)、角果发育期(4月15日)和收获期(5月10日)采5株全株样品,具体方法是:先把地上部分刈割下来,然后往盆钵里加满水浸泡约3 h,待土泡软之后连根带土整体倒出,用自来水冲洗泥土,随着泥土冲走和根系裸露量增多,水龙头开启度逐渐减小,以免冲走小细根,泥土冲先干净后带回实验室进一步清洗干净。从13C标记结束开始收集落叶,为防止植株间落叶的混杂,将盆钵放进在直径1 m、高0.18 m的塑料大盆中,每天收集1次。将样品洗净后按器官分装,烘干后测定干重,粉碎过筛后用Vario PYRO cube元素分析仪(Elemental公司)测定各器官碳素含量和13C丰度。
1.5数据计算与处理
用Microsoft Excel 2007整理试验数据,不同处理间差异采用DPS软件进行差异性检验。碳素累积量、碳素转运比例与转运量按以下公式进行计算:
某器官碳素累积量=该器官碳素含量×该器官干物质累积量;
某器官13C累积量=碳素累积量×该器官13C丰度;
营养器官碳素向生殖器官的转运比例=(生殖器官13C累积量/标记2 d后植株13C累积量)×100%;
营养器官碳素向生殖器官的转运量=标记2 d后植株碳素累积量×营养器官碳素向生殖器官的转运比例;
营养器官中抽薹期累积碳素的减少比例=(标记2 d后植株13C累积量-收获期根、茎、老叶13C累积量)/标记2 d后植株13C累积量。
2 结果
2.1不同氮效率油菜品种油分累积差异
不同氮效率油菜品种在不同供氮水平条件下籽粒油分含量和油分产量分析(图1)表明,在氮胁迫条件下,氮低效品种和氮高效品种的油分含量分别为43.2%和44.1%,在正常供氮条件下,氮低效品种和氮高效品种的油分含量分别为40.5%和41.4%;而上述4个处理的油分产量依次为4.3、6.1、7.1和9.5 g/株。即无论供氮水平如何,氮高效品种的籽粒油分含量略高于氮低效品种,但差异未达到显著水平。但是,由于氮高效品种的籽粒产量显著高于氮低效品种,其油分产量却显著高于氮低效品种。与氮胁迫处理相比,正常供氮处理的2个油菜品种油分含量均有所下降,但差异不显著,而油分产量则显著增加,这也是因为正常供氮处理的籽粒产量显著高于氮胁迫处理所致。

图1 油分含量和油分产量Fig. 1 Oil contents and yields
2.2不同氮效率油菜品种碳素累积与分配差异
碳素是油分形成的重要物质基础。各生育期植株碳素累积量的测定结果(图2)表明,虽然各生育期都表现为氮高效品种略高于氮低效品种,但品种间的差异较小。而同一品种不同供氮水平之间的差异较大,正常供氮处理显著高于氮胁迫处理。
不同氮效率油菜品种各器官碳素分配略有不同(图3)。与氮低效品种相比,氮高效品种抽薹期和开花期叶片和根中碳素分配较多,茎中碳素分配较少;角果发育期角果和叶中碳素分配较多,茎和根中碳素分配较少;收获期籽粒中碳素分配较多,也是茎和根中碳素分配较少,2种供氮水平下的变化趋势类似。说明氮高效品种抽薹期和开花期吸收器官(根系)和同化器官(叶片)中的碳素分配以及角果发育期和收获期生殖器官中的碳素分配有一定优势。
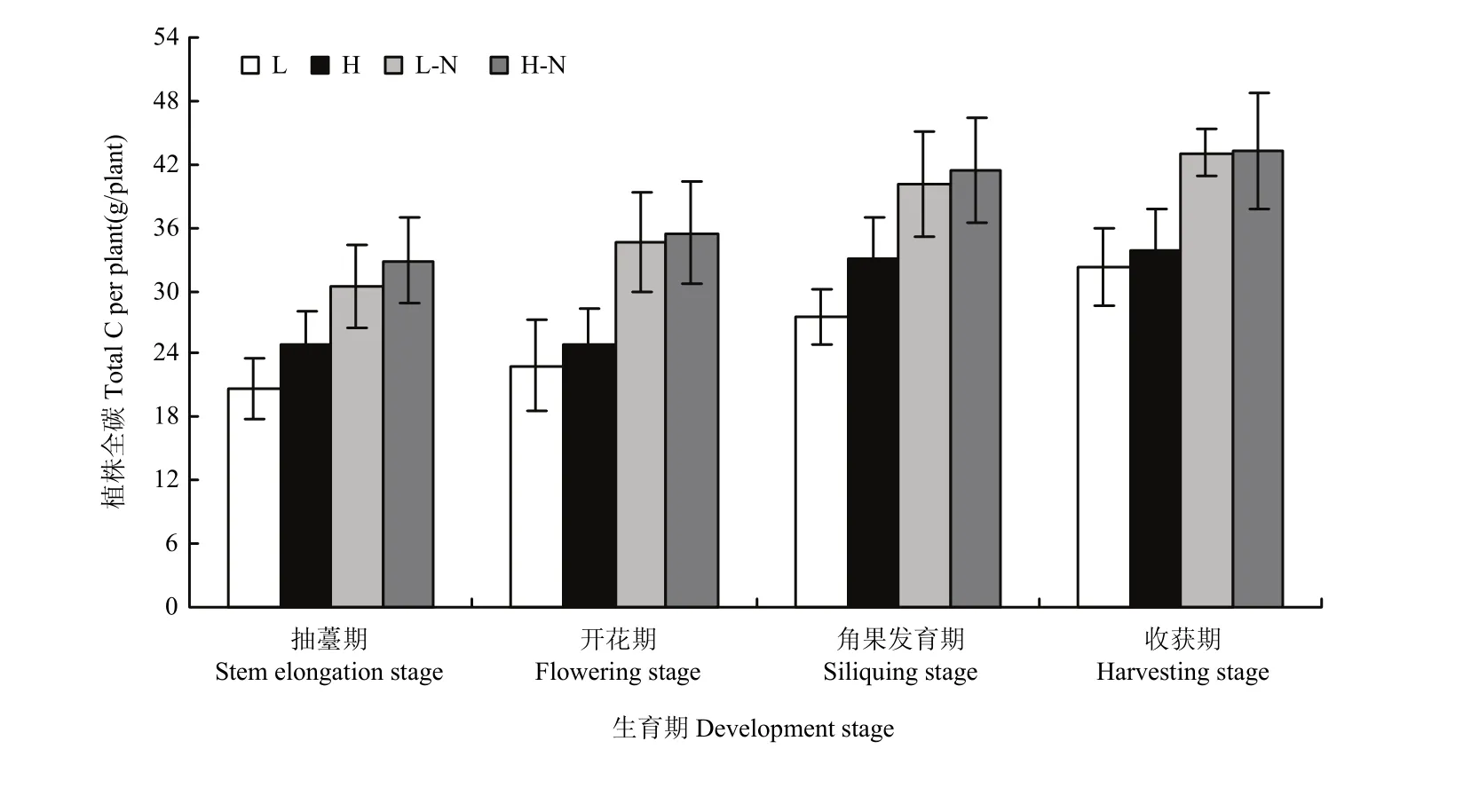
Fig. 2 Carbon accumulation amount in plant图2 植株碳素累积量

图3 各器官碳素累积量占植株总碳素累积量的比例Fig. 3 Carbon allocation proportion in plant organs
比较2种氮水平下碳素分配的差异表明,氮胁迫条件下抽薹期和开花期根中的分配有较大优势,但生育后期该优势减弱,这主要是因为氮胁迫处理促进根系生长,但又引起生长后期根系早衰,而生长前期的根系生长以减少地上部生长为代价,具体表现为抽薹期和开花期叶片以及收获期籽粒中的碳素分配减少。
2.3不同氮效率油菜品种生长后期碳素转运再分配差异
油菜生长后期营养器官碳素向生殖器官转运再分配比例(表2)的测定结果表明,开花期的再分配比例施氮条件下氮高效品种略高于氮低效品种,氮胁迫条件下正好相反,但品种间差异2种氮水平下均未达到显著水平。角果发育期的再分配比例以及收获期向籽粒的再分配比例,2种氮水平下均表现为氮高效品种高于氮低效品种;角果发育期向角果的转运再分配比例,氮高效品种比氮低效品种高出52.3%(正常供氮处理)和25.1%(氮胁迫处理);收获期向籽粒转运再分配的碳素比例,氮高效品种比氮低效品种高出107.9%(正常供氮处理)和31.3%(氮胁迫处理),但只有正常供氮条件下差异达到了显著水平。试验还表明,收获期向角果皮的转运再分配比例,正常供氮条件下品种间差异较小,而在氮胁迫条件下氮高效品种明显低于氮低效品种,说明,在氮胁迫导致碳素累积量下降的情况下,氮高效品种尽可能将有限的碳素分配到籽粒,这无疑是有利于油分的累积。

表2 营养器官碳素向生殖器官的转运比例Table 2 Redistribution proportion of C from vegetative organs to reproductive organs (%)
转运再分配量(表3)的变化趋势与转运比例基本一致,角果发育期向角果的碳素再分配量,氮高效品种比氮低效品种高出50.7%(正常供氮处理)和31.4%(氮胁迫处理);收获期向籽粒的碳素再分配量,氮高效品种比氮低效品种高出75.1%(正常供氮处理)和23.6%(氮胁迫处理)。试验还表明,收获期再分配到生殖器官的总碳中籽粒碳所占的比例,在正常供氮条件下氮高效品种和氮低效品种分别为74.6%和61.1%,在氮胁迫条件下对应值分别为85.0%和66.7%,即无论供氮水平如何氮高效品种向籽粒再分配的碳明显多于氮低效品种,而向角果皮再分配的碳则相反,可见,氮高效品种为籽粒油分形成,提供更好的碳物质基础。
2.4抽薹期营养器官中累积碳素的减少比例

表3 营养器官碳素向生殖器官的转运量Table 3 Redistribution amount of C from vegetative organs to reproductive organs (g)
随着生育期进程,生长前期累积在营养器官中的碳素逐渐减少,其主要去向有二:一是参与呼吸作用后以CO2的形式散失到空气中;二是向生殖器官转运再分配。通过对营养器官中抽薹期累积的碳素的测定,计算营养器官中抽薹期累积的碳素减少比例结果表明(图4),在正常供氮处理的氮高效品种中最高,为74.3%,在正常供氮处理的氮低效品种中最低,为54.2%,2种供氮水平下均表现为氮高效品种大于氮低效品种,但只有在正常供氮条件下的品种间差异达到了显著水平,与碳素转运再分配的变化趋势一致。

图4 抽薹期营养器官中累积碳素的减少比例Fig. 4 Reduction proportion of C accumulation during bolting stage in vegetativeorgans
3 讨论
植株体内的碳素累积直接影响作物产量与品质的形成。由于植物体内各种代谢、尤其是碳氮代谢之间存在碳骨架竞争,导致了粮油作物产量与品质之间的矛盾。即作物开花后仍维持较高的植株氮素水平虽然有利用于籽粒蛋白质含量的提高,但易造成贪青晚熟而减产[14]。而对油菜来说,较好的氮素营养虽可提高籽粒产量和蛋白质含量,但也减少籽粒油分含量。这说明加强作物生长后期的氮素营养以后,生殖器官本身光合速率提高所增多的碳素,往往不足以弥补其含氮物质增多所增加的碳素需求。因此,协调碳氮代谢之间的关系,实现作物的高产优质是农业科学家一直关注的问题。在此方面,小麦、玉米、水稻等禾谷类作物的研究领先于其他作物,研究工作主要集中在如何提高光合速率合成更多碳水化合物以及促进生长后期营养器官碳素(或干物质)向生殖器官的转运再分配[15-17]等方面,而作物不同氮效率品种之间的碳素累积再分配差异及其对品质的影响方面研究报道很少。
目前,有关氮效率的内涵并没有统一的定义,但是在不同作物品种氮效率差异的机理研究方面,多数采用MOLL等[13]的方法评价氮效率,即氮效率指单位介质氮素(肥料氮素+土壤氮素)所形成的产量[18-21]。本试验供试材料的筛选过程中,分别用以下2种方法表示土壤氮素供应量:一是播种前土壤速效氮和油菜生长季内土壤有机氮矿化量加和;二是不施氮区植株氮素吸收量和收获后土壤速效氮加和,发现2种表示方法的氮效率评价结果一致。该评价指标的优点是可将氮效率解析为氮素吸收效率和氮素利用效率(氮效率=氮素吸收效率×氮素利用效率),前者指植株吸收的氮素占氮素供应量之比[氮素吸收效率=(植株氮素累积量/介质氮素)×100%],后者则指植株吸收的单位氮素所形成的产量(氮素利用效率=产量/植株氮素累积量)。但在实际操作过程中,因难以准确计算土壤氮素供应量的具体值,在同一土壤条件和施氮量一致的情况下,一般用产量和植株氮素累积量表示氮效率和氮素吸收效率[22-23]。因此,该评价指标可确定氮效率差异中氮素吸收效率和氮素利用效率的各自贡献,将复杂的氮效率性状分解为更小的单位性状,以便进一步分析氮效率特异性指标。本试验的2个供试品种在等氮水平下产量差异显著,在氮胁迫条件下氮素吸收量差异显著,在正常供氮条件下氮素利用效率差异显著,说明不同供氮水平下氮素吸收效率和氮素利用效率对氮效率差异的贡献有所不同,氮胁迫条件下氮素吸收效率的贡献较大,正常供氮条件下氮素利用效率的贡献较大[14]。但是,氮收获指数的品种间差异并没有因供氮水平不同而发生变化,氮高效品种的氮收获指数2个供氮水平下均显著高于氮低效品种,即无论供氮水平如何氮高效品种都有更多的氮素分配到籽粒中。有趣的是,本试验中氮高效品种的籽粒油分含量并没有因籽粒氮素的增加而降低,说明氮高效油菜品种籽粒形成过程中有更充足的碳素来源。已经证明,角果发育期角果本身的光合状况对籽粒产量、含油量及脂肪酸组成有显著影响[24-25],那么氮高效品种除角果本身具有更强的光合能力之外,还有没有其他碳素来源优势呢?本试验数据显示,2个油菜品种生长前期的碳素总累积量差异不大,但生长后期营养器官碳素向生殖器官的转运再分配氮高效品种有明显优势,这为协调碳氮代谢对碳骨架的竞争、缓解籽粒油分与蛋白质含量的矛盾提供了物质基础。另有研究证明,油菜前期吸收的氮、磷、钾等养分对后期籽粒的充实至关重要[26],且氮高效品种在生长后期氮素转运再利用方面具有明显优势[27]。可以推断,生长后期的碳素转运再利用对油菜籽粒形成期间更好地发挥氮、磷、钾等矿质养分的作用,提高养分利用效率具有重要意义。本试验还表明,在氮胁迫导致碳素总累积量以及转运再分配量减少的情况下,氮高效品种显著降低向角果皮的碳素再分配,可见,氮高效品种的碳素再分配优势不仅在碳素再分配比例和量方面,还体现在可及时根据碳源情况调节碳素再分配方向,以满足籽粒形成过程的碳素需求。这是氮高效品种在高效吸收利用氮素的同时不降低籽粒油分含量,并协同产量优势效应显著提高油分产量的重要机理之一。以上研究为实现油菜籽粒油分和蛋白质含量的双赢提供了理论依据,也为加强氮素营养,提高氮收获指数和氮素利用效率,实现作物高产优质[7-8]的生产实践提供了佐证。
由于生长后期再分配到籽粒中的碳素来自营养器官,因此,收获期营养器官碳素的减少情况可为不同氮效率油菜品种之间的碳素再分配差异提供旁证。本试验中抽薹期累积的碳素减少比例与向生殖器官再分配的碳素比例有相同的变化规律,只是因为呼吸作用释放的CO2较多,向生殖器官再分配的比例远小于营养器官碳素的减少比例。
4 结论
4.1油菜生长后期营养器官碳素向生殖器官的转运再分配比例和量,氮高效品种均高于氮低效品种,正常供氮水平下差异显著。这是氮高效品种籽粒形成过程中争取更多碳源,促进油分形成的重要机理之一。
4.2氮高效品种的植株碳素累积量略高于氮低效品种,生长前期叶片和根的碳素分配比例以及生长后期角果与籽粒的碳素分配比例也较高。
4.3营养器官中抽薹期累积碳素的减少比例与生长后期向生殖器官转运再分配碳素的变化趋势一致。
References
[1] 许育彬, 沈玉芳, 李世清. CO2浓度升高和施氮对冬小麦花前贮存碳氮转运的影响. 作物学报, 2011, 37(8): 1465-1474.
XU Y B, SHEN Y F, LI S Q. Effect of elevated CO2concentration and nitrogen application on trans-location of dry matter and nitrogen restored before anthesis in Winter wheat. Acta Agronomica Sinica,2011, 37(8): 1465-1474. (in Chinese)
[2] 易镇邪, 陈平平, 屠乃美, 王璞. 包膜尿素与普通尿素对夏玉米碳氮代谢的影响. 湖南农业大学学报(自然科学版), 2011, 37(3): 225-232.
YI Z X, CHEN P P, TU N M, WANG P. Effect of coated and uncoated normal urea on carbon and nitrogen metabolism in summer maize. Journal of Hunan Agricultural University (Natural Sciences), 2011,37(3): 225-232. (in Chinese)
[3] 田青兰, 刘波, 钟晓媛, 赵敏, 孙红, 任万军. 不同播栽方式下杂交籼稻非结构性碳水化合物与枝梗和颖花形成及产量性状的关系.中国农业科学, 2016, 49(1): 35-53.
TIAN Q L, LIU B, ZHONG X Y, ZHAO M, SUN H, REN W J. Relationship of NSC with the formation of branches and spikelets and the yield traits of Indica hybrid rice in different planting methods. Scientia Agricultura Sinica, 2016, 49(1): 25-33. (in Chinese)
[4] TOMMASO S, SIMONE B, ROBERTO C. A model to simulate the dynamics of carbohydrate remobilization during rice grain filling. Ecological Modelling, 2016, 320: 366-371.
[5] 潘俊峰, 李国辉, 崔克辉. 水稻茎鞘非结构性碳水化合物再分配及其在稳产和抗逆中的作用. 中国水稻科学, 2014, 28(4): 335-342.
PAN J F, LI G H, CUI K H. Re-partitioning of non-structural carbohydrates in rice stems and their roles in yield stability and stress tolerance. Journal of Rice Science, 2014, 28(4): 335-342. (in Chinese)
[6] CLAUDIA T, GUILLERMO B, GABRIEL E, HERMINDA R. ABA action on the production and redistribution of field-grown maize carbohydrates in semiarid regions. Journal of Plant Growth Regulation, 2012, 67: 27-34.
[7] BENDEVIS M A, SUN Y, ROSENQVIST E, SHABALA S, LIU F,JACOBSEN S E. Photoperiodic effects on short-pulse 14C assimilation and overallcarbon and nitrogen allocation patterns in contrasting quinoacultivars. Environmental and Experimental Botany,2014, 104: 9-15.
[8] CLAUDIA T, ANA C C, HERMINDA R, CARLOS C, RUBE´N B. Exogenous abscisic acid increases carbohydrate accumulation and redistribution to the grains in wheat grown under field conditions of soil water restriction. Journal of Plant Growth Regulation, 2007, 26: 285-289.
[9] 杨铁钢, 戴廷波, 曹卫星. 施氮水平对不同品质类型小麦花后碳氮同化和转运的影响. 南京农业大学学报, 2008, 31(2): 6-10.
YANG T G, DAI T B, CAO W X. Effects of nitrogen rates on assimilation and translocation of carbon and nitrogen after anthesis in two wheat cultivars. Journal of Nanjing Agricultural University, 2008,31(2): 6-10. (in Chinese)
[10] ZHANG Z H, SONG H X, LIU Q, RONG X M, PENG J W, XIE G X,ZHANG Y P, CHEN L R, GUAN C Y, GU J D. Responses of seed yield and quality to nitrogen application levels in two oilseed rape(Brassica napus L.) varieties differing in nitrogen efficiency. Plant Production Science, 2012, 15(4): 265-269.
[11] HAN Y L, SONG H X, LIAO Q, YU Y, JIAN S F, LEPO J E, LIU Q,RONG X M, TIAN C, ZENG J, GUAN C Y, ISMAIL A M, ZHANG Z H. Nitrogen use efficiency is mediated by vacuolar nitrate sequestration capacity in roots of Brassica napus. Plant Physiology,2016, 170(3): 1684-1698.
[12] HAN Y L, LIU Q, GU J D, GONG J M, GUAN C Y, LEPO J E,RONG X R, SONG H X, ZHANG Z H. V-ATPase and V-PPase at the tonoplast affect NO3- content in Brassica napus by controlling distribution of NO3- between the cytoplasm and vacuole. Journal of Plant Growth Regulation, 2015, 34: 22-34.
[13] MOLL R H, KAMPRATH E J, JACKSON W A. Analysis and interpretation of factors which contribution efficiency of nitrogen utilization. Agronomy Journal, 1982, 74: 562-568.
[14] ACRECHEE M M, SLAFER G A. Variation of grain nitrogen content in relation with grain yield in old and modern Spanish wheats grown under a wide range of agronomic conditions in a Mediterranean region. Journal of Agricultural Science, 2009, 147(6): 657-667.
[15] 张法全, 王小燕, 于振文, 王西芝, 白洪立. 公顷产10000 kg小麦氮素和干物质积累与分配特性. 作物学报, 2009, 35(6): 1086-1096.
ZHANG F Q, WANG X Y, YU Z W, WANG X Z, BAI H L. Characteristics of accumulation and distribution of nitrogen and dry matter in wheat at yield level of ten thousand kilograms per hectare. Acta Agronomic Sinica, 2009, 35(6): 1086-1096. (in Chinese)
[16] 吕丽华, 陶洪斌, 王璞, 赵明, 赵久然, 鲁来清. 施氮量对夏玉米碳、氮代谢和氮利用效率的影响. 植物营养与肥料学报, 2008,14(4): 630-637.
LÜ L H, TAO H B, WANG P, ZHAO M, ZHAO J R, LU L Q. The effect of nitrogen application rate on carbon and nitrogen metabolism and nitrogen use efficiency of summer maize. Plant Nutrition and Fertilizer Science, 2008, 14(4): 630-637. (in Chinese)
[17] 李敏, 张洪程, 杨雄, 葛梦婕, 马群, 魏海燕, 戴其根, 霍中洋, 许轲. 水稻高产氮高效型品种的物质积累与转运特性. 作物学报,2013, 39(1): 101-109.
LI M, ZHANG H C, YANG X, GE M J, MA Q, WEI H Y, DAI Q G,HUO Z Y, XU K. Characteristics of dry matter accumulation and translocation in rice cultivars with high yield and high nitrogen use efficiency. Acta Agronomica Sinica, 2013, 39(1): 101-109. (in Chinese)
[18] ALLEN G G, ASHOK K S, DOUGLAS G M. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends in Plant Science, 2004, 9(12): 597-605.
[19] SVEČNJAK Z, RENGEL Z. Canola cultivars differ in Nitrogen utilization efficiency at vegetative stage. Field Crops Research, 2006,97(2): 221-226.
[20] 赵春波, 宋述尧, 赵靖, 张雪梅, 张越, 张松婷. 北方地区不同黄瓜品种氮素吸收与利用效率的差异. 中国农业科学, 2015, 48(8): 1569-1578.
ZHAO C B, SONG S Y, ZHAO J, ZHANG X M, ZHANG Y, ZHANG S T. Variation in Nitrogen uptake and utilization efficiency of different cucumber varieties in Northern China. Scientia Agricultura Sinica,2015, 48(8): 1569-1578. (in Chinese)
[21] ANDREAS S, WOLFGANG F, BENJAMIN W, ROD J S. Complementary diversity for nitrogen uptake and utilisation efficiency reveals broad potential for increased sustainability of oilseed rape production. Plant and Soil, 2016, 400 (1/2): 245-262.
[22] 刘强, 宋海星, 荣湘民, 彭建伟, 谢桂先, 张玉平. 不同品种油菜籽粒产量及氮效率差异研究. 植物营养与肥料学报, 2009, 15(4): 898-903.
LIU Q, SONG H X, RONG X M, PENG J W, XIE G X, ZHANG Y P. Studys on oilseed yield and nitrogen efficiency in different cultivars of oilseed rape (Brassica napus). Plant Nutrition and Fertilizer Science,2009, 15(4): 898-903. (in Chinese)
[23] 姜琳琳, 韩立思, 韩晓日, 战秀梅, 左仁辉, 吴正超, 袁程. 氮素对玉米幼苗生长、根系形态及氮素吸收利用效率的影响. 植物营养与肥料学报, 2011, 17(1): 247-253.
JIANG L L, HAN L S, HAN X R, ZHAN X M, ZUO R H, WU Z C,YUAN C. Effects of nitrogen on growth, root morphological traits,nitrogen uptake and utilization efficiency of maize seedlings. Plant Nutrition and Fertilizer Science, 2011, 17(1): 247-253. (in Chinese)
[24] LI J, ZHANG C L, MA N, YU L P, CHAO Y, LI L. Effects of ABA on photosynthetic characteristics of pods and yield of Brassica napus. Agricultural Science & Technology, 2012, 13(4): 760-762, 903.
[25] 王春丽, 海江波, 田建华, 杨建利, 赵晓光. 油菜终花后角果和叶片光合对籽粒产量和品质的影响. 西北植物学报, 2014, 34(8): 1620-1626.
WANG C L, HAI J B, TIAN J H, YANG J L, ZHAO X G. Infiluence of silique and photosynthesis on yield and quality of oilseed rape(Brassica napus L.) after flowering. Acta Botanica Borealioccidentalia Sinica, 2014, 34(8): 1620-1626. (in Chinese)
[26] 刘晓伟, 鲁剑巍, 李小坤, 卜容燕, 刘波. 冬油菜叶片的干物质及养分积累与转移特性研究. 植物营养与肥料学报, 2011, 17(4): 956-963.
LIU X W, LU J W, LI X K, PU R Y, LIU B. Studies on characteristics of dry matter and nutrient accumulation and transportation in leaves of winter oilseed rape. Plant Nutrition and Fertilizer Science, 2011,17(4): 956-963. (in Chinese)
[27] ZHANG Z H, SONG H X, LIU Q, RONG X M, PENG J W, XIE G X,ZHANG Y P, GUAN C Y. Distribution characters of absorption nitrogen in oilseed rape(Brassica napus L.) at different growth stages. Journal of Plant Nutrition, 2014, 37: 1648-1660.
(责任编辑杨鑫浩,李莉)
Differences in Carbon Accumulation and Transport in Brassica napus with Different Nitrogen Use Efficiency and Its Effects on Oil Formation
ZHANG Xiao-long1, ZHANG Zhen-hua1, SONG Hai-xing1, YU Jia-ling1, GUAN Chun-yun2
【Objective】The differences in carbon (C) accumulation and transport in Brassica napus with different nitrogen use efficiencies (NUE), were studied in order to supply a theoretical basis for elucidating the mechanism of high NUE genotypes in coordinating the contradiction between C and N metabolism and promoting oil formation.【Method】A soil culture experiment was conducted to study the differences of C accumulation and distribution in different organs of B. napus with different NUE at different growth stages (stem elongation stage, flowering stage, silique stage and harvest stage) under normal- and limited-N conditions, and using13C isotope labeling to determine the C redistribution proportion and amount from vegetative organs to reproductive organs,analyze the contribution of C accumulation and transport to grain yield and oil accumulation.【Result】No significant differences in grain oil content between high- and low-NUE genotypes under the normal and limited-N application levels were observed, but the oil yield of high-NUE genotype was significantly higher than that of low-NUE genotype. Compared with limited-N, oil content was slightly decreased, but oil production was increased significantly under normal-N. There was no differences in C accumulation amount between high- and low-NUE genotypes, while the differences in C accumulation amount at different N application levels was significant, C accumulation amount of normal -N was significantly higher than that of limited-N. C allocation proportion in plant organs between high- and low-NUE genotypes was different, C allocation proportion in roots and leaves of high-NUE genotype was higher than that of low-NUE genotype at stem elongation stage and flowering stage, and C allocation proportion in silique and grain of high-NUE genotype was higher than that of low-NUE genotype at silique stage and harvest stage. While, C allocation proportion in stem of high-NUE genotype was lower than that of low-NUE genotype at the whole growth stage, and C allocation proportion in stem and root of high-NUE genotype was lower than that of low-NUE genotype at the whole growth stage. Compared with limited-N,C allocation proportion in leaves was increased at stem elongation stage and flowering stage under normal-N, while C allocation proportion in root was decreased. In addition, C allocation proportion in grain at harvest stage under normal-N was higher than what was observed in limited-N. As the development of plant growth stages, C distribution proportion and amount from vegetative organs to reproductive organs was gradually increased, the differences between genotypes were increased. Redistribution proportion and amount of C from vegetative organs to silique at silique stage and redistribution proportion and amount of C from vegetative organs to grain at harvest stage in high-NUE genotype was higher than that in low-NUE genotype, the significant different was occurred under normal-N. Redistribution proportion and amount of C in silique of high-NUE genotype was lower than that in low-NUE genotype under limited-N, and reversed results was observed under normal-N, the significant difference was occurred under limited-N. The 50% or more of C accumulation at stem elongation stage was left from vegetative organs, which was corresponding with redistribution proportion into reproductive organs, but C reduction proportion was significantly higher than C re-distribution proportion, which was because of the carbohydrate respiration consumption. 【Conclusion】 There was no significant difference in C accumulation between high- and low-NUE B. napus genotypes at different growth stages. However, higher proportion of C was redistributed from vegetative organs to reproductive organs in high-NUE genotype than that in low-NUE genotype at later growth stages. This is one of the important mechanisms for high-NUE genotype possesses with higher C source to relieve the contradictions between C and N, and promote oil formation during the process of grain formation.
Brassica napus; carbon accumulation; carbon transport; oil formation; nitrogen use efficiency
2016-01-25;接受日期:2016-05-23
国家自然科学基金(31372130,31101596)、国家“十二五”科技支撑计划(2012BAD15BO4,2014BAC09B01-01)
联系方式:张晓龙,E-mail:zhangxl0702@163.com。通信作者宋海星,E-mail:shx723@163.com。通信作者张振华,E-mail:zhzh1468@163.com
猜你喜欢
广西糖业(2024年1期)2024-04-25 17:59:28
机械工程师(2021年6期)2021-06-18 06:16:04
装备制造技术(2020年9期)2021-01-26 00:15:08
华北农学报(2020年4期)2020-08-29 04:11:00
工程与建设(2019年5期)2020-01-19 06:22:58
陕西农业科学(2019年11期)2019-12-25 03:37:22
陕西农业科学(2019年11期)2019-12-25 03:37:22
山东冶金(2019年3期)2019-07-10 00:53:58
今日自动化(2018年4期)2018-05-06 00:58:22
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:01
